
Metridia荧光素酶检测条件的优化
2016-12-09 09:09郭志兰车路阳李晶哲孙震晓刘长振
癌变·畸变·突变 2016年6期
郭志兰,车路阳,李晶哲,吴 双,孙震晓,*,刘长振,*
(1.北 京中医药大学中药学院,北京 100102;2.中国中医科学院医学实验中心中医药防治重大疾病基础研究北京市重点实验室,北京 100700;3.中国人民解放军总医院骨科,北京 100853)
Metridia荧光素酶检测条件的优化
郭志兰1,2,车路阳3,李晶哲2,吴 双1,孙震晓1,*,刘长振2,*
(1.北 京中医药大学中药学院,北京 100102;2.中国中医科学院医学实验中心中医药防治重大疾病基础研究北京市重点实验室,北京 100700;3.中国人民解放军总医院骨科,北京 100853)
生物发光是自然界普遍存在的一种自然现象,在细菌、真菌、原生动物、腔肠动物、昆虫和鱼等生物中均存在生物发光现象[1]。生物发光本质是荧光素酶催化其底物荧光素发光的生物过程,荧光素酶是一种氧化酶,在有氧气存在的情况下,催化其各自的底物氧化产生生物光[2-3],一些荧光素酶发挥作用还需要ATP及Mg2+的辅助[4]。
根据荧光素酶表达的位置可将其分为胞质型和分泌型。科研人员在克隆了分泌型荧光素酶的基因后,将其作为报告基因广泛运用到科研及医疗等领域,如
对细胞内基因转录情况[5]、体内蛋白质的代谢分布情况[6]等的实时监测,对信号通路转导、病毒的复制过程及传染机制等的研究[7-8],对肿瘤增殖情况及药物对肿瘤治疗效果的判断[9]及高通量药物的筛选[10-11]等等。
Metridia荧光素酶(Metridia luciferase,MLuc)来源于海洋桡足动物Metridia longa,可与特异性底物腔肠素(coelenterazine,CTZ)反应产生蓝光(λmax=480 nm)[13],是一种热稳定性高的分泌型荧光素酶。Markova等[12]在2004年从Metridia longa中首次克隆出了MLuc的cDNA,全长897 bp,其中开放阅读框656 bp,编码相对分子质量2.39×104的氨基酸多肽。之后研究人员对MLuc尝试了一系列的改造,如Takenaka等[13]克隆出了野生型MLuc的两种相对分子质量较小的变体,分别为2.27×104和2.03×104;Markova等[14]又于2012年报道了在N末端截去一些氨基酸序列的高活性MLuc 变体,接着又在2015年报道了目前最小的,相对分子质量为1.65×104的高活性MLuc变体[15]。
本实验室在使用MLuc作为报告基因的过程中发现,MLuc酶的发光反应受环境影响较大,对结果的稳定性、可靠性造成了较大的影响,而并未查阅到相关报道的文献,故本实验室在不同检测条件下对其发光强度及稳定性进行了初步比较,并尝试摸索其合适的检测条件。
1 材料与方法
1.1 材料与试剂
感受态大肠杆菌E.coli DH5α购于天根生化科技(北京)有限公司;人胚肾上皮包装细胞GP2-293、人宫颈癌细胞系HeLa由本实验室保存;pQCXIP载体购于Clontech公司;NF-κB序列、MLuc序列由北京普尔普乐生物科技有限公司合成。限制性内切酶、连接酶购于Fermentas公司;腔肠素购于Sigma公司;质粒小提试剂盒、琼脂糖凝胶回收试剂盒均购于天根生化科技(北京)有限公司;Ready-To-GlowTMSecreted Luciferase Reporter System试剂盒购于Clontech公司;DMEM培养基购于Gibco公司、胎牛血清购于HyClone公司。
