
干旱胁迫下油橄榄品种光合特性研究
2017-01-03 01:39徐应杰丁春邦苏光灿
西北植物学报 2016年11期
兰 艳,徐应杰,谭 枫,丁春邦,李 天,*,苏光灿
(1 四川农业大学 农学院,四川温江611130; 2 四川农业大学 生命科学学院, 四川雅安 625014;3 凉山州中泽新技术开发有限责任公司,四川西昌615000)
干旱胁迫下油橄榄品种光合特性研究
兰 艳1,徐应杰1,谭 枫1,丁春邦2,李 天1,2*,苏光灿3
(1 四川农业大学 农学院,四川温江611130; 2 四川农业大学 生命科学学院, 四川雅安 625014;3 凉山州中泽新技术开发有限责任公司,四川西昌615000)
为揭示油橄榄 (OleaeuropaeaL.) 耐旱性与光合特性之间的关系,以筛选出的适宜于半干旱川西南地区种植的7个引进油橄榄品种为供试材料,采用盆栽模拟干旱胁迫的方法,研究持续干旱胁迫对其光合特性的影响。结果表明:(1) 随着干旱胁迫程度加剧,7个油橄榄品种叶片相对含水量均显著降低,至干旱胁迫后期 (25 d),各品种叶片均出现大幅失水,其中品种‘科拉蒂’失水率最高 (45.79%),而品种‘小苹果’失水率最低(25.52%),说明‘小苹果’叶片在干旱胁迫下较其他油橄榄品种具有更高保水能力。(2) 随着干旱胁迫程度加剧,7个油橄榄品种叶片光合色素含量均不同程度降低,表明光合色素分解量大于合成量;干旱胁迫持续25 d时,品种‘豆果’的叶绿素a和叶绿素b 含量下降幅度最大 (P<0.05),品种‘皮削利’类胡萝卜素含量下降幅度最大 (P<0.05),而品种‘小苹果’叶绿素a含量下降幅度最小。(3) 随着干旱胁迫的持续进行,各油橄榄品种叶片净光合速率 (Pn)、蒸腾速率 (Tr)、气孔导度 (Gs) 和胞间CO2浓度 (Ci) 均不同程度降低,而水分利用效率 (WUE) 则呈上升趋势;干旱胁迫期间,品种‘佛奥’的Pn、Tr和Ci以及‘皮削利’的Gs降幅均高于其他品种,而‘小苹果’的Pn、Gs和Ci降幅均为最小且WUE上升幅度最大。研究发现,在持续干旱胁迫条件下,油橄榄幼苗叶片均大幅失水,光合色素结构被破坏、色素分解、含量降低,同时气孔关闭蒸发减少,光合作用减弱,而供试油橄榄品种中‘小苹果’对干旱胁迫的适应性最强,适宜于在半干旱的川西地区种植。
干旱胁迫;油橄榄;光合特性;光合色素
近年来,随着全球气候与环境的不断恶化,加之季节性降水和降水分布区域的不均衡,导致水资源匮乏日趋明显,大部分国家和地区都受到不同程度的干旱威胁[1-3]。干旱导致土壤有效含水量减少,严重影响植物的生长、发育、繁殖并造成减产。因此,干旱胁迫已经成为限制植物正常生长的主要环境因素之一[4-6]。
光合作用是保障植物生长的最基本生理过程,是植物合成有机物和获取能量的根本来源。而干旱胁迫常常被认为是制约作物光合作用和生产力的最主要生态环境因子[5]。干旱胁迫下,叶绿素合成途径受阻且已合成的叶绿素分解加快导致叶绿素含量降低。李倩等[7]发现华山新麦草 (PsathyrostachyshuashanicaKeng) 叶绿素含量随干旱胁迫加强而不断下降。此外,干旱胁迫导致叶片气孔关闭,阻碍CO2吸收[8],降低光合效率,且降低的幅度取决于植物抗旱能力的强弱以及干旱胁迫的程度[4]。植物对干旱胁迫的响应是一个复杂的过程,可通过叶绿素含量的变化、气体交换规律以及内源调节物质改变响应抗旱性。因此,研究耐旱性与光合特性之间的关系尤为重要[9-11]。
油橄榄 (OleaeuropaeaL.) 属木犀科木犀榄属常绿乔木,又名洋橄榄、木犀榄、阿列布,是世界著名的四大木本油料经济树种之一。由其成熟鲜果经压榨获得的橄榄油被誉为“植物油中的皇后”和“飘香的软黄金”[12]。橄榄油营养丰富,不饱和脂肪酸和微量元素含量高[13, 14],具有阻止人体自由基的形成、增强人体免疫功能、预防心血管疾病等功效,可用于医疗用品、保健品和化妆品的生产,具有良好的营养和保健价值[12]。同时油橄榄树形美观,根系发达,具有保持水土、涵养水源和调节气候的作用[15]。近年来,随着栽培区域的扩展,在干旱和半干旱地区油橄榄栽种面积不断增加,然而由于灌溉条件限制,油橄榄生长受到严重抑制,从而制约该树种的经济和生态效益。因此,迫切需要筛选出耐旱的油橄榄品种,以满足干旱半干旱地区的栽培需求。
