
丙肝感染宿主亲嗜性基因OCLN和CD81在猕猴川西亚种组织中的分布
2016-10-31 08:52陈正礼罗启慧史良琴程安春
浙江农业学报 2016年1期
何 柳,陈正礼,罗启慧,邓 娟,夏 玉,史良琴,程安春
(1. 四川农业大学 动物医学院 实验动物疾病模型研究室,四川 温江 611130; 2. 四川农业大学 动物医学院/动物疾病与人类健康中心四川省重点实验室, 四川 雅安 625014)
丙肝感染宿主亲嗜性基因OCLN和CD81在猕猴川西亚种组织中的分布
何 柳,陈正礼,罗启慧,邓 娟,夏 玉,史良琴,程安春
(1. 四川农业大学 动物医学院 实验动物疾病模型研究室,四川 温江 611130; 2. 四川农业大学 动物医学院/动物疾病与人类健康中心四川省重点实验室, 四川 雅安 625014)
丙型肝炎动物模型缺乏限制了丙型肝炎的研究,如果与人类基因背景相似的猕猴能作为丙型肝炎动物模型将有重大意义。文章拟通过检测决定HCV感染物种特异性的关键基因OCLN与CD81在猕猴不同组织的表达量,并以HCV易感的人肝癌细胞系Huh 7.5.1为对照,来探究猕猴作为丙型肝炎动物模型的可能性。结果表明,OCLN与CD81在猕猴川西亚种的肝脏、脾脏、肺脏、淋巴结和脊髓中均有表达,但表达量均极显著低于在Huh 7.5.1细胞系中的表达量(P<0.01)。肝脏中的OCLN表达量略低于肺脏中的表达量,而肝脏CD81的表达量均高于其他组织。并且,OCLN在不同组织中的表达水平与人体OCLN表达规律相符合(脾<肝<肺)。该结果提示HCV感染的宿主亲嗜性基因OCLN与CD81在猕猴中的低表达量可能导致HCV不能有效地结合组织靶细胞,因此推测,猕猴不能直接用于丙型肝炎动物造模可能与此有关。今后可通过转基因等技术增强OCLN与CD81基因的表达,从而使猕猴获得直接感染HCV的能力。
HCV;OCLN;CD81;猕猴川西亚种;动物模型
