
云顶山亚高山草甸植被分布、物种多样性与土壤化学因子的相关性
2016-11-10 03:16杜京旗张巧仙田晓东许馨云袁自强
植物研究 2016年3期
杜京旗 张巧仙* 田晓东 王 悦 许馨云 宋 丽 袁自强
(1.吕梁学院植物化学研究所,吕梁 033000; 2.草地农业系统国家重点实验室,兰州 730000)
* 通信作者:443481677@qq.com
云顶山亚高山草甸植被分布、物种多样性与土壤化学因子的相关性
杜京旗1张巧仙1*田晓东1王 悦1许馨云1宋 丽1袁自强2
(1.吕梁学院植物化学研究所,吕梁 033000;2.草地农业系统国家重点实验室,兰州 730000)
根据36个样地27个种的野外调查数据,运用双向指示种分析(TWINSPAN)将云顶山亚高山草甸植被分为蒲公英+钝裂银莲花+车前群丛、委陵菜+鳞叶龙胆+蒲公英+车前群丛、蒲公英+委陵菜+百里香群丛、车前+委陵菜+百里香+白莲蒿群丛、车前+白莲蒿+委陵菜群丛和蒲公英+羊茅+车前群丛。运用典范对应分析法(CCA)分析了pH、全氮(TN)、全磷(TP)、K+、Cl-、Ca2+、Mg2+、Zn2+、Fe、有机质(OM)10个土壤化学因子和云顶山亚高山草甸植被分布的相关关系。CCA结果表明,TN、Ca2+、Mg2+、Fe是影响云顶山亚高山草甸植被分布的主要土壤化学因子,6个群丛分别对应不同的土壤生境。土壤化学因子与物种多样性指数的相关分析表明,TN、Ca2+与Shannon-Weiner指数H呈正相关;TN、Mg2+与丰富度指数R分别呈正相关、负相关;K+与R、H、均匀度指数E均呈正相关。因此,物种多样性的保护应结合当地的土壤特点。
物种多样性;分布;TWINSPAN;CCA;土壤化学因子
群落与环境因子之间的关系是生态学的一个重要研究内容[1~3],土壤因子与植被多样性之间的关系是其中一个研究热点[4~6]。土壤是影响植物种子萌发、生长与发育的重要生态因子[7~8],是生态系统中矿质营养的储存场所,另外,土壤空间异质性会影响群落的演替过程[9]。土壤的理化性质、生物性质会影响植被多样性。不同的土壤条件下,群落的物种多样性不同。群落中的大气候因素相似,则群落中土壤理化性质不同是物种多样性形成的一个重要原因。土壤的物理性质有:组成、质地、结构、水分、温度、空气;土壤的化学性质包括pH、盐分、有机质、矿质元素。与土壤的物理性质相比,土壤的化学性质与植被分布、物种多样性的关系更为密切[10~12]。
云顶山亚高山草甸是天然优质草场,然而,随着旅游热的升温和放牧强度的增加,云顶山的植被遭到了破坏,与亚高山草甸的经济价值相比,其净化环境、休憩旅游等服务功能价值更高。与云顶山相关的研究包括:虎榛子(Ostryopsisdavidiana)群落的群落学特征[13]、云顶山植被的垂直分布特征[14]、植被群落生态分析[15]、格局分析[16]、优势种群的生态位[17]、干扰对多样性的影响[18]、野生资源研究[19~20]、植被群落的排序[21]。未见土壤理化性质与植被分布、物种多样性相关性的相关报道。
本研究运用植物群落数量分类和典范对应分析,定量研究了土壤化学因子对云顶山亚高山草甸的影响,确定了影响植被分布和物种多样性的主要化学因子,进一步揭示了草甸群落特征以及植被分布与生态因子之间的密切关系,为云顶山植物的保护及可持续利用提供了理论依据。
1 研究区概况
