
白菜型油菜亚种间杂种的合成
2016-10-31 09:01陈纪鹏于恩厂廖婷婷朱一鸣
浙江农业学报 2016年1期
陈纪鹏,于恩厂,廖婷婷,朱一鸣
(1. 宜春学院 生命科学与资源环境学院/江西省植物生长发育调控重点实验室,江西 宜春 336000; 2. 驻马店农业学校,河南 驻马店 463000)
白菜型油菜亚种间杂种的合成
陈纪鹏,于恩厂,廖婷婷,朱一鸣
(1. 宜春学院 生命科学与资源环境学院/江西省植物生长发育调控重点实验室,江西 宜春 336000; 2. 驻马店农业学校,河南 驻马店 463000)
采用蕾期人工去雄与辅助授粉的方式,以白菜型油菜十月红菜薹和黄芽白进行正反交,以较高的频率获得了杂种后代。杂种后代叶型、株型和花等形态均表现为双亲的中间型。叶片颜色比黄芽白更深,均偏向十月红菜薹的紫红色;植株表现出较强的杂种优势,株高大于双亲,产生大量的分枝,角果数远多于双亲。杂种后代花器官发育正常,可育花粉为99%以上,且自交结实率较高。花粉母细胞减数分裂比较正常,95%的花粉母细胞在终变期所有染色体进行同源配对,形成10个二价体,减数第二次分裂中期有部分出现染色体落后现象。杂种后代极强的杂种优势和较好的自交结实性为进一步选育具有优良特性的新型白菜型油菜提供了可能。
红菜薹;黄芽白;亚种间杂交;胚培养
通过人工杂交与辅助授粉的方式对作物近缘种进行杂交,可以获得具有优良特性的相应杂交新品种,进而改良原有作物的不良特性。人们从19世纪开始就对芸薹属种间的杂交及其与异属间的杂交,获得亲缘关系较近的种间杂交种[1]、属间杂交种[2]及亲缘关系甚远的族间杂交种[3]。通过把近缘种的优良基因转入到芸薹属栽培植物中,可以对栽培种进行改良,从而创造出新的作物品种;也可以运用杂交的方式对植物进行改造来获取新的植物类型[4]。虽然现代生物技术发展较快,可以将外源基因通过转基因方式直接导入农作物中对作物进行遗传改良。但由于当前克隆技术还不够成熟,控制数量性状的基因获取难度较大。所以,种间和种内杂交依然是利用近缘物种的优良基因来改良作物品种遗传性状的有效途径[5-6]。伴随着现代生物技术的迅速发展,植物组织培养技术中的幼胚培养、子房培养及体细胞融合等方法在远缘杂交中的应用日益广泛,通过这些方法获得了相应的杂种后代,这解决了杂交过程中存在的一些生殖隔离障碍[7-8]。当前,杂交在芸薹属油料作物育种中的应用相当广泛,如通过白菜型油菜与甘蓝型油菜进行杂交,解决了甘蓝型油菜的晚熟性问题;通过与黄籽油菜之间的杂交已获得了具有黄色种皮的杂种后代,进而选育出优良性状的甘蓝型油菜后代[9-10]。
白菜型油菜(Brassicacampestris, AA, 2n=20)是十字花科芸薹属小白菜的一个变种,它是芸薹属中重要的油料栽培作物,并且分布较为广泛。其中,白菜型油菜的优良性状有:分枝性强、耐贫瘠、抗寒性强且生育期短等[11]。红菜薹(B.campestrisL. ssp. Chinenses var. purpurea Bialey, 2n=20)是十字花科芸薹属芸薹种的菜薹变种,又称紫菜薹、红油菜薹,是原产于中国的富有营养价值的蔬菜,备受人们喜爱,其中主要以武汉、长沙、成都3个地区种植最早[12]。十月红是由红菜薹品种胭脂红通过天然杂交产生的后代中选育出来的具有稳定遗传性状的晚熟品种。