1.2 方法
1.2.1 pQCXIP-NF-κB-MLuc的构建及其鉴定 首先将逆转录病毒载体pQCXIP利用Bgl II、Age I进行双酶切,去除pQCXIP原有CMV启动子,然后将合成的含有转录因子NF-κB增强子和TATA box序列(序列来源于Clontech公司,两端含有Bgl II和Age I的酶切位点)的DNA片段经Bgl II、Age I双酶切装载入改造后的pQCXIP质粒上。MLuc(序列来源于Clontech公司,两端含有Age I和EcoR I的酶切位点)与上述得到的pQCXIPNF-κB质粒分别用Age I、EcoR I进行双酶切,连接并转化获得重组的逆转录病毒载体pQCXIP-NF-κBMLuc。本实验合成的MLuc序列的N端部分含有分泌信号肽,可帮助蛋白分泌到细胞外的培养基中。
1.2.2 MLuc酶的制备 利用阳离子脂质体转染法将构建的pQCXIP-NF-κB-MLuc质粒瞬时转染到人的宫颈癌细胞系HeLa中进行表达,其中质粒与lipo2000的质量体积比为1∶2,转染后仍使用低血清培养基Opti-MEM进行培养。48h后,收集培养基上清,离心去除残留的细胞碎片,分装,-80℃ 保存。
1.2.3 MLuc酶的检测 在常规培养基或缓冲液中加入底物CTZ至相应浓度,按每孔100μL加至黑色96孔板中,然后加入制备的MLuc酶或含MLuc酶的细胞上清10μL,或直接在含分泌的MLuc酶的培养基上清中添加CTZ至相应浓度,充分混匀,于化学发光仪(BioTek Synergy 2)中进行相对发光单位 (relative luminescence unit,RLU)值读取。
1.2.4 PBS+0.02% NP-40缓冲液与Clontech试剂盒检测灵敏度的比较 取上述制备的MLuc酶,先用培养基(RPMI 1640+10%胎牛血清,R10)稀释10倍作为母液,再依次4、16、64、256、1 024倍梯度稀释,各取100μL,加入10μL添加底物CTZ(30μmol/L)的反应buffer(Ready-To-GlowTMSecreted Luciferase Reporter System,Clontech),充分混匀后检测RLU值;取上述制备的MLuc酶作为母液,用R10培养基4、16、64、256、1 024倍梯度稀释,各取10μL,再加入100μL添加底物CTZ(3μmol/L)的PBS+0.02% NP-40缓冲液,充分混匀后检测。
2 结果
2.1 pQCXIP-NF-κB-MLuc的构建及其鉴定
利用Bgl II、Age I双酶切鉴定表明(图1A),双酶切出来的核酸条带与理论插入的NF-κB增强子/TATA box序列条带大小吻合,说明已构建获得重组质粒pQCXIP-NF-κB。随后将MLuc与重组质粒pQCXIPNF-κB经Age I、EcoR I双酶切、连接和转化,并经Bgl II、EcoR I双酶切鉴定(图1B),结果表明MLuc序列被成功插入pQCXIP-NF-κB中,获得了目标质粒pQCXIP- NF-κB-MLuc。该质粒经测序证明序列完全正确。
2.2 MLuc酶检测过程中的影响因素
2.2.1 pH的影响 调节培养基(RPMI 1640+10%胎牛
血清,R10)的pH,检测pH从5.0到9.0范围内MLuc与底物CTZ反应的发光强度,结果如图2A,pH的波动对发光值大小的影响非常明显。pH在6.5时RLU值最大,7~7.5之间较平稳,而在pH<6或>8时,其RLU值大幅下降。