咬合蛋白Occludin(OCLN)是构成紧密连接(tight junctions,TJ)的组成成分之一,其进化保守,小鼠、人类和犬的OCLN基因有90%的同源性[1]。在正常发育的肺、肝等组织中OCLN具有高表达水平[2]。研究表明,紧密连接蛋白OCLN是HCV进入细胞的关键因素之一[3]。Cluster designation 81(CD81)是4次跨膜蛋白超家族成员之一,与细胞的吸附、激活、增殖与分化等有关[4]。研究发现,CD81是HCV入胞的受体,其通过与HCV E2结合,使HCV吸附于细胞表面。Ploss等[5]将人CD81和OCLN以及鼠的CLDN1和SR-BI转染到非肝来源的细胞中,发现这些原本无法被HCV感染的细胞获得了HCV易感性,进而将可以感染HCV的Huh 7.5与HepB细胞中的OCLN,CD81分子表达沉默,发现两种细胞感染HCVpp和HCVcc的能力均有下降。该结论证明,OCLN与CD81是介导HCV感染细胞的关键因素之一,并且可能与HCV感染的物种特异性有关[6-7]。
现有研究发现,丙型肝炎病毒(hepatitis C virus,HCV)只能感染人类与黑猩猩,丙型肝炎动物模型的缺乏限制了丙型肝炎的研究。猕猴川西亚种作为中国特有的物种,分布广泛,其亲缘关系与人类非常相近,是良好的人类疾病模型动物。然而Bukh[8]研究发现,猕猴不能感染丙型肝炎。由于病毒特异性受体的表达和分布是影响HCV感染的宿主亲嗜性的重要限制因素,并且其表达量低会限制HCV的感染,因此研究猕猴不同组织的OCLN与CD81表达量对分析猕猴作为丙型肝炎模型的潜力有重大意义。本试验通过对猕猴川西亚种的不同组织中OCLN与CD81的表达量进行检测,并将其与人类肝癌细胞系Huh 7.5.1的OCLN与CD81的表达量相比较,探究猕猴川西亚种作为丙型肝炎动物模型的可能性。丁红方等[9]发现,丙肝患者除了在肝脏组织中检测到HCV阳性以外,在肺,脾,淋巴结等肝外组织中均可以检测到HCV抗原阳性。因此,我们选取了猕猴的肝、脾、肺、淋巴结和脊髓作为材料,通过荧光定量PCR检测OCLN和CD81在这些潜在HCV易感组织中的表达量。
1 材料与方法
1.1 材料
1.1.1 试验动物本试验所用5只健康雄性成年猕猴川西亚种均由四川农业大学实验动物工程技术中心/国家实验用猕猴种源基地(四川雅安)提供[SCXK(川):2013-105]。所有实验动物均由四川省雅安市雅安普莱美生物科技有限公司伦理委员会审查批准使用。实验操作于四川农业大学实验动物工程技术中心[SYXK(川)2014-187]进行。Huh 7.5.1细胞系由武汉病毒研究所提供。