目前,国内很多学者对红菜薹的研究集中在传统的品种选育[13]与雄性不育的利用[14-18]等方面,也进行了甘蓝型油菜与红菜薹的杂种后代的遗传分析[19]。因为红菜薹属于异化授粉植物,花器官较小,不便于单花授粉结实,所以表现出较强的杂种优势。伴随杂交育种工作大量进行,红菜薹杂交种的推广较为广泛。所以一些优良的品种种质资源也将面临丢失或已经丢失,迫切需要对其种质资源进行收集、保护及研究。基于南方白菜型油菜大多是自交不亲和性的类型,本项试验通过人工去雄、辅助授粉的方法将红菜薹与黄芽白进行正反交,研究杂交亲和性,并对其杂种后代进行遗传分析,一方面可以解决白菜型油菜自交不亲和或亲和力低的问题,另一方面扩充了红菜薹的遗传基础,进而为选育出其他新品种提供材料。
1 材料与方法
1.1 材料
供试材料白菜型油菜十月红菜薹与黄芽白作亲本进行正反交。十月红菜薹是在天然杂交后代中选育出的常规品种,长势好,茎叶呈紫红色,可食用部分味微甜,具有较强的耐旱和耐渍性,抗病毒病和霜霉病。黄芽白(B.campestrisssp. pekinensis)是十字花科白菜型油菜的白菜亚种,在全国范围内广泛种植。为了满足上述材料的生长发育特性,选择在10月上旬的适宜天气分别播种于田间。第二年春季进行杂交,进行胚培养或直接收取成熟种子。将杂交种子种植用于杂种形态学观察和花粉育性观察,并进行花粉母细胞减数分裂观察。
1.2 方法
1.2.1 亚种间杂交在杂交前1 d对父本进行套袋处理。在母本盛花期时选择晴天上午,用经酒精处理过的镊子把母本的雄蕊剥除干净彻底,并套上纸袋。次日上午,在已套袋的父本上选择刚刚开放的新鲜花粉均匀地涂于相应的杂交母本柱头上。授粉完全后,再将纸袋套上防止相互串粉。授粉7 d左右,将纸袋摘掉,以便植株更好地生长。授粉15~20 d 时取部分膨大的角果进行胚培养,剩余角果成熟后收获种子。每10朵花为一个重复,重复3次。计数每一杂交组合所获得的结角数、饱满种子数及出芽率,并采用Excel 2007进行单因素方差分析比较正反交之间的差异。
1.2.2 胚培养授粉15~20 d时,从母本植株上取下人工授粉后膨大的角果。将取下的角果放在装有75%酒精的烧杯中处理2~3 min后取出,再放入0.1%升汞溶液中处理15 min,对角果进行表面消毒。将角果从升汞中取出用蒸馏水漂洗3遍,洗净残留在角果表面的有毒物质。在无菌操作台上用已消好毒的镊子、刀片轻轻地剥开角果,再将幼胚从胚珠中取出并接种于MS培养基表面,置于25 ℃培养箱培养。
待幼胚长出芽后,将幼芽转接在MS+0.25 mg·L-1NAA+1.5 mg·L-16-BA培养基上进行快速繁殖。得到足够的新幼芽后转入不加激素的MS培养基中进行生根培养;待幼芽生出较多根时即可将组培苗从培养基中取出移栽至田间。
1.2.3 花粉育性观察选择上午气温较高的晴好天气、花粉活力最强的时候分别在每个待检测的植株上各取5朵花,取下其花药并用镊子将花药挤破使花粉散发出来,并把花药壁残片去除。用胶头滴管滴加少量1%醋酸洋红染色液,并把盖玻片轻轻盖上。显微镜10倍物镜下观察其花粉形态和颜色,其中,可育花粉的形状是大而圆且被染成红色,不育花粉的形状是小而瘪着色浅或不着色。
1.2.4 细胞遗传学观察有丝分裂观察:于杂种植株现蕾初期取下其幼嫩花蕾,挑取子房,用0.1% 8-羟基喹啉溶液处理3~4 h,使其有丝分裂停留于中期。然后,将其放入卡诺固定液(无水乙醇和冰乙酸体积比为3∶1)中固定24 h以上,固定完全后转入75%酒精中,并贮藏在-20 ℃冰箱中备用。从冰箱中挑取已固定好的子房,先置于1 mol·L-1盐酸中,60 ℃水浴加热后,再用卡宝品红进行染色压片,最后显微镜下观察,并记录其体细胞染色体的数目。