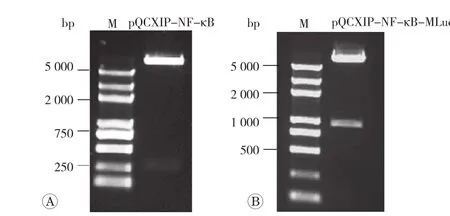
图1 pQCXIP-NF-κB-Mluc的构建及鉴定
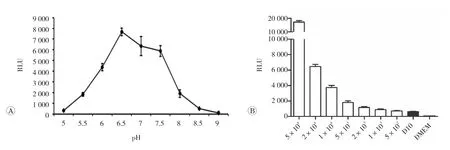
图2 培养基对MLuc酶发光反应的影响
2.2.2 血清的影响 将瞬时转染pQCXIP-NF-κB-MLuc 24h后的HeLa细胞分别以每孔5×104、2×104、1×104、5×103、2×103、1×103、500个/孔的量接种于96孔板中,每组5个复孔,培养24h后,取培养基上清进行RLU值的检测,结果如图2B,在每孔5×103~5×104个细胞时,MLuc酶的表达量与细胞数之间呈现良好的线性关系,而≤2×103时无明显差异。各组上清经精密pH试纸验证,几乎无差别,故排除pH的影响。而同样检测条件下,取HeLa培养基(DMEM培养基加10%的血清,D10)及DMEM培养基进行RLU值的检测,结果含血清的培养基其RLU值为609±61,与按每孔5×102个细胞接种的上清RLU值已相近,而DMEM仅为28± 4,也就意味着是培养基中的血清带来检测背景。
2.3 MLuc酶检测条件的优化
2.3.1 缓冲液的选择 我们选择了可用于细胞实验的Tris、Hepes、PBS 3种缓冲液,pH均调为7.4。在含有底物CTZ(3μmol/L)的3种缓冲液(100μL)中,分别加入上述制备的MLuc酶10μL,充分混匀后,于发光检测仪中检测,结果如图3,可见相比于Tris及Hepes,PBS缓冲液无论是在发光强度及稳定性方面都具有明显的优势。
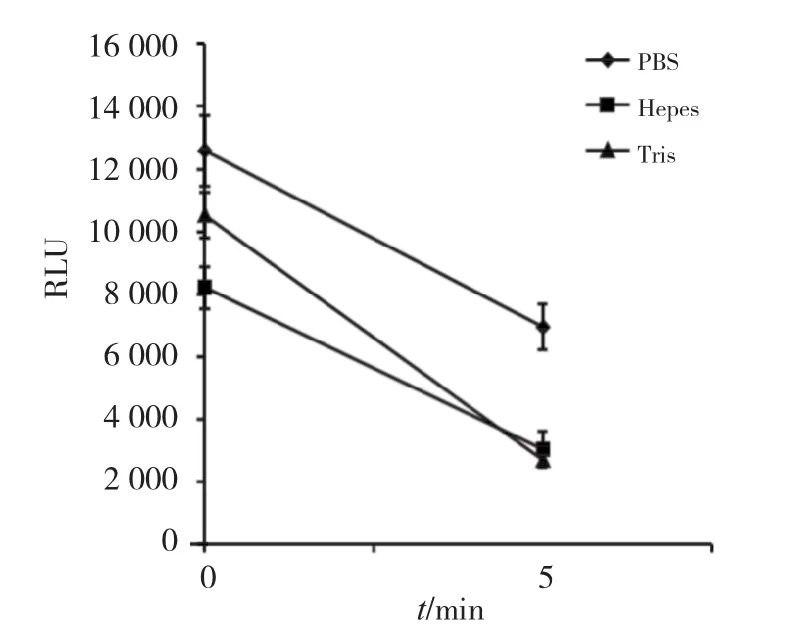
图3 MLuc在不同缓冲液中的发光情况
2.3.2 表面活性剂的影响 在确定了选用PBS缓冲液后,我们尝试通过添加表面活性剂来进一步改善c
mLuc酶发光的性能。在比较了几种常用的及不常用的表面活性剂后,我们发现非离子型表面活性剂NP-40会对MLuc酶的发光反应产生较大影响,如图4,在PBS中添加0.01% NP-40(V/V)可显著增加其发光强度,增加NP-40浓度至0.05%可显著增加其稳定性,但同时使其RLU值略有降低。此外,添加0.01%及0.02% NP-40几乎不会产生背景(RLU值在20附近),而添加0.03% ~0.05% NP-40则使检测背景略有提高(RLU值在200附近)。

图4 NP-40对MLuc发光性能的影响
2.4 PBS+0.02% NP-40与Clontech缓冲液检测灵敏度的比较
在同样MLuc浓度及底物浓度下,比较Clontech缓冲液与PBS+0.02% NP-40能够检到的最低酶量,结果如图5,PBS+0.02% NP-40缓冲液在上述制备的MLuc稀释2 560倍时依然与其RLU值呈现较好的线性关系,而Clontech缓冲液在稀释160倍后其RLU值开始趋于平缓,无法体现MLuc浓度的差异。
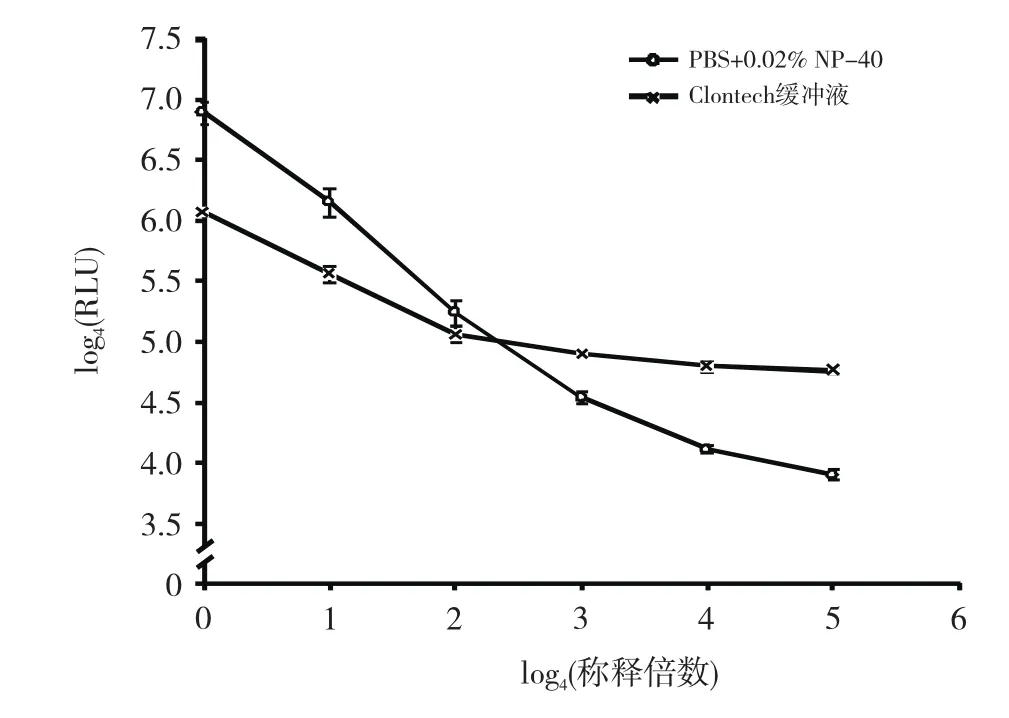
图5 PBS+0.02% NP-40与Clontech缓冲液检测灵敏度比较
3 讨 论
培养基在细胞培养过程及储存过程中,由于与空气的接触,细胞增殖代谢等原因,pH会不可避免地发生变化,pH在6~8范围内的波动很常见,这就意味着会对MLuc酶发光值大小产生很大影响,从而影响实验结果的判断。而pH变化对MLuc的RLU值影响可能是因为pH对MLuc酶的活性产生较大影响,从而影响其与CTZ反应的发光值,当然也不排除pH会影响底物CTZ活性的可能。