1.1.2 主要试剂TaKaRa RNAiso Plus、反转录试剂盒TaKaRa PrimerScriptTMRT reagent Kit with gDNA Eraser、ExTaq酶、dNTPs与SYBRⅡ均购自宝生物工程(大连)有限公司;DEPC、三氯甲烷、异丙醇、无水乙醇为国产分析纯试剂。实时荧光定量PCR仪为Bio-Rad CFX96TM。
1.2 方法
1.2.1 荧光定量PCR引物设计参照GenBank中注册的猕猴OCLN,CD81及内参β-actin基因序列(序列号分别为XM_002804403.1,NM_001266249.1与NM_001033084.1),分别选取种属间保守序列,参考PCR引物设计原则,借助计算机软件Primer 6设计扩增引物。OCLN上游引物:5′-ATGACCCGCAGACCACCAA-3′,下游引物:5′-CCAGGAAGCCGACGAACATC-3′;CD81上游引物:5′-ATGACCCGCAGACCACCAA-3′,下游引物:5′-CCAGGAAGCCGACGAACATC-3′;β-actin上游引物:5′-AAGGAGAAGCTGTGCTACG-TCG-3′,下游引物5′-TGCCCAGGAAGGAAGGTT-
G-3′。引物由上海英骏生物技术有限公司合成。
1.2.2 标本采集解剖猕猴,采集肝脏、脾脏、肺脏、淋巴结和脊髓,冻存于-20 ℃冰箱中以备使用。
1.2.3 RNA提取
取0.1 g组织,置于预冷的经DEPC水处理的灭菌研钵中,液氮研磨后加入1 mL Trizol混合,并移至2 mL灭菌离心管中,按照操作说明分别从猕猴的肝脏、肺脏、脾脏、淋巴结以及脊髓组织中提取总RNA。离心收集培养好的Huh 7.5.1细胞,提取细胞总RNA。
1.2.4 反转录获得模板cDNA分别将提取的猕猴组织和细胞的总RNA用RNase-free水溶解后,采用以下体系去除基因组DNA:5×gDNA Eraser Buffer 2 μL, gDNA Eraser 1 μL, 总RNA 3 μL, RNase Free dH2O 4 μL配置成10 μL体系,42 ℃水浴2 min,置于冰上冷激。