基于此,本研究以四川省凉山州引进的皮削利 (Picholine)、皮瓜尔 (Picual)、小苹果 (Manzanillo)、克罗莱卡 (Koroneiki)、佛奥 (Frantoio)、豆果 (Arbequina) 和科拉蒂 (Coratina) 等7个油橄榄品种为供试材料,采用盆栽模拟干旱胁迫的方法,探讨其在干旱胁迫下叶片的光合特性动态变化,旨在揭示干旱胁迫对引进油橄榄品种生长和光合作用的影响机制,为筛选出适宜于半干旱川西南地区种植的油橄榄品种及油橄榄的合理灌溉提供理论依据。
1 材料和方法
1.1 供试材料与试验地概况
选择凉山州中泽新技术开发有限公司引进的7个油橄榄品种的2年生无性系幼苗,在西昌北河油橄榄基地 (102.29 °E,27.92 °N) 进行试验。试验区域属亚热带高原季风气候,海拔1 540 m,气温年较差大,日较差小,年平均温度为17.2 ℃,年有效积温 (≥10 ℃) 3 500 ℃以上 (其中极端低温在1月,为-7.8 ℃,均温9.5 ℃;极端高温出现在7月,为36.5 ℃,均温22.6 ℃)。 太阳辐射强,日照充足。干湿季分明,降雨集中,降水时空分布差异明显,季节性用水不足,大部分农业生产需额外补水。
1.2 试验设计
2014年11月下旬,挑选长势相近的健康幼苗进行带土移栽至规格为26 cm×21 cm的黑色塑料钵内 (高21 cm),进行正常田间管理以保证其正常生长,待翌年7月份油橄榄生长需水期进行控水处理。试验在通风大棚内进行,除无降雨影响外,其他环境因子和自然条件相似。栽培土壤为沙壤土 (基质配比为沙土∶壤土∶有机肥=2∶2∶1),经测定,土壤肥力中上,具体营养含量分别为全N 0.100 g·kg-1、全P 0.046 g·kg-1、全K 2.383 g·kg-1、碱解N 1.066 mg·kg-1、速效P 0.654 mg·kg-1、速效K 73.449 mg·kg-1,田间最大持水量为25.20% (重量百分比),土壤容重为1.43 g·cm-3。
2015年7月中旬,从预处理材料中选取长势相近的各品种健康幼苗20株,带钵移至大棚内, 对所有供试幼苗进行透水处理后,用薄膜包住塑料钵以防止试验过程中土壤表面水分蒸发,同时阻止外界水分进入盆栽土壤,以自然干旱方式降低土壤水分。于透水处理后当日采样,作为试验对照 (正常供水条件,CK),之后每5 d为一个处理期,即分别干旱胁迫5、10、15、20、25 d时采样作为处理组,每次采样取3株重复。
1.3 测定项目与方法
1.3.1 叶片相对含水量 叶片相对含水量的测定采用饱和称重法。带回的样品迅速称其鲜重,3次重复,然后用蒸馏水浸泡6~8 h,至重量不再增加,得饱和重,用滤纸吸干表面水分,迅速称取饱和鲜重,再于105 ℃下杀青30 min,80 ℃下烘至恒重,即干重。
相对含水量=(鲜叶重-干重)÷(饱和鲜叶重-干重)×100%
1.3.2 光合色素含量 叶绿素含量测定采用张金政[3]等方法。将混合均匀叶样洗净去中脉剪碎,每品种称取3份,每份0.100 0 g。提取液配比为95%乙醇∶丙酮=1∶1,然后在UV-1800分光光度计下测定波长为663、646和470 nm时的吸光值A663、A646和A470,并按以下公式计算叶绿素a (Chla)、叶绿素b (Chlb) 和类胡萝卜素 (Car) 含量:
叶绿素a浓度Ca=12.21A663-2.81A646
叶绿素b浓度Cb=20.13A646-5.03A663
类胡萝卜素浓度CCar= (1 000A470-3.27Ca-104Cb)÷229
叶绿体色素含量(mg·g-1)=光合色素浓度×提取液体积×稀释倍数÷样品干重
1.3.3 光合气体交换参数 开始干旱处理后,每次取样前上午9:00~11:00,每个处理随机选择3盆,用GFS-3000便携式光合仪 (Walz , Germany),测定不同土壤水分条件下油橄榄幼苗顶端第4~6叶的光合强度。设置外界光强为1 000 μmol·m-2·s-1,测定指标为净光合速率(Pn)、蒸腾速率 (Tr)、气孔导度 (Gs) 和胞间CO2浓度 (Ci) 等参数,并计算水分利用效率(WUE)。
WUE=Pn/Tr
1.4 数据分析
采用SPSS 19.0对3次重复试验数据进行统计分析,利用最小显著差数法 (LSD)在α=0.05水平上做多重比较。
2 结果与分析
2.1 干旱胁迫下油橄榄品种叶片相对含水量的变化