针对血清也会使底物CTZ发光的现象,猜测其原因可能是血清中含丰富的蛋白质及多肽等,其某些氨基酸也可以与底物CTZ相应位点结合产生类似于MLuc酶的功效。Vassel等[16]对白蛋白能使CTZ化学发光的研究部分解释了我们的实验结果,我们认为血清中肯定还有其他成分也有类似的作用,还需进一步研究。对于一些比较小的体系,如小量接种的96孔板,甚至单细胞体系,血清带来的背景,将会导致对实际表达酶量及各组之间酶量比例关系判断产生很大误差;此外,对于一些不太明显的组间差异,很有可能会因为血清带来的相对较大的背景而被忽略。也就是说,血清带来的背景影响使MLuc报告基因本身的准确性及灵敏都降低了。
在发现上述MLuc酶活检测过程中存在的问题后,我们开始验证猜想并尝试找寻解决方案。如图2所示,MLuc酶在不同pH条件下RLU值波动明显,培养基的血清导致检测背景。考虑到上述问题集中在MLuc酶所在的环境中——培养基,故我们考虑更换MLuc酶的检测环境,选用一种缓冲液来减弱由于培养基的pH、所含血清等带来的一系列问题。如图3及图4所示,在比较了几种常用的缓冲体系及添加NP-40后,本研究最终选定了添加0.02%(V/V)NP-40的PBS缓冲体系,因该体系能较好地避免上述培养基的弊端,具有稳定的pH检测环境、检测背景低,而且MLuc酶与CTZ反应的发光值也相对稳定。该本系与目前市场上Clontech生产的针对MLuc酶检测的试剂盒相比,灵敏度更高(如图5所示)。由此,我们认为添加0.02% NP-40的PBS缓冲液更适合于对MLuc酶的检测。此外,除了本研究中所涉及的MLuc酶,其他荧光素酶等报告基因也存在类似的检测问题,通过类似本研究的调整方式也可进行检测方式的优化,更灵敏且准确地体现所研究的相应转录因子的含量或变化量 ,从而更好地对相应信号通路的变化情况作出判断。
[1]markova SV,Vysotski ES.Coelenterazine-dependent luciferases[ J].B iochemistry (Moscow),2015,80(6):714-732.
[2]hastings JW.Chemistries and colors of bioluminescent reactions:a review[J].Gene,1996,173(1):5-11.
[3] Wilson T,Hastings JW.Bioluminescence[J] .Annu Rev Cell Dev Biol,1998,14:197-230.
[4] Badr CE,Tannous BA.Bioluminescence imaging:progress and applications[J].Trends Biotechnol,2011,29(12):624-633.
[5] Lupold SE,Johnson T,Chowdhury WH,etal.A real timemetridia luciferase based non-invasive reporter assay ofmammalian cell viability and cytotoxicity via the β-actin promoter and enhancer[J].PLoS One,2012,7(5):e36535.