抽取上述反应液,采用反转录体系:反应液10 μL, PrimerScriptRT Enzyme Mix1 1 μL,RT Primer Mix 1 μL,5×PrimerScript Buffer 4 μL,RNase Free dH2O 4 μL配置成20 μL体系,37 ℃保温15 min,85 ℃加热5 s,置于冰上冷激后得到可用于荧光定量PCR的cDNA。
1.2.5 PCR扩增为鉴定引物的扩增特异性,采用上述合成的特异性扩增引物,以反转录获得的cDNA为模板,分别进行丙肝感染关键基因OCLN和CD81、内参基因β-actin的PCR扩增。将获得的PCR产物进行1.2 %琼脂糖电泳,并将PCR产物送至上海生工测序。
1.2.6 实时荧光定量PCR对5只猕猴的各个组织cDNA进行定量扩增,采用25 μL荧光定量体系如下:cDNA模板2 μL, SYBR ®PremixExTaqTMⅡ 12.5 μL,上下游引物各1 μL,RNase free dH2O 8.5 μL。反应条件:95 ℃预变性3 min,95 ℃变性10 s,58.6 ℃延伸30 s,72 ℃复性30 s,40个循环。
2 结果与分析
2.1OCLN,CD81与β-actin基因PCR扩增结果与测序分析用设计的引物进行PCR扩增,分别从猕猴组织细胞系中得到约169,126与179 bp的目的片段,与预期的条带大小一致,见图1。将PCR产物送检测序,将其测序结果与GenBank进行BLAST比对,结果证实该扩增产物与基因模板序列同源性为100%。
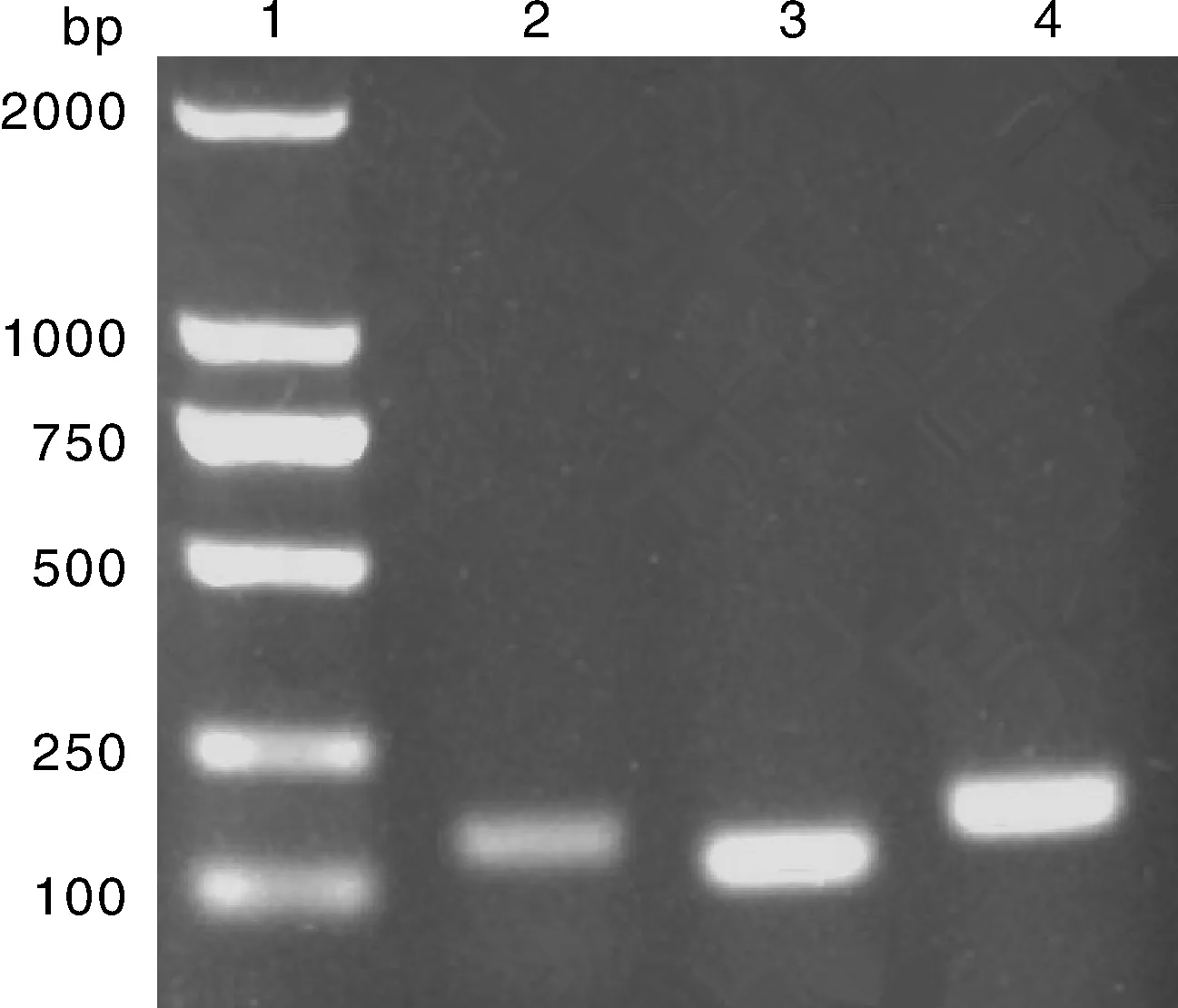
泳道1为DL 2 000 marker; 泳道2为以猕猴川西亚种组织cDNA为模板扩增出的β-actin片段(169 bp); 泳道3为以猕猴川西亚种组织cDNA作为模板扩增出的CD81片段(126 bp); 泳道4为以猕猴川西亚种组织cDNA作为模板扩增出的OCLN片段(179 bp)。图1 OCLN和CD81基因PCR扩增结果Fig.1 PCR amplification product of OCLN and CD81 genes
2.2 实时荧光定量PCR