7个油橄榄品种的叶片相对含水量均随干旱胁迫时间延长和胁迫程度的加剧呈下降趋势,但下降幅度有所不同,且不同品种间差异显著 (表1)。其中,在干旱胁迫处理前 (CK),‘克罗莱卡’的叶片相对含水量显著低于其他品种 (P<0.05)。干旱胁迫后,各品种叶片相对含水量变化趋势一致,但变化率表现出明显差异,具体为‘小苹果’<‘豆果’<‘佛奥’<‘克罗莱卡’<‘皮削利’<‘皮瓜尔’<‘科拉蒂’。至干旱胁迫后期,各品种叶片均出现大幅度失水,形态上表现出萎蔫;胁迫至25 d时,‘科拉蒂’失水率最高 (45.79%),‘小苹果’最低 (25.52%),两者的叶片相对含水量均与其它品种差异显著 (P<0.05)。可见,在土壤自然干旱胁迫条件下,‘小苹果’叶片比其它油橄榄品种具有更强保水能力,而品种‘科拉蒂’保水能力则明显较弱。
2.2 干旱胁迫下油橄榄品种叶片光合色素变化特征
光合色素参与光合作用过程,其含量直接影响植物的光合能力。光合色素中叶绿素(Chl) 和类胡萝卜素(Car) 与植物光合作用关系密切,尤其以叶绿素a (Chla) 最为重要。如图1,A 所示,随着干旱胁迫的加剧,各品种油橄榄叶片叶绿素a含量均呈逐渐下降趋势;在胁迫结束时(胁迫25 d),7个品种叶绿素a含量与CK相比均有不同程度的下降(‘豆果’>‘克罗莱卡’>‘佛奥’>‘皮削利’>‘科拉蒂’>‘皮瓜尔’>‘小苹果’),并以品种‘豆果’下降率最大(28.62%),‘小苹果’下降率最小(16.74%),此时的‘小苹果’、‘科拉蒂’叶绿素a含量显著低于其他品种(P<0.05),而‘皮削利’、‘皮瓜尔’和‘克罗莱卡’间叶绿素a含量差异均不显著 (P>0.05)。这说明不同品种油橄榄叶绿素含量a对干旱胁迫敏感性不一致,并以‘豆果’最敏感,‘小苹果’最稳定。
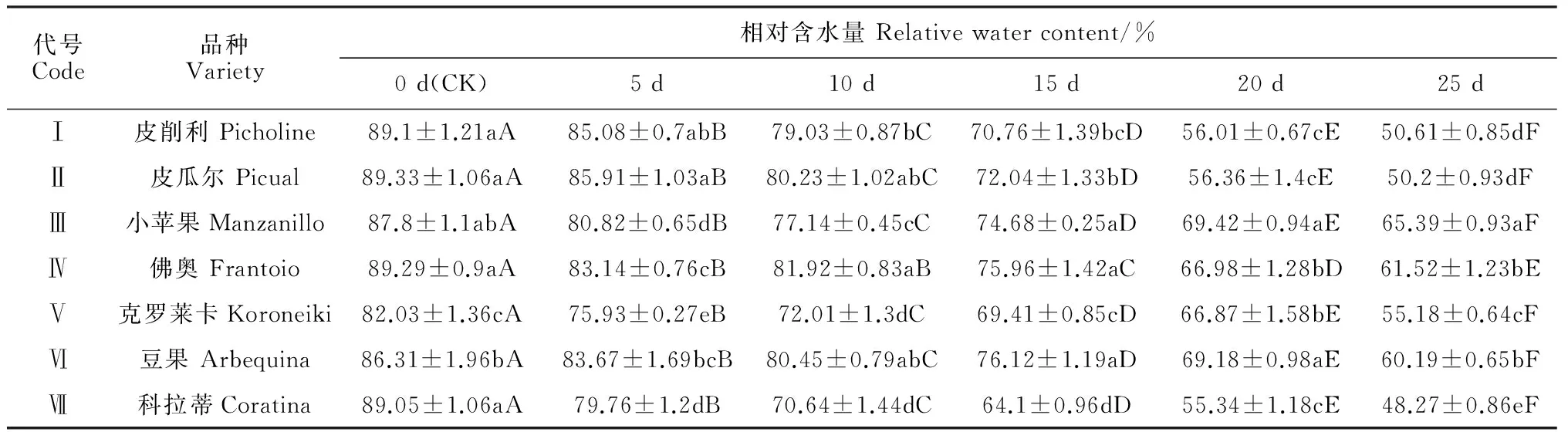
表1 干旱胁迫下油橄榄叶片相对含水量变化
注:表中同一列不同小写字母表示各处理在 0.05 水平上存在显著差异, 而同行不同大写字母表示各处理在0.05水平上存在显著差异
Note:The different lowercase letters indicate the significant differences among different varieties within the same time at 0.05 level. While the different capital letters indicate the significant differences among different treatments within the same variety at 0.05 level
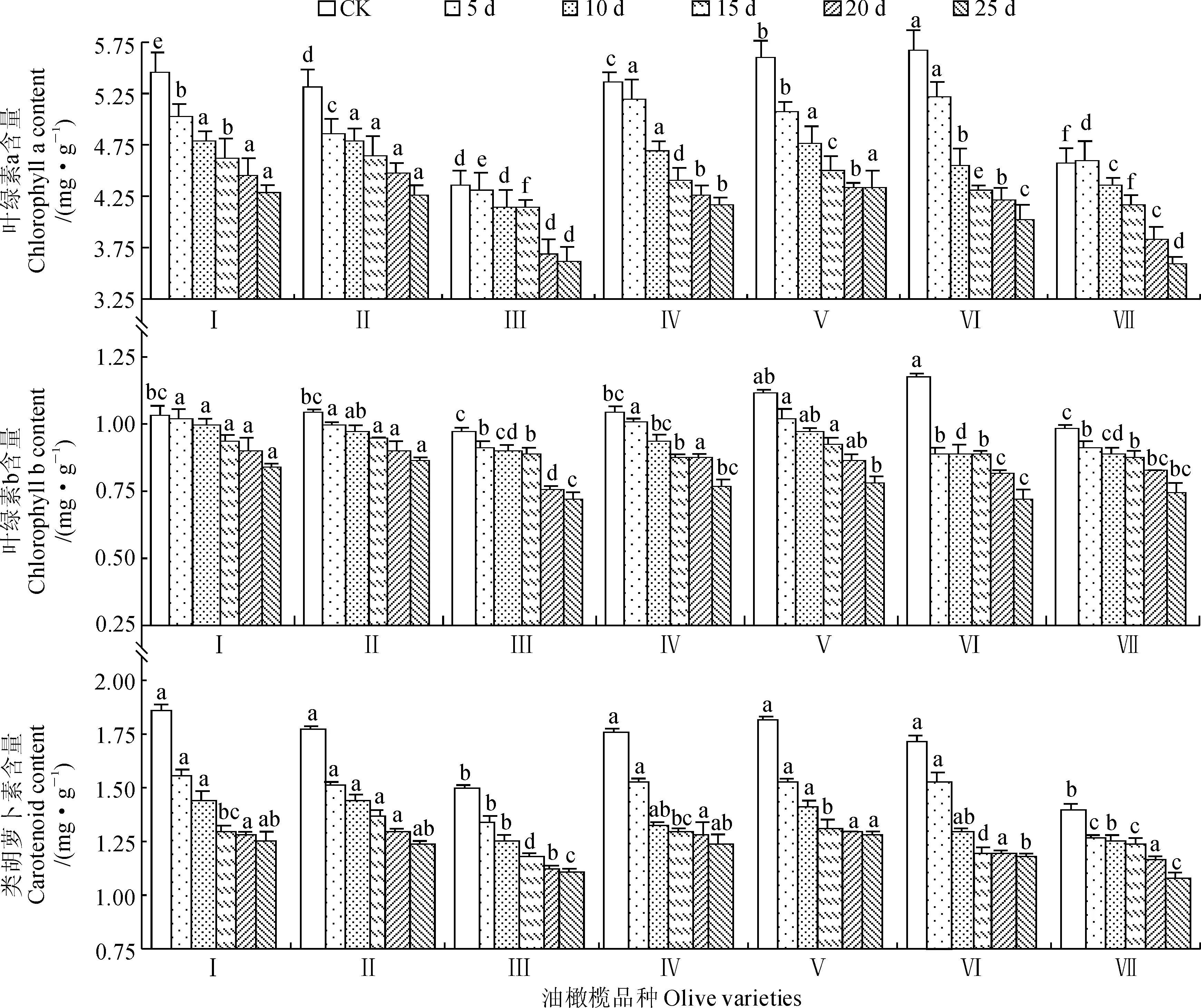
图1 干旱胁迫对不同油橄榄品种光合色素含量的影响Fig.1 Effects of drought stress on the photosgnthetic pigment content of different olive varieties
同时,随着干旱胁迫的加剧,各品种叶绿素b (Chlb) 含量也出现不同程度的下降 (图1,B)。胁迫结束时,7个品种叶绿素b含量与CK相比均有不同程度的下降 (‘豆果’>‘克罗莱卡’>‘佛奥’>‘小苹果’>‘科拉蒂’>‘皮削利’>‘皮瓜尔’),并且‘豆果’下降率最大 (38.64%),‘皮瓜尔’最小 (17.31%)。干旱胁迫前,‘豆果’与‘克罗莱卡’叶绿素b含量差异不显著 (P>0.05),但它们叶绿素b含量在胁迫至25 d时差异显著 (P<0.05),表明在胁迫过程中两者叶绿素b含量变化差异较大,‘豆果’叶绿素b含量对干旱胁迫更敏感。
另外,各品种叶片类胡萝卜素含量随着干旱胁迫的加剧和胁迫时间的延长也均呈逐渐下降趋势 ,降幅表现为‘皮削利’>‘豆果’>‘皮瓜尔’>‘克罗莱卡’>‘佛奥’>‘小苹果’>‘科拉蒂’,其中‘皮削利’下降率最大(30.54%),‘科拉蒂’最小 (21.09%)。在相同水分胁迫条件下,7个油橄榄品种类胡萝卜素表现出一定的差异性。在胁迫处理前,‘小苹果’和‘科拉蒂’的类胡萝卜素含量较低,且与其他品种差异显著 (P<0.05),而‘豆果’、‘佛奥’、‘皮瓜尔’和‘克罗莱卡’4个品种含量较高且其间差异不显著 (P>0.05);在胁迫结束时,‘小苹果’和‘科拉蒂’的类胡萝卜素含量仍然显著低于其他品种 (P<0.05)。表明持续干旱胁迫对‘小苹果’和‘科拉蒂’叶片类胡萝卜素含量变化影响较其他品种小,因此,较其他品种而言,‘小苹果’和‘科拉蒂’抗旱能力更强。