云顶山位于山西省的方山县、娄烦县与交城县的交界处(37°50′49″N~38°02′15″N,111°30′33″E~111°47′11″E),南与庞泉沟相连,于2002年成为省级自然保护区,生境复杂,动植物种类较多。因海拔高(最高处海拔为2 708 m),顶入云端而得名,面积有23 863 hm2。
云顶山属大陆性季风气候,年均温7℃,年均降水量为700 mm,雨季在6~9月,植被类型主要为落叶阔叶林、针叶林、亚高山草甸,土壤类型包括棕色森林土、亚高山草甸土、褐色土。常见草本植物有车前(Plantagoasiatica)、委陵菜(Potentillachinensis)、蒲公英(Taraxacummongolicum)等。
2 实验设计
2.1 野外取样
2012、2013年的8月下旬分别进行野外取样(海拔2 622~2 627 m)。云顶山存在放牧和旅游开发等干扰,因此,设置样地时,先充分了解了研究地区的植被环境、放牧情况、旅游开发等人为活动,在没有人为因素的干扰或人为因素干扰少的地区取样。取样采用样带法,设置三条南北平行样带,间距为500 m,每条样带上每隔250 m设置5 m×5 m的样地,每个样地设置三个1 m×1 m的样方,样地的个数为36个。每个草本样地记录物种数,物种名称;测量株高,密度的计测采用直接计数法,投影盖度和多度的计测采用目测估计法。共记录35个物种,将频度<5%的偶见种、稀有种除去,得到27个物种。在植物取样的同时进行土样的采集:采用S型取样法(设置5个取样点,每个取样点3次重复)在每个草本样方0~20 cm处取土样,并将同一样地中的土样均匀混合,用作土壤指标的测定。
2.2 数据采集
pH值采用水土比5∶1的浸出液进行测定(电位法),Zn2+、Mg2+、K+、Ca2+、Fe采用原子吸收仪进行测定,OM采用高温电炉灼烧法,采用AgNO3滴定法测定Cl-的含量,TP采用H2SO4-HClO4-钼锑抗比色法,TN用凯氏定氮法测定[22]。
2.3 数据分析
①物种优势度用重要值来衡量。

(1)
②数量分类:使用Win TWINS软件对样地—重要值矩阵(36×27)进行数量分类。
③排序:使用Canoco for Windows 4.5软件对样地—重要值矩阵和样地—环境因子矩阵(36×10)进行典范对应分析(CCA),使用CanoDraw for Windows绘制CCA排序图。
④物种多样性相关指数:
丰富度指数:R=S
(2)
式中:S为样地所包含的物种数。
Shannon-Wiener指数:

(3)
式中:Pi为重要值。
均匀度指数:
E=H/lnS
(4)
⑤数据整理使用Microsoft Excel 2007软件,绘图使用Origin 8.0软件,不同群丛的土壤化学因子,多样性指数的方差分析、多重比较(LSD),物种多样性指数(R、H、E)与土壤因子之间的Spearman
相关性分析,多样性指数之间的Pearman相关性分析以及土壤因子之间的Pearman相关性分析使用SPSS 17.0软件。
3 实验结果
3.1 TWINSPAN分类结果
用云顶山亚高山草甸的样地—重要值矩阵(样地数量:36,物种数量:27)的分类结果(图1),同时结合调查实际,得到6个不同的群丛,群丛的命名依据:优势种和TWINSPAN划分的指示种。

图1 TWINSPAN分类结果矩阵图 数字代表重要值等级Fig.1 The classification matrix of TWINSPAN Figures represent important value levels.