减数分裂观察:先用卡诺固定液对盛花期取下的小花蕾进行固定,方法与子房固定相同。固定后花蕾褪色直至为白色,置于4 ℃冰箱中贮存。从冰箱中挑取已固定好的花药放至浓度为1 mol·L-1的盐酸中,进行60 ℃水浴加热3 min后,取出并放入清水中,接着用卡宝品红进行染色压片,将压片置于显微镜下观察其花粉母细胞染色体行为。
2 结果与分析
2.1 杂交亲和性观察
红菜薹与黄芽白进行正、反交,采用胚培养与直接收获种子两种方式均获得杂种后代。授粉后每朵花都能形成角果,以红菜薹为母本的正交组合每个角果形成饱满种子数较多,平均每个角果形成6.7粒饱满种子。仅有少数种子发育不完全,形成不饱满种子。而反交组合每个角果平均形成3.8粒种子,也形成少量不饱满种子。正交组合不但形成种子数量较多,而且饱满种子发芽率也高于反交组合。而正、反交组合所形成的不饱满种子均不能发芽。正交组合更容易获得杂种后代,其亲和指数显著高于反交组合(F=45,P<0.05,表1)。
表1 黄芽白与十月红菜薹杂交亲和性
Table 1 Hybridization compatibility ofB.campestrisssp. pekinensis andB.campestrisL. var. purpurea
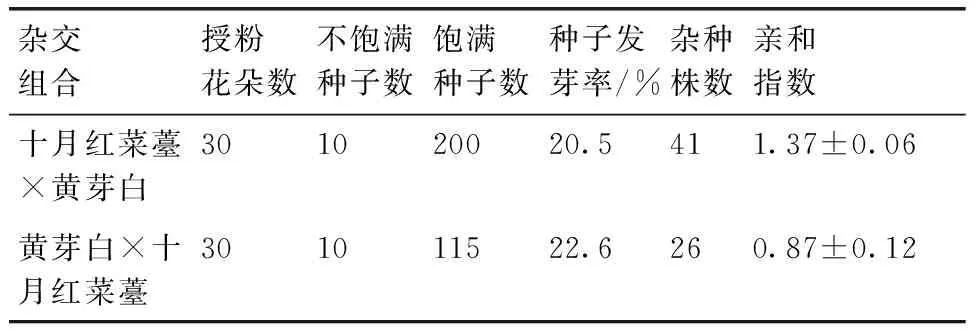
杂交组合授粉花朵数不饱满种子数饱满种子数种子发芽率/%杂种株数亲和指数十月红菜薹×黄芽白301020020.5411.37±0.06黄芽白×十月红菜薹301011522.6260.87±0.12
注:亲和指数=获得杂种植株数/授粉花朵数。
2.2 胚培养结果
在授粉后20 d后对正交组合进行胚培养,培养5 d左右幼胚开始产生大量白色根毛,10 d后开始生芽。80%的胚发育成苗,其余的胚发育成畸形胚不能成苗,只有极少数的幼胚死亡。与直接收种子相比,胚培养可提高杂种的成苗率。不论是直接收种子还是胚培养所获得的植株,经形态学和细胞学鉴定均为真杂种。
2.3 杂种的形态学及花粉育性特征
正、反交杂种后代形态极相似,幼苗期且植株叶柄及叶脉颜色与黄芽白相比较深,都偏向于十月红菜薹的紫红色(表2,图1-A,B,C,D)。植株叶色较深,叶缘呈波状且有不规则的叶裂(图1-E)。成熟期植株表现出较强的杂种优势,植株高度均高于双亲。而且植株形成更多分枝,且分枝粗壮,基部着生。植株下部茎秆也呈现十月红菜薹紫红色。上部茎秆的紫色虽不是很明显,但整体上颜色还是较黄芽白深。正反交组合杂种植株花器官都发育正常,花丝较长,花药饱满,伸到柱头顶部。晴好天气产生大量花粉,可育花粉均达到99%以上(图1-F)。正反交杂种植株自交结实性较好,几乎每朵花授粉后都能形成角果,每个角果都能形成较多种子。反交组合杂种在成熟期生长更加旺盛,植株分枝更多,形成更多角果。
2.4 正反交杂种的细胞学观察
形态学鉴定结果显示所得的杂交植株是白菜型油菜与十月红菜薹的真杂种。杂种植株体细胞染色体数目为20,未发现非整倍体现象。在花粉母细胞减数分裂前期,所有染色体正常联会配对,形成10个二价体,无单价体出现(图1-G)。在减速第Ⅱ次中期染色体整齐地排列在赤道板上,但有少部分细胞出现滞后染色体(图1-H,I),后期Ⅱ可观察到染色体均正常地平均分配到4个细胞中去。整个减数分裂过程表现比较正常,因此所形成的花粉育性很好。
表2 黄芽白与十月红菜薹杂种的形态学特征