2.3 干旱胁迫对各油橄榄品种光合气体交换参数特性的影响
2.3.1 净光合速率 在持续干旱条件下,各油橄榄品种叶片净光合速率(Pn) 均随着胁迫程度加剧而逐渐降低(图2,A)。其中,在正常供水(CK) 条件下,各油橄榄品种的Pn以‘小苹果’最高(24.00 μmol·m-2·s-1),而‘皮瓜尔’最低 (19.47 μmol·m-2·s-1);在干旱胁迫结束后,‘小苹果’的Pn依然保持最高(5.64 μmol·m-2·s-1),而‘科拉蒂’Pn最低(2.81 μmol·m-2·s-1),此时叶片Pn除在‘皮削利’与‘皮瓜尔’间差异不显著 (P>0.05) 外,在其余品种间均差异显著 (P<0.05)。同时,各油橄榄品种Pn在不同干旱胁迫阶段的下降程度不同,如干旱胁迫至25 d时,7个品种Pn下降幅度 (85.96%~76.50%) 从大到小依次排序为‘佛奥’、‘科拉蒂’、‘皮削利’、‘豆果’、‘克罗莱卡’、‘皮瓜尔’、‘小苹果’。可见,‘小苹果’在各品种中表现出较强的耐旱性。

图2 干旱胁迫对不同油橄榄品种各光合参数的影响Fig.2 Effects of drought stress on the photosynthetic parameters of different olive varieties
2.3.2 蒸腾速率 图2,B 表明,在正常供水条件下,‘豆果’的蒸腾速率 (Tr) 最高 (6.88 mmol·m-2·s-1),而‘皮削利’最低 (5.39 mmol·m-2·s-1)。随着干旱胁迫时间延长和胁迫程度的加剧,各油橄榄品种叶片Tr均呈逐渐下降趋势,表明在干旱胁迫下各品种油橄榄幼苗均会通过降低蒸腾速率保持自身水分,以更好抵抗干旱。其中,在胁迫第一阶段 (5 d),‘小苹果’Tr下降率最低,而‘皮瓜尔’Tr下降率最高且达到显著水平 (P<0.05),表明‘皮瓜尔’蒸腾速率对干旱胁迫较敏感;当干旱胁迫至10 d时,‘豆果’和‘小苹果’的Tr下降率较高,表明‘豆果’Tr变化对干旱胁迫反应迟缓;在胁迫结束时,7个品种Tr下降幅度表现为‘佛奥’>‘科拉蒂’>‘豆果’>‘皮削利’>‘皮瓜尔’>‘小苹果’>‘克罗莱卡’,且品种间差异显著 (P<0.05),此时‘小苹果’的Tr最高,而‘佛奥’Tr最低,两者相比对照分别下降了87.27%、90.32%。
2.3.3 气孔导度 随着干旱胁迫时间延长,各油橄榄品种叶片气孔导度 (Gs) 也呈现出逐渐下降趋势,但品种不同下降幅度呈现差异 (图2,C)。其中,在正常供水 (CK) 条件下,‘小苹果’的Gs最高 (403.71 μmol·m-2·s-1),而‘佛奥’的Gs最低 (285.2 μmol·m-2·s-1),各品种Gs差异均达显著水平 (P<0.05);在干旱胁迫第一阶段 (5 d),‘小苹果’和‘佛奥’的Gs下降幅度较小,表明在轻度干旱胁迫下二者气孔导度敏感度较其他品种低;在干旱胁迫第二阶段 (5~10 d),‘小苹果’的Gs大幅度下降,表现出较其他品种更强的敏感性 ;在干旱胁迫第三阶段 (10~15 d),‘克罗莱卡’的Gs降幅最大;干旱胁迫结束后,7个品种Gs下降率表现为:‘皮削利’ (93.03%)>‘佛奥’ (92.02%) >‘科拉蒂’ (90.87%) > ‘豆果’ (90.59%)>‘克罗莱卡’ (87.12%)>‘皮瓜尔’ (84.79%)>‘小苹果’ (81.14%)。
2.3.4 胞间CO2浓度 图2,D显示,各油橄榄品种叶片胞间CO2浓度 (Ci)下降幅度在胁迫第一阶段 (5 d) 较大,在胁迫5~15 d 期间趋于平缓,但在胁迫后期 (20~25 d) 加速下降 ,总体呈现出“快-慢-快”的下降趋势。在正常供水(CK)条件下,油橄榄品种‘佛奥’的Ci最高 (384.01 μmol·mol-1),品种‘小苹果’则最低 (309.27 μmol·mol-1);在干旱胁迫结束时,各品种油橄榄叶片Ci下降率表现为‘佛奥’>‘皮削利’>‘克罗莱卡’>‘豆果’>‘皮瓜尔’>‘科拉蒂’>‘小苹果’,且各品种间差异显著,表明各品种Ci的变化对干旱胁迫的响应存在明显的差异。