(1)群丛Ⅰ。蒲公英+钝裂银莲花+车前(Ass.Taraxacummongolicum+Anemoneobtusiloba+Plantagoasiatica)。群落中蒲公英密度27±10株·m-2(平均值±标准差),盖度33%±15%;钝裂银莲花密度198±101株·m-2,盖度43%±19%;车前密度16±10株·m-2,盖度4%±6%。群落中主要的伴生种有:北乌头、水杨梅(盖度小于5%)。
(2)群丛Ⅱ。委陵菜+鳞叶龙胆+蒲公英+车前(Ass.Potentillachinensis+Gentianasquarrosa+Taraxacummongolicum+Plantagoasiatica)。群落中鳞叶龙胆密度77±20株·m-2,盖度22%±13%;委陵菜密度98±39株·m-2,盖度7%±9%;车前密度33±11株·m-2,盖度5%±7%;蒲公英密度16±10株·m-2,盖度7%±5%。群落中主要的伴生种有:蒿草、东方草莓,盖度均小于5%。
(3)群丛Ⅲ。蒲公英+委陵菜+百里香(Ass.Taraxacummongolicum+Potentillachinensis+Thymusmongolicus)。群落中委陵菜密度74±37株·m-2,盖度9%±6%;蒲公英密度63±22株·m-2,盖度54%±30%;百里香密度7±11株·m-2,盖度5%±2%。群落中主要的伴生种有:车前、北乌头、老鹳草(盖度小于5%)。
(4)群丛Ⅳ。车前+委陵菜+百里香+白莲蒿(Ass.Plantagoasiatica+Potentillachinensis+Thymusmongolicus+Artemisiasacrorum)。群落中委陵菜密度85±69株·m-2,盖度10%±7%;车前密度71±35株·m-2,盖度7%±2%;百里香密度42±28株·m-2,盖度17%±13%;白莲蒿密度22±12株·m-2,盖度8%±6%。群落中主要的伴生种有:平车前(盖度小于5%)、牛尾蒿。
(5)群丛Ⅴ。车前+白莲蒿+委陵菜(Ass.Plantagoasiatica+Artemisiasacrorum+Potentillachinensis)。群落中车前密度62±43株·m-2,盖度9%±6%;委陵菜密度124±86株·m-2,盖度18%±18%;白莲蒿密度10±11株·m-2,盖度5%±6%。群落中主要的伴生种有:披针薹草(盖度小于5%)、蒲公英和小红菊(盖度小于5%)。
(6)群丛Ⅵ。蒲公英+羊茅+车前(Ass.Taraxacummongolicum+Festucaovina+Plantagoasiatica)。群落中蒲公英密度19±7株·m-2,盖度88%±4%;羊茅密度17±21株·m-2,盖度7%±10%;车前密度5±5株·m-2,盖度2%±3%。群落中主要的伴生种有:委陵菜和老鹳草,盖度均小于5%。
3.2 土壤因子与样地的分布
CCA排序图中前两个轴的物种与环境相关系数较高,解释了物种—环境关系总方差的51.9%。保留前两轴可以较好地反映云顶山植被与土壤化学因子之间的关系。
在CCA排序图中,带箭头的连线表示土壤因子,线段的长度表示样地、物种分布和该土壤因子的相关性大小,箭头连线和排序轴的夹角代表该土壤环境因子和排序轴的相关性大小。
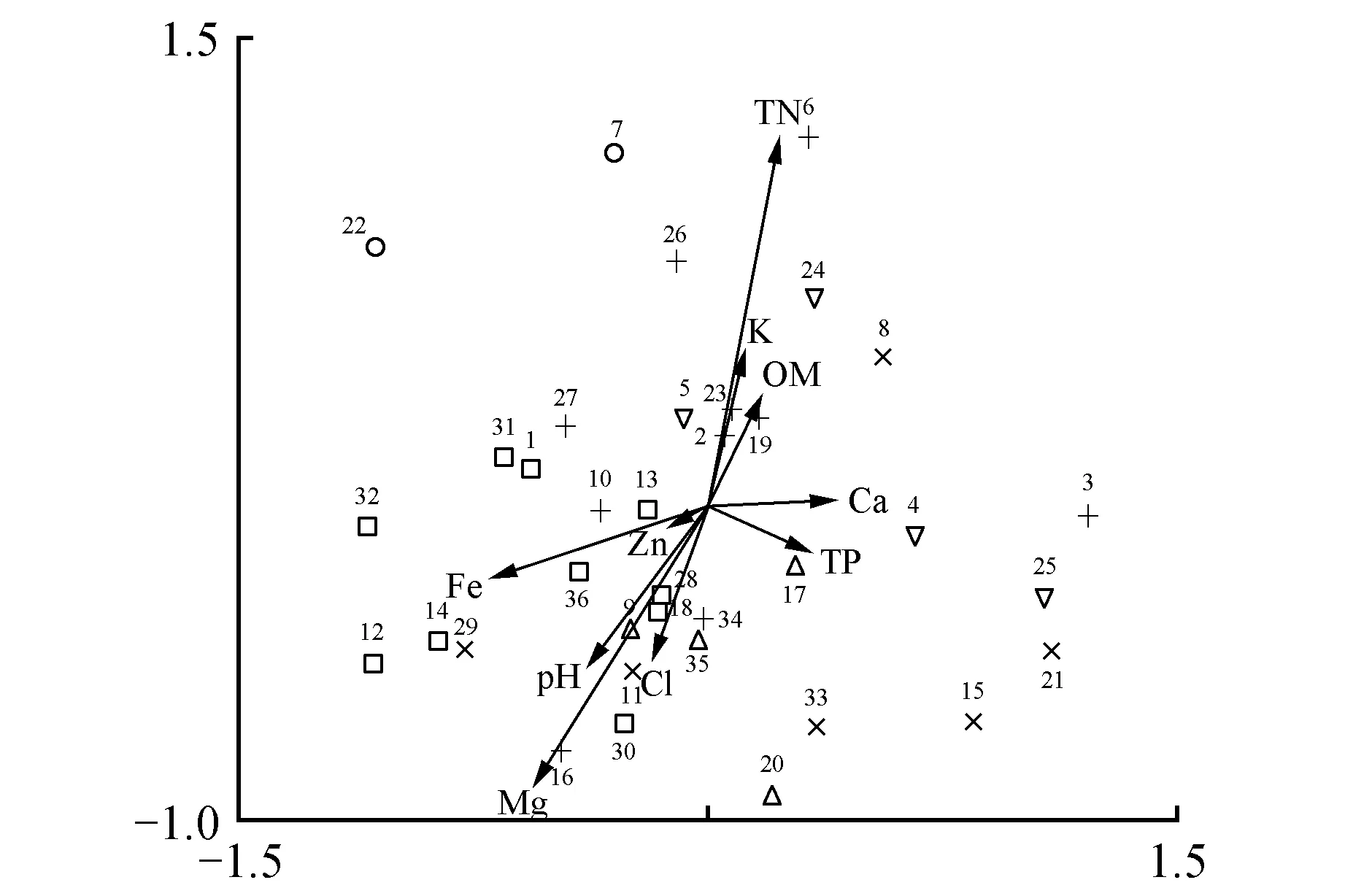