[6] EL-Amouri SS,Cao P,Miao C,etal.Secreted luciferase for in vivo evaluationof systemic protein delivery inmice[J].Mol Biotechnol,2013,53(1):63-73.
[7]haugwitzm,Nourzaie O,Garachtchenko T,etal.Multiplexing bioluminescentand fluorescent reporters tomonitor live cells[J].Curr Chem Genomics,2008,1,11-19.
[8]msaouel P,Dispenzieri A,Galanis E.Clinical testing of engineered oncolyticmeasles virus strainsin the treatmentof cancer:anoverview[J].Curropinmol Ther,2009,11(1):43-53.
[9] BovenbergmS,DegelingmH,Hejazi S,etal.Multiplex blood reporters for simultaneousmonitoring of cellular processes[J].Anal Chem,2013,85(21):10205-10210.
[10] Orlando RA,Gonzales AM,Hunsaker LA,etal.Inhibitionof nuclear factor κB activation and cyclooxygenase-2 expression by aqueous extracts ofhispanicmedicinalherbs[J].Jmed Food,2010,13(4):888-895.
[11] Cheung L,Yu DM,Neiron Z,etal.Identificationof newmRP4 inhibitors from a library of FDA approved drugs using ahigh-throughput bioluminescence screen[J].Biochem Pharmacol,2015,93(3):380-388.
[12]markova SV,Golz S,Frank LA,etal.Cloning and expressionof cDNA for a luciferase from themarine copepodmetridia longa.A novel secreted bioluminescent reporter enzyme[J].J Biol Chem,2004,279(5):3212-3217.
[13] Takenaka Y,Masudah,Yamaguchi A,etal.Twoforms of secreted and thermostable luciferases from themarine copepod crustacean,Metridia pacifica[J].Gene,2008,425(1/2):28-35.
[14]markova SV,Burakova LP,Vysotski ES.High-active truncated luciferase of copepodmetridia longa[J].Biochem Biophys Res Commun,2012,417(1):98-103.