Table 2 Morphology of hybrids betweenB.campestrisssp. pekinensis andB.campestrisL. var. purpurea
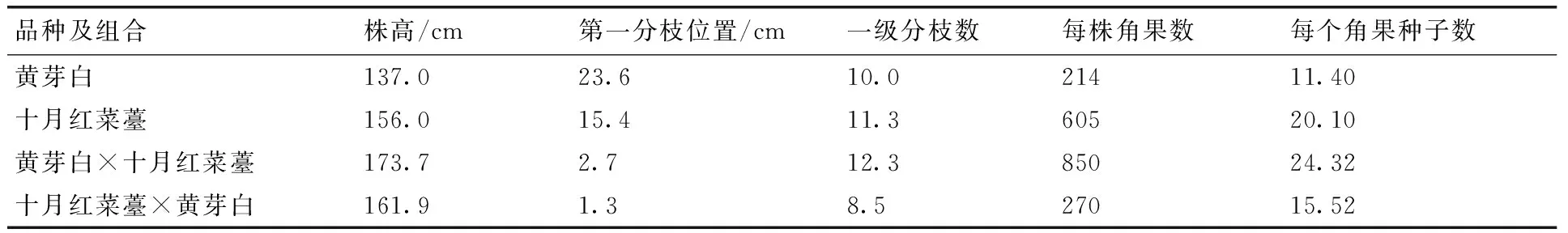
品种及组合株高/cm第一分枝位置/cm一级分枝数每株角果数每个角果种子数黄芽白137.023.610.021411.40十月红菜薹156.015.411.360520.10黄芽白×十月红菜薹173.72.712.385024.32十月红菜薹×黄芽白161.91.38.527015.52

A: 黄芽白; B: 黄芽白×十月红菜薹; C: 十月红菜薹×黄芽白; D: 十月红菜薹; E: 叶片(从左至右依次是黄芽白,十月红菜薹×黄芽白,十月红菜薹)标尺=10 cm; F: 花粉,标尺=100 μm; G: 终变期花粉母细胞; H: 后期Ⅰ花粉母细胞; Ⅰ: 带有落后染色体(箭头所示)的中期Ⅱ花粉母细胞。 标尺=30 μm。图1 黄芽白与十月红菜薹杂种形态学及细胞遗传学Fig.1 Morphology and cytogenetics of hybrids between B.campestris ssp. pekinensis and B.campestris L. var. purpurea
3 结论与讨论
利用白菜型油菜十月红菜薹与黄芽白分别进行正、反交试验,采用胚培养与直接收获种子两种方式均获相应的杂交种子。从杂交结实的情况来看,以十月红菜薹为母本的正交组合获得杂交后代饱满种子数较多,相对较容易些;而以黄芽白为母本的反交组合获得杂交后代饱满种子数较少,相对较难些。在芸薹属其他杂交中也发现有类似的现象[20-22],发现在远缘杂交过程中具有单向不亲和性的特点,同一杂交组合染色体数目不同的亲本进行正反交,一般选用数目多的为母本[23]。本试验选择黄芽白和十月红菜薹两个亲本虽然染色体数相同,但正反交难易程度却有所不同。
通过对本次试验正反交获得的杂交后代群进行花粉可染性、形态学与细胞学观察。正反交杂种(AA,2n=20)在形态上很相似,生长旺盛,表现出极强的杂种优势。当前春油菜区种植的白菜型油菜多是普通品种,所以利用白菜型油菜的杂种优势来选育出新的油菜品种对改善一些高原地区现有的油菜种植现状具有实际意义。Meng等[24]研究显示,获得白菜型油菜A基因的新型甘蓝型油菜比亲本甘蓝型油菜更表现出亚基因间杂种优势。本试验通过对十月红菜薹与黄芽白进行正、反交获得的新油菜品种表现出较强的亚基因间杂种优势。这不仅可以改良现有品种的不足,而且对后期油菜的选育工作提供了新的种质资源。
随着育种实践工作的深入发展,现代生物技术的发展也趋于成熟,可以采用不同方法来克服芸薹属远缘杂交过程中存在的杂交不亲和现状。利用体细胞融合的方法避免了有性杂交的障碍,同时扩大了杂交亲本的选取范围。胚培养、胚抢救、子房培养[25]等组织培养技术不仅克服了芸薹属远缘杂交的不亲和,将不同种属间的优良基因进行转移,从而获得优良品种,而且还可减少杂种后代稳定的育种年限。本项研究通过杂交和收获种子的方法获得了杂种后代,可见,十月红菜薹与黄芽白之间杂交亲和性较好,可以不借助胚培养方法较容易地获得杂种后代。
[1] HERBERT W. On hybridization amongst vegetables[M]. London: Horticultural Society of London, 1847.