图2 云顶山亚高山草甸群落样地和土壤因子的CCA排序图 △.Ⅰ;▽.Ⅱ;×.Ⅲ;+.Ⅳ;□.Ⅴ;○.ⅥFig.2 CCA bioplot of samples and soil factors of community of subalpine meadow of Yundingshan △.Ⅰ;▽.Ⅱ;×.Ⅲ;+.Ⅳ;□.Ⅴ;○.Ⅵ

表1 不同群丛的土壤化学因子
从表1和图2可以看出不同群丛的化学因子及其在CCA排序图上的分布:
群丛Ⅰ位于第三、四象限中部,其生境中Cl-含量、Mg2+含量较高;群丛Ⅱ位于第一、四象限中部,其生境中TN、TP、OM、Ca2+含量较高;群丛Ⅴ位于第三象限最左端,其生境中Mg2+、Zn2+、Fe的含量较高。
群丛Ⅲ、群丛Ⅳ、群丛Ⅵ分别位于第四象限右端,第一、二象限中部和第二象限左端。
第一排序轴与Ca2+呈正相关,与Fe呈负相关;第二排序轴与TN呈正相关,与Mg2+呈负相关(图2)。
据此可以认为:CCA排序结果较好地反映了云顶山亚高山草甸群落与土壤环境因子的关系。

图3 不同群丛的物种多样性指数Fig.3 Species diversity indexes in different associations
3.3 不同群丛的物种多样性指数
从图3可以看出,不同群丛之间的丰富度指数和Shannon-Weiner指数的变化趋势一致:Ⅱ>Ⅳ>Ⅰ>Ⅲ>Ⅴ>Ⅵ;群丛Ⅰ的丰富度指数与其他群丛的丰富度指数均差异显著(P<0.05),群丛Ⅱ的丰富度指数只与群丛Ⅳ的丰富度指数无显著差异(P>0.05);群丛Ⅰ的Shannon-Weiner指数只与群丛Ⅲ的Shannon-Weiner指数无显著差异(P>0.05);群丛Ⅱ的Shannon-Weiner指数只与群丛Ⅳ的Shannon-Weiner指数无显著差异(P>0.05);不同群丛的均匀度指数之间无显著差异(群丛Ⅳ除外)(P>0.05)。
3.4 土壤因子与物种多样性指数的相关性
Mg2+(P<0.05)、Cl-(P<0.01)含量与丰富度指数呈负相关,TN和K+含量与丰富度指数呈正相关(P<0.05);Cl-含量与Shannon-Weiner指数呈极显著负相关(P<0.001),TN(P<0.05)、OM(P<0.05)、Ca2+(P<0.05)和K+(P<0.01)含量与Shannon-Weiner指数呈正相关;仅K+含量与均匀度指数呈正相关(P<0.05)(表2)。
表2土壤化学因子与物种多样性指数之间的相关性
Table2Correlationsofspeciesdiversityindexesandsoilchemicalfactors
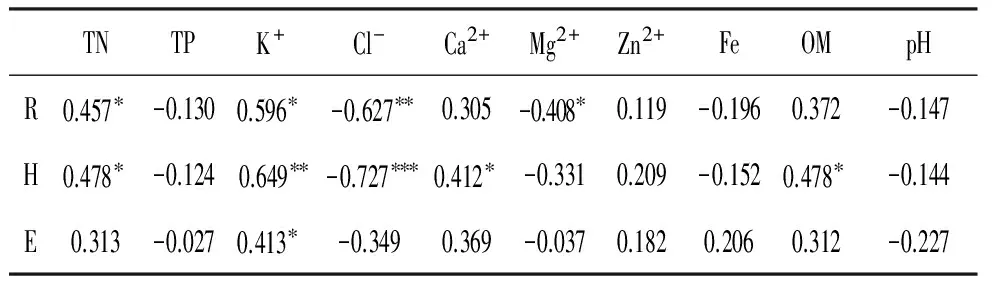
TNTPK+Cl-Ca2+Mg2+Zn2+FeOMpHR0.457∗-0.1300.596∗-0.627∗∗0.305-0.408∗0.119-0.1960.372-0.147H0.478∗-0.1240.649∗∗-0.727∗∗∗0.412∗-0.3310.209-0.1520.478∗-0.144E0.313-0.0270.413∗-0.3490.369-0.0370.1820.2060.312-0.227
注:***P<0.001,**P<0.01,*P<0.05,下同。
Note:***P<0.001,**P<0.01,*P<0.05,the same as below.