综上表明,持续干旱胁迫对各品种油橄榄光合气体参数变化均有影响,且在各品种间有差异;其中品种‘小苹果’表现出较其他品种更显著的耐旱性。
2.4 干旱胁迫下油橄榄品种叶片水分利用效率变化
图3表明,在正常供水条件下 (CK),油橄榄品种‘皮削利’叶片水分利用效率最高 (4.42 μmol·mmol-1),而品种‘豆果’最低 (3.07 μmol·mmol-1)。随着干旱胁迫程度的加剧,品种‘皮削利’、‘佛奥’、‘豆果’和‘科拉蒂’叶片水分利用效率持续升高;品种‘小苹果’和‘克罗莱卡’在胁迫第一阶段略有降低,后期连续升高;品种‘皮瓜尔’则在胁迫第二阶段有下降趋势,但随后直至胁迫结束又持续上升。在干旱胁迫结束时,各品种油橄榄叶片水分利用效率均有不同程度的增加,且差异显著 (‘皮瓜尔’和‘佛奥’除外),其中的‘小苹果’叶片水分利用效率 (6.71 μmol·mmol-1) 显著高于其他品种 (P<0.05),‘科拉蒂’的水分利用效率 (4.36 μmol·mmol-1) 却显著低于其他品种 (P< 0.05)。以上结果表明各油橄榄品种的水分利用效率对持续干旱胁迫的响应规律并不一致。

图3 干旱胁迫对不同油橄榄品种水分利用效率的影响Fig.3 Effects of drought stress on the water use efficiency of different olive varieties
3 讨 论
3.1 油橄榄叶片相对含水量对干旱胁迫的响应特征
充足、合理均衡的水分是油橄榄幼苗生长发育过程中的必要环境因子之一[16]。土壤自然失水干旱和模拟梯度干旱是目前研究干旱胁迫的两种主要形式,为贴近油橄榄大规模种植的自然条件,本试验采用土壤自然失水干旱研究方法。叶片相对含水量作为水分生理指标,能够间接反映干旱胁迫下叶水势的高低,以及随干旱胁迫程度加深植物叶片水分亏缺的变化情况。因此,干旱胁迫下植物叶片相对含水量能反映植物的抗旱能力。前人在对高丛蓝莓、薰衣草属植物、结缕草等[17-19]植物的研究中发现,随着土壤含水量下降,叶片相对含水量明显下降,束缚水与自由水比值增大。本研究结果显示,大部分油橄榄品种随干旱胁迫时间延长而下降,这与Boussadia[2]、Guerfel等[20]的研究结果一致。在本试验干旱胁迫结束时,7个油橄榄品种叶片相对含水量表现为‘小苹果’>‘豆果’>‘佛奥’>‘克罗莱卡’>‘皮削利’>‘皮瓜尔’>‘科拉蒂’,说明品种‘小苹果’的抗旱能力较强,而品种‘科拉蒂’相对较弱。
3.2 油橄榄叶片光合色素含量对干旱胁迫对的响应特征
光合色素是植物叶片光合作用的物质基础,其含量的高低能直接反映植物叶片光合同化能力的强弱和植物生长发育的优劣。其中,叶绿素和类胡萝卜素与光合作用的关系较密切。叶绿素相对含量的变化能指示植物对干旱胁迫的敏感性,同时反映植物在逆境胁迫下同化物质的能力。蔡海霞等研究发现干旱胁迫显著降低高山柳 (Salixparaqplesia) 和沙棘 (Hippophaerhamnoides) 幼苗叶片的Chla、Chlb和Car含量[5]。李力等研究表明,持续高温干旱胁迫下3种北美红枫(AcerrubrumL.) 叶绿素相对含量降低[21]。已有研究证实,‘克罗莱卡’、‘皮削利’等油橄榄品种的Chla、Chlb和Car含量均随着干旱胁迫加剧而降低[22]。在本研究中,各品种油橄榄幼苗叶片Chla、Chlb和Car含量均随着干旱胁迫的加剧出现不同程度的降低,这与Ahmed[22]、Roussos等[23]的研究结果一致。分析原因主要是干旱胁迫致使油橄榄叶片水分减少,叶绿素分解加快且合成受阻,因而导致叶绿素含量下降;此外,活性氧积累、电解质外渗、渗透物质含量增加,抑制了光合色素活性。另外,研究还发现同一品种油橄榄叶片的Chla、Chlb和Car含量在干旱胁迫下下降趋势并不一致,且干旱胁迫期间品种间的Chla、Chlb和Car含量均存在显著差异,说明各种光合色素对干旱胁迫的响应程度不尽相同,不同品种油橄榄对干旱胁迫敏感性也不同。
3.3 油橄榄光合参数及水分利用效率对干旱胁迫的响应特征