[2] MOHANTY A , CHRUNGU B , VERMA N , et al. Broadening the genetic base of crop brassicas by production of new intergeneric hybrid[J].CzechJournalofGeneticsandPlantBreeding, 2009, 45: 117-122.
[3] WEI W H, LI Y C, WANG L J, et al. Development of a novelSinapisarvensisdisomic addition line inBrassicanapuscontaining the restorer gene for Nsa CMS and improved resistance toSclerotiniasclerotiorumand pod shattering[J].TheoreticalandAppliedGenetics, 2010, 120(6): 1089-1097.
[4] PRAKASH S, BHAT S R, QUIROS C F, et al.Brassicaand its close allies: cytogenetics and evolution[J].PlantBreedingReviews, 2009, 31: 21-187.
[5] GARG H, BANGA S, BANSAL P, et al. HybridizingBrassicarapawith wild crucifersDiplotaxiserucoidesandBrassicamaurorum[J].Euphytica, 2007,156(3): 417-424.
[6] WEI W H, LI Y C, WANG L J, et al. Development of a novelSinapisarvensisdisomic addition line inBrassicanapuscontaining the restorer gene for Nsa CMS and improved resistance toSclerotiniasclerotiorumand pod shattering[J].TheoreticalandAppliedGenetics, 2010, 120(6): 1089-1097.
[7] MEI J, LI Q, YANG X, et al. Genomic relationship between wild and cultivatedBrassicaoleraceaL. with emphasis on the origination of cultivated crops[J].GeneticResourcesandCropEvolution, 2010, 57(5): 687-692.
[8] TU Y, SUN J, GE X, et al. Chromosome elimination, addition and introgression in intertribal partial hybrids betweenBrassicarapaandIsatisindigotica[J].AnnalsofBotany, 2009, 103(7): 1039-1048.
[9] 刘忠松,官春云,李栒.甘蓝型油菜与芥菜型油菜种间杂交研究[J].中国油料作物学报,2001,23(2):82-86.
[10] 文静.芸薹属种间杂交合成甘蓝型黄籽油菜及杂交后代的研究[D].武汉:华中农业大学,2008.
[11] 张万仓,范惠玲,叶剑,等.白菜型油菜自交亲和性变异分析[J].西北植物学报,2006,26(4):688-695.
[12] 吴朝林,陈文超.中国紫菜薹地方品种初步研究[J].中国种业,7(3):7-9.
[13] 吴朝林.紫菜薹新品种五彩薹二号的选育[J].长江蔬菜,2006,20(6):48-49.
[14] 向长萍,晏儒来,李锡香,等.紫菜薹雄性不育系的选育和应用[J].中国蔬菜,2000,21(5):28-30.
[15] 宋秋,许明,司龙亭,等. 紫菜薹细胞核雄性不育系数量性状配合力分析[J].中国农学通报,2006,22(1):343-345.
[16] 黄邦全,李薇,居超民,等.Ogura 雄性不育细胞质导入紫菜薹及杂种优势利用初报[J].种子,1999 (3):57.
[17] 许明,白明义,魏毓棠.不同发育期紫菜薹细胞质雄性不育系及保持系植株体内物质代谢的差异[J].沈阳农业大学学报,2008,39(2):152-155.
[18] 王学芳,李殿荣,张彦锋,等.紫菜薹胞质雄性不育系2A及其杂交种的选育[J].西北农业学报,2008,17(3):250-253.