3.5 多样性指数间的相关性
Shannon-Weiner指数与丰富度指数(P<0.001)、均匀度指数(P<0.01)均呈正相关;丰富度指数与均匀度指数无显著相关性(P>0.05)(表3)。
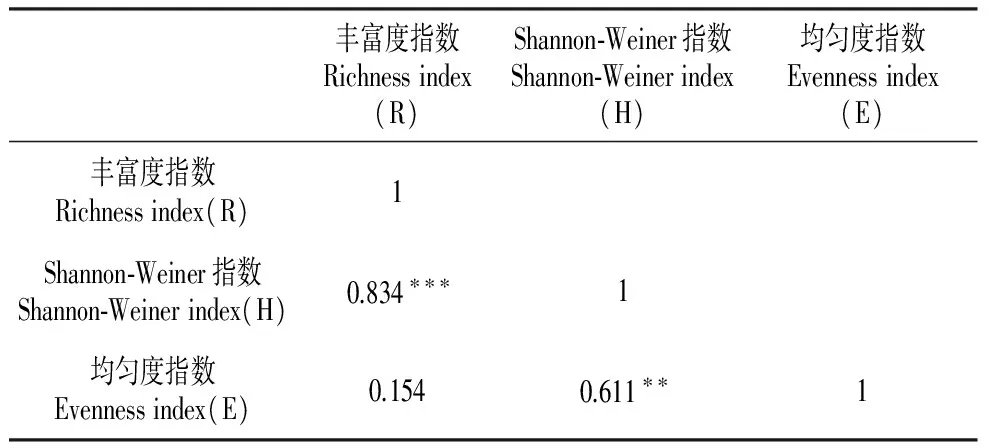
表3 物种多样性指数之间的相关性

表4 土壤因子之间的相关性
3.6 土壤因子间的相关性
TN与TP(P<0.05)、OM(P<0.01)、K+(P<0.01)、Ca2+(P<0.01)、Zn2+(P<0.01)均呈正相关,与Cl-呈负相关(P<0.01);Fe与TP(P<0.01)、OM(P<0.05)、K+(P<0.01)、Ca2+(P<0.01)、Zn2+(P<0.01)、Mg2+(P<0.01)均呈正相关(表4)。
4 讨论
土壤化学因子直接影响植物所需养料物质的来源和吸收,如OM是土壤中养料物质的主要来源,pH影响矿质元素的有效性,因此,土壤化学因子影响植被分布、物种多样性。
相关研究表明土壤化学因子是影响植被分布的关键因素[10,12],本文的研究结果进一步证实了这一点。TN、Ca2+、Mg2+、Fe是影响云顶山亚高山草甸植被分布的主要土壤化学因子。不同的群落类型分布在适宜的土壤化学因子范围内:由蒲公英、钝裂银莲花、车前为主要物种的群丛Ⅰ中Mg2+、Cl-含量较高;由委陵菜、鳞叶龙胆、蒲公英、车前为主要物种的群丛Ⅱ中TN、Ca2+的含量最高,由车前、白莲蒿、委陵菜为主要物种的群丛Ⅴ中Mg2+、Fe含量最高。相关研究表明TN是影响湿地植被分布的关键因子[23],土壤含水量和电导率是影响黄河三角洲海滨草甸的关键因素[24],海拔和水分是影响云顶山植被分布的关键因素[21],盐分、pH是影响盐沼植被分布的主要因素[12]。由此可见,不同地域影响其植被分布的土壤化学因子不同,土壤化学因子会影响植被的分布,反过来,植被的分布也会影响土壤化学因子的含量。
氮和钾均是植物生长发育所需的大量元素。在黄土高原,氮是植物生长的主要限制性因子之一。云顶山亚高山草甸研究中TN和K+的含量均与丰富度指数、Shannon-Weiner指数呈正相关,表明了TN和K+对物种多样性影响较大。相关研究表明TN与多样性指数之间的关系包括三种情形:正相关[25~26]、负相关[11,27]、无相关性[10,12,23~24,28~29];K+与多样性指数之间的关系也包括同样的三种情形[10,23~24,27,29~30]。土壤有机质与多样性之间的关系无固定规律可循。一种看法认为:群落的物种多样性最高时,土壤养分位于“中间位置”[31];另一种看法认为:随着土壤矿质营养的增加,物种丰富度并未降低,在土壤矿质营养水平最高时,物种的丰富度最高[31]。本研究中,丰富度指数和Shannon-Weiner指数在有机质含量最高时最大,与第一种看法一致,并且有机质与Shannon-Weiner指数呈正相关。一方面,物种多样性影响土壤中有机质、氮和钾的含量,云顶山亚高山草甸的草本植物残体进入土壤后,在微生物的作用下容易分解,增加土壤中有机质的含量,另外,TN、K+含量和有机质含量呈正相关,土壤中有机质含量增加的同时氮、钾的含量也增加。Tilman认为:物种多样性越高,土壤中的氮被利用的越充分,同时增加了土壤中矿质营养的量[32~33]。另一方面,有机质、氮和钾的含量也影响着物种多样性。据此可以认为:TN、OM与物种多样性的关系在不同的研究区域、对不同的研究对象是不一致的。