光合作用是植物生长发育过程中物质和能量的来源,因此植物在干旱胁迫的逆境下光合参数会随干旱胁迫程度增加而变化。研究表明,植物在干旱胁迫状态下光合速率降低的幅度与干旱胁迫的程度和植物的耐旱能力密切相关[4, 11]。本研究中,各品种油橄榄幼苗的净光合速率、蒸腾速率、气孔导度和胞间二氧化碳浓度均呈现降低趋势,不同品种降低幅度不同且差异显著。综合各油橄榄品种光合参数表现发现,品种‘小苹果’耐旱性较强。干旱胁迫造成油橄榄光合作用下降原因主要包括叶绿体结构被破坏、叶绿体色素分解、气孔关闭、水分利用率降低等。植物Pn对干旱胁迫较为敏感,随着干旱胁迫时间延长, 其叶片气孔逐渐关闭,Pn和Tr均有所下降。同时,Gs降低是引起植物光合速率下降的重要原因之一。在逆境胁迫期间,若叶片Pn、Gs同时下降,光合作用下降的主要原因是气孔因素;随胁迫程度的增加和时间的延长,若Pn降低伴随着Ci不变或增加,则说明非气孔因素是光合作用下降主要原因[3, 8, 24]。从本试验中各品种油橄榄幼苗的光合参数均下降的表现可知,净光合速率的下降主要原因是气孔限制。
水分利用效率 (WUE) 一般是指植叶片Pn与Tr的比值,其代表某特定时间内,部分植物叶片的行为。植物受到干旱胁迫时,通常Tr下降速率较Pn更快,导致植物WUE升高[25]。但某些研究指出,胁迫超过一定程度后,光合系统的结构和功能受到破坏,Pn下降较Tr更快,此时WUE则有所降低[26]。在本试验中,持续干旱胁迫结束时,各品种油橄榄的WUE均上升,说明适度的干旱有利于油橄榄幼苗水分利用效率的提高,这与Boughalleb等[9]的研究结果一致。其中,品种‘小苹果’的水分利用效率呈现先降低后增加的趋势, 并在胁迫结束时显著高于其他品种,表明‘小苹果’具有较强的耐旱性。
总之,综合分析7个引进油橄榄品种幼苗在持续干旱胁迫下的光合指标发现, 各品种的叶片相对含水量、光合气体交换参数 (Pn、Tr、Gs和Ci)、光合色素 (Chla、Chlb和Car) 含量随干旱胁迫时间的延长均显著降低, 而WUE则持续上升,同时品种间光合特性表现差异达到显著水平。在干旱胁迫至后期时,各品种叶片均发生不同程度萎蔫,而品种‘小苹果’含水量显著高于其他品种,表现出更强的保水能力;在干旱胁迫结束时,‘小苹果’的Pn、Tr、Gs、Ci和WUE均显著高于其他品种,表现出较强的耐旱性。 因此,在供试油橄榄品种中‘小苹果’对干旱胁迫的适应性最强,适宜于在半干旱的川西地区种植。
[1] FARALONI C, CUTIONO I, PETRYCCELLI R,etal. Chlorophyll fluorescence technique as a rapid tool forinvitroscreening of olive cultivars (OleaeuropaeaL.) tolerant to drought stress[J].EnvironmentalandExperimentalBotany, 2011,73: 49-56.
[2] BOUSSADIA O, MARIEM F B, MECHRI B,etal. Response to drought of two olive tree cultivars (cv Koroneki and Meski)[J].ScientiaHorticulturae, 2008,116(4): 388-393.
[3] 张金政, 张起源, 孙国峰, 等. 干旱胁迫及复水对玉簪生长和光合作用的影响[J]. 草业学报, 2014,23(1):167-176.
ZHANG J Z, ZHANG Q Y, SUN G F,etal. Effects of drought stress and re-watering on growth and photosynthesis ofHosta[J].ActaPrataculturaeSinica, 2014,23(1): 167-176.
[4] 韩 博, 李志勇, 郭 浩, 等. 干旱胁迫下5种幼苗光合特性的研究[J]. 林业科学研究, 2014,27(1): 92-98.
HAN B, LI Z Y, GUO H,etal. Studies on seedling photosynthetic characteristics of five tree species under drought stress[J].ForestResearch, 2014,27(1): 92-98.
[5] 蔡海霞, 吴福忠, 杨万勤. 干旱胁迫对高山柳和沙棘幼苗光合生理特征的影响[J]. 生态学报, 2011,31(9):2 430-2 436.