[19] 李亭亭,徐跃进,万正杰,等.甘蓝型油菜与红菜薹的杂种及其后代遗传变异分析[J].植物科学学报,2011,29(2):189-193.
[20] 周清元,李加纳,殷家明,等.白菜型油菜与羽衣甘蓝种间杂交的初步研究取材时间对子房离体培养结籽率的影响[J].西北农业大学学报,2003,25(6):495-497.
[21] LEFION M, EBER F, LETANNEUR J C, et al. Pairing and recombination at meiosis ofBrassicarapa(AA)×B.napus(AACC) hybrids[J].TheoreticalandAppliedGenetics, 2006, 113(8): 1467-1480.
[22] 邓婧,李栒.芸薹属植物远缘杂交不亲和性以及杂种后代遗传特性的研究进展[J].作物研究,2006 (5): 459-464.
[23] 刘宏波,刘忠松.油菜远缘杂交亲和性研究进展[J].作物研究,2006,6(5):456-458.
[24] MENG J, LI M, CHEN X, et al. Intersubgenomic heterosis in rapeseed production with a partial new-typedBrassicanapuscontaining subgenome ar fromB.rapaand Cc fromB.carinata[J].CropScience, 2006, 46(1): 234-242.
[25] SAMPSON D R. Intergeric pollen stigma incompatibility in the Cruciferae[J].CanadianJournalofGeneticsandCytology, 1962,4(1): 38-49.
(责任编辑 张 韵)
Production of intersubspecific hybrids of Brassica campestris
CHEN Ji-peng1, YU En-chang2, LIAO Ting-ting1, ZHU Yi-ming1(1.CollegeofLifeScienceandEnvironmentandResource/JiangxiProvinceKeyLaboratoryofPlantGrowthandRegulation,YichunUniversity,Yichun336000,China; 2.AgriculturalSchoolofZhumadian,Zhumadian463000,China)
By artificial pollination and supplementary pollination in bud stage, hybrids were obtained through reciprocal cross ofBrassicacampestrisL. ssp. Chinenses var. purpurea Bialey andB.campestrisssp. pekinensis in high frequency. Leaf, plant type and flower morphology of hybrids were characterized by intermediate type of their parents. The leaf color of hybrid was darker thanB.campestrisssp. pekinensis, more closer to purple color ofBrassicacampestrisL. ssp. Chinenses var. purpurea Bialey. The hybrids showed high heterosis for higher plant height, more branches and more pods than the parents, and they had a good seed setting via cross by themselves because of mature flowers which could produce more than 99% fertile pollens. It also indicated that the female gemmates were fertile in the hybrids. All of the chromosomes paired to 10 bivalents at diakinesis in 95% pollen mother cells. A few of lagged chromosomes were observed at metaphase Ⅱ. The hybrids with heterosis and good seed setting was useful for breeding new type ofBrassicacampestris.
B.campestrisL. var. purpurea;B.campestrisvar. pekinensis; hybridization; embryo culture
http://www.zjnyxb.cn
10.3969/j.issn.1004-1524.2016.01.02
2015-06-24
国家自然科学基金项目(31460357);江西省科技支撑项目(20141BBF60010);江西省自然科学基金(20151BAB204026)
陈纪鹏(1974—),男,河南西平人,博士,副教授,主要从事油菜远缘杂交和细胞遗传学研究。E-mail:chensi20020606@163.com
S565.4
A
1004-1524(2016)01-0009-06
浙江农业学报ActaAgriculturaeZhejiangensis, 2016,28(1):9-14
陈纪鹏,于恩厂,廖婷婷,等. 白菜型油菜亚种间杂种的合成[J].浙江农业学报,2016,28(1):9-14.
猜你喜欢
资源导刊(2022年12期)2023-01-03
中国饲料(2022年5期)2022-04-26
南方农业(2021年28期)2021-12-25
杂草学报(2021年2期)2021-10-27
西北园艺(综合)(2021年4期)2021-07-22
食品与健康(2020年6期)2020-06-19
农业科技与装备(2017年9期)2018-03-02
饮食科学(2017年4期)2017-05-03
长江蔬菜(2016年10期)2016-12-01
湖南师范大学自然科学学报(2015年2期)2015-02-27