本研究中,Ca2+与Shannon-Weiner指数呈正相关是因为Ca2+在维持细胞内离子的稳态中起重要作用,较高的Ca2+含量可以提高植物对逆境的抗性,有利于群落多样性的提高,并且有研究表明土壤中Ca2+含量升高可以提高物种丰富度指数[34]。云顶山亚高山草甸土壤中Mg2+含量和Cl-含量对植被影响较大:Mg2+与丰富度指数呈负相关,Cl-与丰富度指数、Shannon-Weiner指数呈负相关。Mg2+是植物生长必需的大量元素,是植物生长的限制因素,但贺强等在研究黄河河口盐沼植被分布、多样性与土壤化学因子的相关关系时认为Na+可以促进Mg2+的吸收,提高物种多样性[12]。另有研究表明,土层的深度不同时Mg2+与多样性指数的相关性也不同[10]。Cl-是植物必需的微量元素之一,含量较高对植物有毒害作用,会影响亚高山草甸群落物种多样性。
与云顶山相关的研究表明:土壤含水量、干扰是影响云顶山亚高山草甸植被分布的重要因素[15,18],结合土壤的化学性质,可以说明土壤的理化性质、干扰等因素共同决定了云顶山亚高山草甸植被分布的复杂性。王琳等通过对历山草甸的研究认为,Cu2+与草甸群落物种多样性指数呈正相关[27],而本研究未测定Cu2+的含量,如要更全面的研究云顶山亚高山草甸和土壤因子之间的关系还需测定其他的土壤因子,如:全碳、速效磷、速效钾、速效氮、Cu2+、Mn2+、Na+、土壤孔隙度、土壤含水量、土壤含盐量(电导率)、土壤容重等。
植物群落和土壤是一个有机整体,植物群落是各种土壤因子综合作用的表现形式,同时,植物群落也会影响土壤的理化性质,因此,土壤因子与植被分布、物种多样性相关性的研究结果会随着研究地域、土壤类型、气候条件的不同而不同。
综上,TN、Ca2+、Mg2+、Fe是影响云顶山亚高山草甸植被分布的主要土壤化学因子。Mg2+、Cl-、TN、K+对丰富度指数有显著的影响;Ca2+、Cl-、TN、OM和K+对Shannon-Weiner指数有显著的影响。因此,物种多样性的保护与恢复应结合当地的土壤特点。
1.Goldberg D E,Miller T E.Effects of different resource additions on species diversity in an annual plant community[J].Ecology,1990,71(1):213-225.
2.江洪,黄建辉,陈灵芝,等.东灵山植物群落的排序、数量分类与环境解释[J].植物学报,1994,36(7):539-551.
3.廉凯敏,吴应建,张丽,等.太宽河自然保护区板栗群落数量分类与排序[J].生态学杂志, 2015,34(1):33-39.
4.李伟,张翠萍,魏润鹏.广东中西部桉树人工林植物多样性与林龄和土壤因子的关系[J].生态学报,2014,34(17):4957-4965.