CAI H X, WU F Z, YANG W Q. Effects of drought stress on the photosynthesis ofSalixparaqplesiaandHippophaerhamnoidesseedlings[J].ActaEcologicaSinica, 2011,31(9): 2 430-2 436.
[6] 王振兴, 陈 丽, 艾 军, 等. 不同干旱胁迫对山葡萄的光合作用和光系统II活性的影响[J]. 植物生理学报, 2014,50(8): 1 171-1 176.
WANG Z X, CHEN L, AI J,etal. Effects of different drought stress on photosynthesis and activity of photosystem II in leaves of amur grape (Vitisamurensis)[J].PlantPhysiologyJournal, 2014,50(8): 1 171-1 176.
[7] 李 倩, 王 明, 王雯雯, 等. 华山新麦草光合特性对干旱胁迫的响应[J]. 生态学报, 2012,32(13): 4 278-4 284.
LI Q, WANG M, WANG W W,etal. Response of photosynthetic characteristics ofPsathyrostachyshuashanicaKeng to drought stress[J].ActaEcologicaSinica, 2012,32(13): 4 278-4 284.
[8] 穆蓁蓁, 王一静, 艾合买提阿布都热依木, 等. 高温干旱对库尔勒香梨光合特性的影响[J]. 北方园艺, 2015,(6): 18-22.
MU Z Z, WANG Y J, A H M TABUDUREYIMU,etal. Effect of high temperature and drought on Korla Fragrant pear photosynthetic characteristics[J].NorthernHorticulture, 2015,(6): 18-22.
[9] BOUGHALLEB F, HAJLAOUI H. Physiological and anatomical changes induced by drought in two olive cultivars (cvZalmatiandChemlali)[J].ActaPhysiologiaePlantarum, 2011,33(1): 53-65.
[10] 孙善文, 章永江, 曹坤芳. 热带季雨林不同小生境大戟科植物幼树的叶片结构、耐旱性和光合能力之间的相关性[J]. 植物生态学报, 2014,38(4): 311-324.
SUN S W, ZHANG Y J, CAO K F. Correlations among leaf structure, drought tolerance and photosynthetic capacity in saplings ofEuphorbiaceaefrom different micro-habitats in a seasonal tropical rainforest[J].ChineseJournalofPlantEcology, 2014,38(4): 311-324.
[11] 陈春梅, 高聚林, 苏治军, 等. 玉米自交系吐丝期叶片光合参数与其耐旱性的关系[J]. 作物学报, 2014,40(9): 1 667-1 676.
CHEN C M, GAO J L, SU Z J,etal. Relationship between leaf photosynthetic parameters and drought resistance at silking stage in maize inbred lines[J].ActaAgronomicaSinica, 2014,40(9): 1 667-1 676.
[12] 陈炜青, 姜成英, 吴文俊, 等. 5个西班牙油橄榄品种苗木的年生长规律[J]. 经济林研究, 2015,33(3): 130-133.
CHEN W Q, JIANG C Y, WU W J,etal. Annual growth rhythm of seedlings of five introduced olive cultivars from Spain[J].NonwoodForestResearch, 2015,33(3): 130-133.
[13] CAVALHEIRO C V, PICOLOTO R S, CICHOSKI A J,etal. Olive leaves offer more than phenolic compounds - Fatty acids and mineral composition of varieties from Southern Brazil[J].IndustrialCropsandProducts, 2015,71: 122-127.
[15] 令 凡, 焦 健, 李朝周, 等. 不同油橄榄品种对低温胁迫的生理响应及抗寒性综合评价[J]. 西北植物学报, 2015,35(3): 508-515.
LING F, JIAO J, LI C Z,etal. Physiological response and comprehensive evaluation of cold resistance under cold stress for different varieties ofOleaeuropaea[J].ActaBot.Boreal.-Occident.Sin. 2015,35(3): 508-515.
[16] SOFO A, DICHIO B, XLLOYANNIS C,etal. Lipoxygenase activity and proline accumulation in leaves and roots of olive trees in response to drought stress[J].PhysiologiaPlantarum, 2004,121(1): 58-65.
[17] 陈淑燕, 毛培春, 田小霞, 等. 4种薰衣草属植物抗旱性综合评价[J]. 干旱地区农业研究, 2013,31(6): 152-158.
CHEN S Y, MAO P C, TIAN X X,etal. Comprehensive evaluation of drought resistance of four lavender species[J].AgriculturalResearchintheAridAreas, 2013,31(6): 152-158.
[18] 陈文荣, 曾玮玮, 李云霞, 等. 高丛蓝莓对干旱胁迫的生理响应及其抗旱性综合评价[J]. 园艺学报, 2012,39(4): 637-646.
CHEN W R, ZENG W W, LI Y X,etal. The physiological responds of highbush blueberry to drought stress and the comprehensive evaluation on their drought resistance capacity[J].ActaHorticulturaeSinica, 2012,39(4): 637-646.
[19] 汪 毅, 郭海林, 陈静波, 等. 国审品种‘苏植1号’杂交结缕草抗旱性初步评价与分析[J]. 草业学报, 2016,25(5): 30-39.
WANG Y, GUO H L, CHEN J B,etal. Preliminary evaluation of drought resistance for the hybrid zoysiagrass ‘Suzhi 1’[J].ActaPrataculturaeSinica, 2016,25(5): 30-39.
[20] GUERFEL M, BACCOURI O, BOUINAH D,etal. Impacts of water stress on gas exchange, water relations, chlorophyll content and leaf structure in the two main Tunisian olive (OleaeuropaeaL.) cultivars[J].ScientiaHorticulturae, 2009,119(3): 257-263.
[21] 李 力, 刘玉民, 王 敏, 等. 3种北美红枫对持续高温干旱胁迫的生理响应机制[J]. 生态学报, 2014,34(22): 6 471-6 480.