[15]markova SV,LarionovamD,Burakova LP,etal.The smallest naturalhigh-active luciferase:cloning and characterizationof novel 16.5-kDa luciferase from copepodmetridia longa[J].Biochem Biophys Res Commun,2015,457(1):77-82.
[16] Vassel N,Cox CD,Naseem R,etal.Enzymatic activity of albumin shown by coelenterazine chemiluminescence[J].Luminescence,2012,27(3):234-241.
Amodified assay system formetridia luciferase
GUO Zhilan1,2,CHE Luyang3,LI Jingzhe2,WU Shuang1,SUN Zhenxiao1,*,LIU Changzhen2,*
(1.College of Chinese Pharmacy, Beijing University of Chinesemedicine, Beijing 100102; 2.Beijing Key Laboratory of Research of Chinesemedicine on Prevention and Treatment formajor Diseases, Experimental Research Center, China Academy of Chinesemedical Sciences, Beijing 100700; 3.Departmentof Orthopedics, Chinese People’s Liberation Army Generalhospital, Beijing 100853, China)
目的: 探讨培养基pH、血清对Metridia荧光素酶(MLuc)检测的影响并尝试找到一种优化的检测条件。方法:构建表达MLuc的逆转录病毒载体,瞬时转染人宫颈癌细胞系HeLa,制备收获MLuc;利用化学发光仪检测不同条件下的培养基对MLuc发光性能的影响,并尝试通过不同缓冲液与试剂的组合代替培养基来获得最佳条件。结果:培养基pH变化导致MLuc发光值产生较大波动,血清的存在会导致检测背景;添加0.02%(V/V)NP-40的磷酸盐缓冲液(PBS)(pH=7.4)作为MLuc检测的缓冲体系代替培养基,其发光值大小相当,检测背景值由原来的600左右降低到20左右。结论:以添加0.02% NP-40的PBS作为MLuc检测的缓冲体系可消除培养基pH、血清的影响,使检测结果更加准确和灵敏。
荧光素酶;pH;血清;检测;优化
OBJECTIVE:To investigate whether the pH and serum of cell culturemedium would affect themetridia luciferase (MLuc) assay or not,and to explore amodified assaymethod formLuc.METHODS:A recombinant retrovirus vector which expressedmLuc was constructed and then transiently transfected intohuman cervical cancer cell lineheLa to producemLuc.Then,the effects of cell culturemedium (e.g.buffers combined with reagents) on bioluminescence ofmLuc were assayed.RESULTS:Various pH levels of cell culturemedium caused great fluctuationof the bioluminescence intensity ofmLuc.In addition,serum in the cell culturemedium would alter the background ofmLuc.When the cell culturemedium was replaced by phosphate buffered saline (PBS) supplemented with 0.02% (V/V) NP-40 (pH=7.4),the bioluminescence intensity between them was comparable.However,the background level ofmLuc was decreased from 600 to 20.CONCLUSION:Using PBS supplemented with 0.02% NP-40 as an assay buffer formLuc,the side effects from pH and serum of cell culturemedium duringmLuc assay would be reduce and applicationofmLucin research can be improved.
luciferase;pH;serum;assay;modify
Q819
A
1004-616X(2016)06-0438-05
1 0.3969/j.issn.1004-616x.2016.06.005
2016-08-28;
2016-10-31
国家自然科学基金(31170829,81171762,81550017,81473418);中国中医科学院自主选题项目(ZZ2015015)
作者信息:郭志兰,E-mail:gzl20141019@163.com。*通信作者,刘长振,E-mail:lcz0220@163.com;孙震晓,E-mail:s unzxcn@hotmail.c om
猜你喜欢
天津医科大学学报(2021年4期)2021-08-21
中日友好医院学报(2021年1期)2021-04-14
科学(2020年2期)2020-08-24
湖北大学学报(自然科学版)(2020年4期)2020-07-15
山东医药(2020年9期)2020-05-20
山东化工(2019年18期)2019-10-16
科技资讯(2017年12期)2017-06-09
天津科技大学学报(2016年3期)2016-08-02
哈尔滨商业大学学报(自然科学版)(2016年1期)2016-04-22
中国医药导报(2015年27期)2015-02-28