5.王顺忠,陈桂琛,柏玉平,等.青海湖鸟岛地区植物群落物种多样性与土壤环境因子的关系[J].应用生态学报,2005,16(1):186-188.
6.赵学春,来利明,朱林海,等.三工河流域琵琶柴群落特征与土壤因子的相关分析[J].生态学报,2014,34(4):878-889.
7.潘瑞炽.植物生理学:第6版[M].北京:高等教育出版社,2008:28-33.
8.杨允菲,祝廷成.植物生态学:第2版[M].北京:高等教育出版社,2011:277-286.
9.Holmes P M.Shrubland resoration following woody alien invasion and mining:Effects of top soil depth,seed source,and fertilizer addition[J].Restoration Ecology,2001,9(1):71-84.
10.张林静,岳明,顾峰雪,等.新疆阜康绿洲荒漠过渡带植物群落物种多样性与土壤环境因子的耦合关系[J].应用生态学报,2002,13(6):658-662.
11.宋创业,郭柯,刘高焕.浑善达克沙地植物群落物种多样性与土壤因子的关系[J].生态学杂志,2008,27(1):8-13.
12.贺强,崔保山,赵欣胜,等.黄河河口盐沼植被分布、多样性与土壤化学因子的相关关系[J].生态学报,2009,29(2):676-687.
13.上官铁梁,张峰.云顶山虎榛子灌丛群落学特征及生物量[J].山西大学学报,1989,12(4):347-352.
14.上官铁梁,张峰.云顶山植被及其垂直特征研究[J].山地研究,1991,9(1):19-26.
15.李素清,张金屯.山西云顶山亚高山草甸群落生态分析[J].地理研究,2007,26(1):83-90.
16.李素清,杨斌胜,张金屯.山西云顶山亚高山草甸优势种群和群落的格局分析[J].应用与环境生物学报,2007,13(1):9-13.
17.李斌,李素清,张金屯.云顶山亚高山草甸优势种群生态位研究[J].草业学报,2010,19(1):6-13.
18.卢爱英,张先平,王世裕.干扰对云顶山亚高山草甸群落物种多样性的研究[J].植物研究,2011,31(1):73-78.
19.刘莹,张峰,梁小明,等.山西云顶山自然保护区野生种子植物区系研究[J].植物科学学报,2012,30(1):31-39.
20.刘海强,刘莹,张峰,等.山西云顶山自然保护区野生资源植物研究[J].山西大学学报:自然科学版,2013,36(1):133-138.
21.刘明光,刘莹,张峰,等.云顶山自然保护区植物群落的分类与排序[J].林业资源管理,2011,8(4):82-88.
22.鲍士旦.土壤农化分析[M].北京:中国农业出版社,2000:177-201.
23.张江英,周华荣,高梅.艾里克湖湿地植物群落特征指数与土壤因子的关系[J].生态学杂志,2007,26(7):983-988.
24.谭向峰,杜宁,葛秀丽,等.黄河三角洲滨海草甸与土壤因子的关系[J].生态学报,2012, 32(19):5998-6005.
25.白永飞,李凌浩,王其兵,等.锡林河流域草原群落植物多样性和初级生产力沿水热梯度变化的样带研究[J].植物生态学报,2000,24(6):667-673.
26.李新荣,张景光,刘立超,等.我国干旱沙漠地区人工植被与环境演变过程中植物多样性的研究[J].植物生态学报,2000,24(3):257-261.
27.王琳,张金屯,上官铁梁,等.历山亚高山草甸的物种多样性及其与土壤理化性质的关系[J].应用与环境生物学报,2004,10(1):18-22.
28.崔宁洁,张丹桔,刘洋,等.不同林龄马尾松人工林林下植物多样性与土壤理化性质[J].生态学杂志,2014,33(10):2610-2617.
29.王慧敏,张峰,庞春花,等.汾河流域中下游植物群落物种多样性与土壤因子的关系[J].西北植物学报,2013,33(10):2077-2085.
30.杨小波,张桃林,吴庆书.海南琼北地区不同植被类型物种多样性与土壤肥力的关系[J]. 生态学报,2002,22(2):190-196.
31.Gentry A H.Changes in plant community diversity and floristic composition on environmental and geographical gradients[J].Ann Missouri Bot Gard,1988,75(1):1-34.
32.Tilmam D,Haddi A.Drought and biodiversity in grasslands[J].Oecology,1992,89(2):257-264.
33.Tilmam D.Causes,consequences and ethics of biodiversity[J].Nature,2000,405:208-211.