LI L, LIU Y M, WANG M,etal. Physiological response mechanism of three kinds ofAcerrubrumL. under continuous high temperature and drought stress[J].ActaEcologicaSinica, 2014,34(22): 6 471-6 480.
[22] AHMED C B, ROUINA B B, SENSOY S,etal. Changes in gas exchange, proline accumulation and antioxidative enzyme activities in three olive cultivars under contrasting water availability regimes[J].EnvironmentalandEperimentalBotany, 2009,67(2): 345-352.
[23] ROUSSOS P A, DENAXA N, DAMVAKARIS T,etal. Effect of alleviating products with different mode of action on physiology and yield of olive under drought[J].ScientiaHorticulturae, 2010,125(4): 700-711.
[24] 徐艳平, 胡翠美, 张文会, 等. 干旱胁迫对野生大豆幼苗光合作用相关指标的影响[J]. 大豆科学, 2013,32(3): 341-344, 348.
XU Y P, HU C M, ZHANG W H,etal. Effect of simulated drought stress on photosynthesis related indexes at seedling stage of wild soybeans[J].SoybeanScience. 2013,32(3): 341-344, 348.
[25] 张 怡, 罗晓芳, 沈应柏. 干旱胁迫下四倍体刺槐幼苗水分利用效率及稳定碳同位素组成的研究[J]. 西北植物学报, 2009,29(7): 1 460-1 464.
ZHANG Y, LUO X F, SHEN Y B. Water use efficiency and carbon isotopic compositions in tetraploid clones of black locust under drought resistance[J].ActaBot.Boreal.-Occident.Sin. 2009,29(7): 1 460-1 464.
[26] 张学霞, 杨璐璐, 华 开. CO2浓度对高羊茅抗旱性及水分利用效率的影响分析[J]. 草地学报, 2015,23(3): 502-509.
ZHANG X X, YANG L L, HUA K. Effects of CO2concentration on tall fescue drought resistance and water use efficiency[J].ActaPrataculturaeSinica, 2015,23(3): 502-509.
(编辑:裴阿卫)
Photosynthetic Characters of Different Introduced Olive Varieties under Drought Stress
LAN Yan1, XU Yingjie1, TAN Feng1, DING Chunbang2, LI Tian1, 2*, SU Guangcan3
(1 Agricultural College, Sichuan Agricultural University, Wenjiang, Sichuan 611130, China; 2 College of Life Sciences,Sichuan Agricultural University,Yaan, Sichuan 625014, China;3 Liangshan Zhongze New Tech Development Co.,Ltd., Xichang, Sichuan 6150004, China)
Olive (OleaeuropaeaL.) is one of the four famous woody oil species across the world with important nutritional and pharmaceutical values. In order to reveal the relationship between drought resistance and photosynthesis characteristics, we conducted pot experiments to investigate the effect of drought stress on the photosynthetic characteristics of seven introduced olive varieties including Picholine, Picual, Manzanillo, Koroneiki, Frantoio, Arbequina and Coratina, which had been filtered the suitable for growth in the semi-arid region of the Southwest of Sichuan Province. The results showed that: (1) the leaf relative water content (LRWC) of all the tested olive varieties was significantly reduced with the intensification of the degree of drought stress (P<0.05); At the later period of stress (25 days), the cultivars showed different levels of wilting symptom compared to control. Among them, Coratina showed the highest water loss rate while Manzanillo slightly affected, indicting the Manzanillo leaves exhibited a higher water retaining capacity compared to other cultivars under drought stress. (2) The concentrations of photosynthetic pigment in all the tested olive varieties were significantly decreased with the increasing duration of drought stress (P<0.05), which was mainly resulted from the rapid degradation of chlorophyll than biosynthesis; the Chla and Chlb contents in Arbequina, as well as Car content in Picholine were significantly declined after 25 d drought stress (P<0.05), while the Chla content of Manzanillo was not decreased obviously (P>0.05). (3) The photosynthetic parameters of the cultivars varied in response to drought stress. Net photosynthetic rate (Pn), transpiration rate (Tr), stomatal conductance (Gs) and intercellular CO2concentration (Ci) in leaves decreased, while the water use efficiency (WUE) increased with the drought. In addition, the drop ofPn,TrandCiof Frantoio andGsof Picholine were higher than those of other varieties, while slightly decreased inPn,GsandCiand sharply increased inWUEwas observed in Manzanillo. Our research showed that under the persistent drought stress the olive seedling leaves were substantial loss of water and the photosynthetic pigment structure was destroyed and decomposed, as well as stomatals closed and photosynthesis decreased. Among the tested olive varieties, Manzanillo shows excellent adaptability to drought stress and is suitable for planting in the semi-arid region of the Southwest of Sichuan Province.
drought stress;OleaeuropaeaL.; photosynthetic character; photosynthetic pigment
1000-4025(2016)11-2247-09
10.7606/j.issn.1000-4025.2016.11.2247
2016-07-15;修改稿收到日期:2016-10-10
四川省科技厅科技支撑计划项目(2014NZ0103)
兰 艳(1992-),女,在读硕士研究生,主要从事作物高产优质栽培研究。E-mail: lanyansicau@163.com
*通信作者:李天,教授,博士生导师,主要从事作物高产优质与生态环境的关系研究。E-mail: lit@sicau.edu.cn
Q945.78;S565.7
A
猜你喜欢
汽车观察(2020年4期)2020-05-27
汽车观察(2019年9期)2019-10-22
汽车观察(2019年4期)2019-07-01
世界汽车(2019年2期)2019-03-01
中成药(2018年4期)2018-04-26
中国果业信息(2017年8期)2017-09-12
中国农业信息(2016年16期)2016-02-06
园艺与种苗(2015年7期)2015-12-09
绿色科技(2015年5期)2015-07-05
中国洗涤用品工业(2015年8期)2015-02-28