34.Roem W J,Klees H,Berendse F.Effects of nutrient addition and acidification on plant species diversity and seed germination in heathland[J].Journal of Applied Ecology,2002,39(6):937-948.
投稿须知
1.投稿前请登陆本刊网站(http://bbr.nefu.edu.cn/)认真阅读本刊投稿中心中的各项说明,并按照本刊要求修改和补充论文中的有关内容,使论文符合本刊要求。投稿时请附作者单位介绍信或全体作者签名,说明无泄密、署名无误和未一稿多投,网上投稿请签订版权转让协议。作者自留底稿及软盘备份。
2.投稿时,请登陆本刊网站在线提交论文。投稿后,可随时登陆查询稿件处理状况。
3.稿件如初审通过,请作者按要求邮寄已签名的《版权转让协议》。
4.稿件经过外审和编审后,如符合我刊要求,须交寄发表费,待发表费收到后,作者可登陆本刊网站后,在发稿状态下打印《稿件录用通知》。
5.作者文责自负。本编辑部对来稿可做必要修改。
6.稿件一经发表,酌致稿酬,并赠样刊2本。
7.凡在本刊发表的论文,如荣获省部级以上成果奖,请及时通知本编辑部,并提供获奖证书复印件。
《植物研究》编辑部
RelationshipsbetweenVegetationDistribution,SpeciesDiversityofSubalpineMeadowandSoilChemicalFactorsintheYundingshan,China
DU Jing-Qi1ZHANG Qiao-Xian1*TIAN Xiao-Dong1WANG Yue1XU Xin-Yun1SONG Li1YUAN Zi-Qiang2
(1. Institute of Plant Chemistry,Lüliang University,Lüliang 033000;2.State Key Laboratory of Grassland Agro-Ecosystems,Lanzhou 730000)
With the data from field investigation of 36 sampling areas with 27 species of the community of subalpine meadow in the Yundingshan, China, we used Two-way Indicator Species Analysis(TWINSPAN) to compartmentalize the sampling areas into different associations. Six associations were obtained: Ass.Taraxacummongolicum+Anemoneobtusiloba+Plantagoasiatica, Ass.Potentillachinensis+Gentianasquarrosa+Taraxacummongolicum+Plantagoasiatica, Ass.Taraxacummongolicum+Potentillachinensis+Thymusmongolicus, Ass.Plantagoasiatica+Potentillachinensis+Thymusmongolicus+Artemisiasacrorum, Ass.Plantagoasiatica+Artemisiasacrorum+Potentillachinensis, and Ass.Taraxacummongolicum+Festucaovina+Plantagoasiatica. By Canonical Correspondence Analysis(CCA), we analyzed the relationships between the meadow vegetation distribution patterns and soil chemical factors. Soil chemical factors consisted of soil pH, total nitrogen, total phosphprus, K+, Cl-, Ca2+, Mg2+, Zn2+, Fe, organic matter(OM). The total nitrogen, Ca2+, Mg2+, Fe were the main soil environmental factors which affected distribution of the community of subalpine meadow in the Yundingshan. Six associations were, respectively, correlated with different habitat. Correlation was carried out between species richness index, evenness index and Shannon-Weiner index, the results showed that total nitrogen, Ca2+showed a significant positive correlation with Shannon-Weiner index(P<0.05), a positive relationship was between total nitrogen and species richness index(P<0.05), a negative relationship was between species richness index and Mg2+(P<0.05), a positive relationship of between species richness index(P<0.05), evenness index(P<0.05), Shannon-Weiner index(P<0.01) and K+. Therefore, the protecting species diversity should be combined with local characteristics of the soil.
species diversity;distribution;Two-way Indicator Species Analysis(TWINSPAN);Canonical Correspondence Analysis(CCA);soil chemical factors
山西省基础研究自然科学基金项目(2013011029-1);山西省高等学校科技创新项目(2013167);吕梁学院校内基金(ZRXN201309)
杜京旗(1984—),男,讲师,硕士,主要从事群落生态学、农业生态学研究。
2015-11-29
Q948
A
10.7525/j.issn.1673-5102.2016.03.019
猜你喜欢
成都信息工程大学学报(2022年4期)2022-11-18
青海草业(2022年2期)2022-07-23
昆明医科大学学报(2022年2期)2022-03-29
食品安全导刊(2021年20期)2021-08-30
黑龙江工程学院学报(2020年5期)2020-10-21
河南科学(2020年3期)2020-06-02
水土保持研究(2019年6期)2019-10-19
活力(2019年21期)2019-04-01
河北林业科技(2016年5期)2016-11-08
草业科学(2015年9期)2015-12-11