采用实时定量PCR对猕猴川西亚种的肝脏、脾脏、肺脏、淋巴结和脊髓组织,以及Huh7.5.1细胞中的OCLN,CD81和β-actin进行扩增,所有的样品均出现单一的S型扩增曲线,且重复性好,结果见图2。
2.3 熔解曲线分析
所有样品中OCLN的熔解温度为79.5~80.0 ℃,差值范围≤0.5 ℃,属于正常差值范围,CD81的熔解温度为86 ℃,β-actin的熔解温度为88.5 ℃。所有基因的熔解曲线有一个特异峰,说明实验所用引物特异性好,见图3。
2.4 相对表达量分析
采用Fold=2-△△CT对数据进行处理。结果发现,在Huh7.5.1中,CD81的表达量是OCLN的3.84倍(图4)。猕猴不同组织中OCLN基因表达量明显不同,一般肺部表达量最高,而淋巴结中最少,均极显著低于Huh7.5.1中两种基因的表达量(P<0.01)。针对OCLN在肝、脾和肺这3个主要脏器的表达量分析发现,5只猕猴符合以下规律:脾<肝<肺。淋巴结中OCLN的表达量最低,仅为Huh 7.5.1细胞中表达量的0.13%,远远低于Huh 7.5.1细胞中的表达量。猕猴肝脏中OCLN的表达量仅为Huh 7.5.1细胞中OCLN表达量的2.0%~3.5%,所检测的所有肝脏样品中,最高表达量是最低表达量的2.2倍,个体之间具有明显差异(图5、图6)。而CD81在猕猴肝脏中的表达量最高,为Huh 7.5.1细胞中CD81表达量的4%~14%,最高表达量是最低表达量的3.5倍,不同个体之间差异明显。在脊髓中表达量最低,脊髓中的CD81表达量仅为Huh 7.5.1细胞中表达量的1%,个体差异不明显。与Huh 7.5.1细胞中CD81的表达量相比,淋巴结与脾脏中CD81表达量分别为Huh 7.5.1细胞的1%~6%与2%~5%,两者表达量相当(图5、图6)。
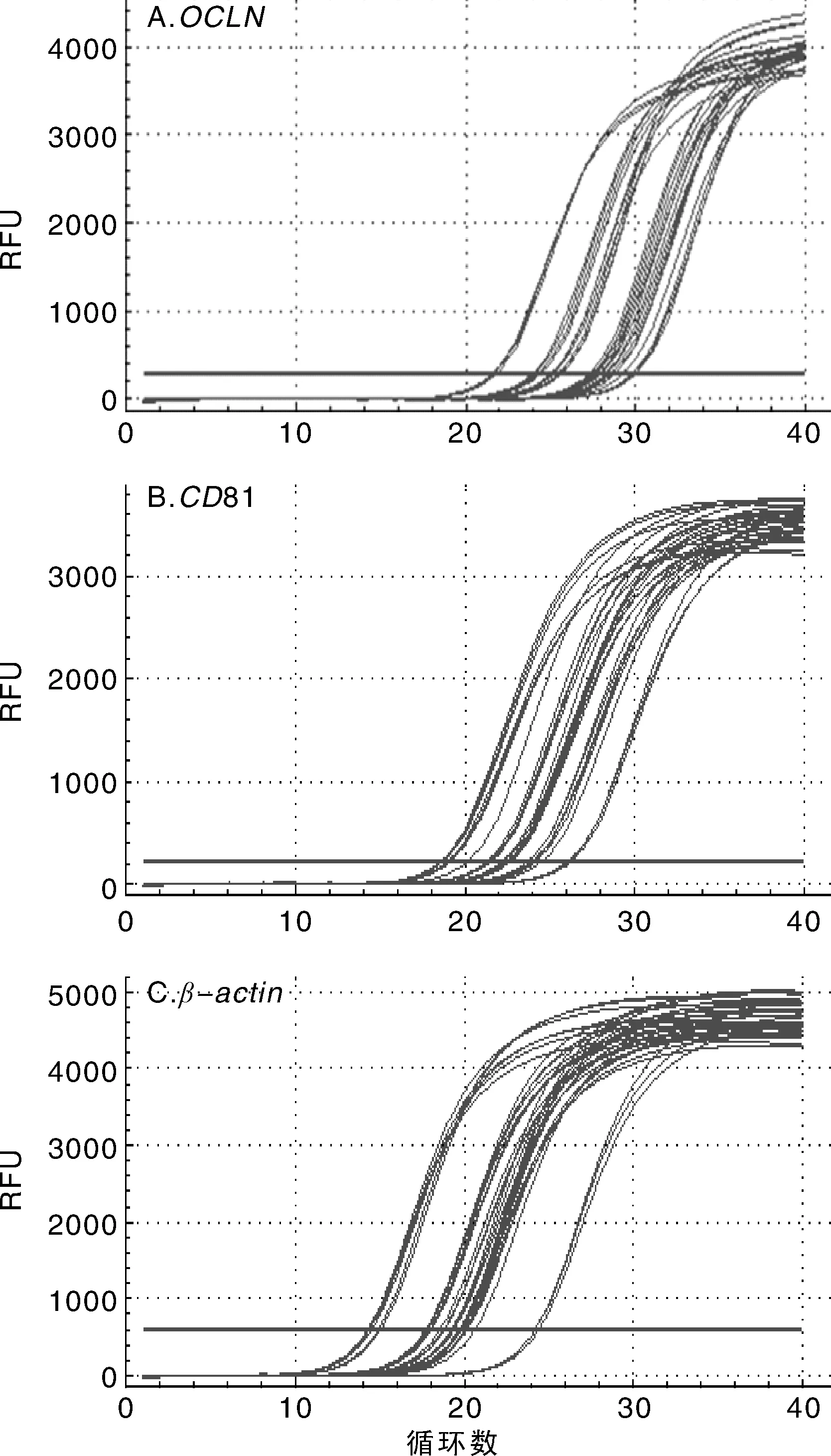
图2 Huh7.5.1细胞系与猕猴不同组织中OCLN,CD81和β-actin扩增曲线Fig.2 Quantitative PCR amplification plot of OCLN, CD81 and β-actin mRNA in Huh7.5.1 cells and different tissues of macaque
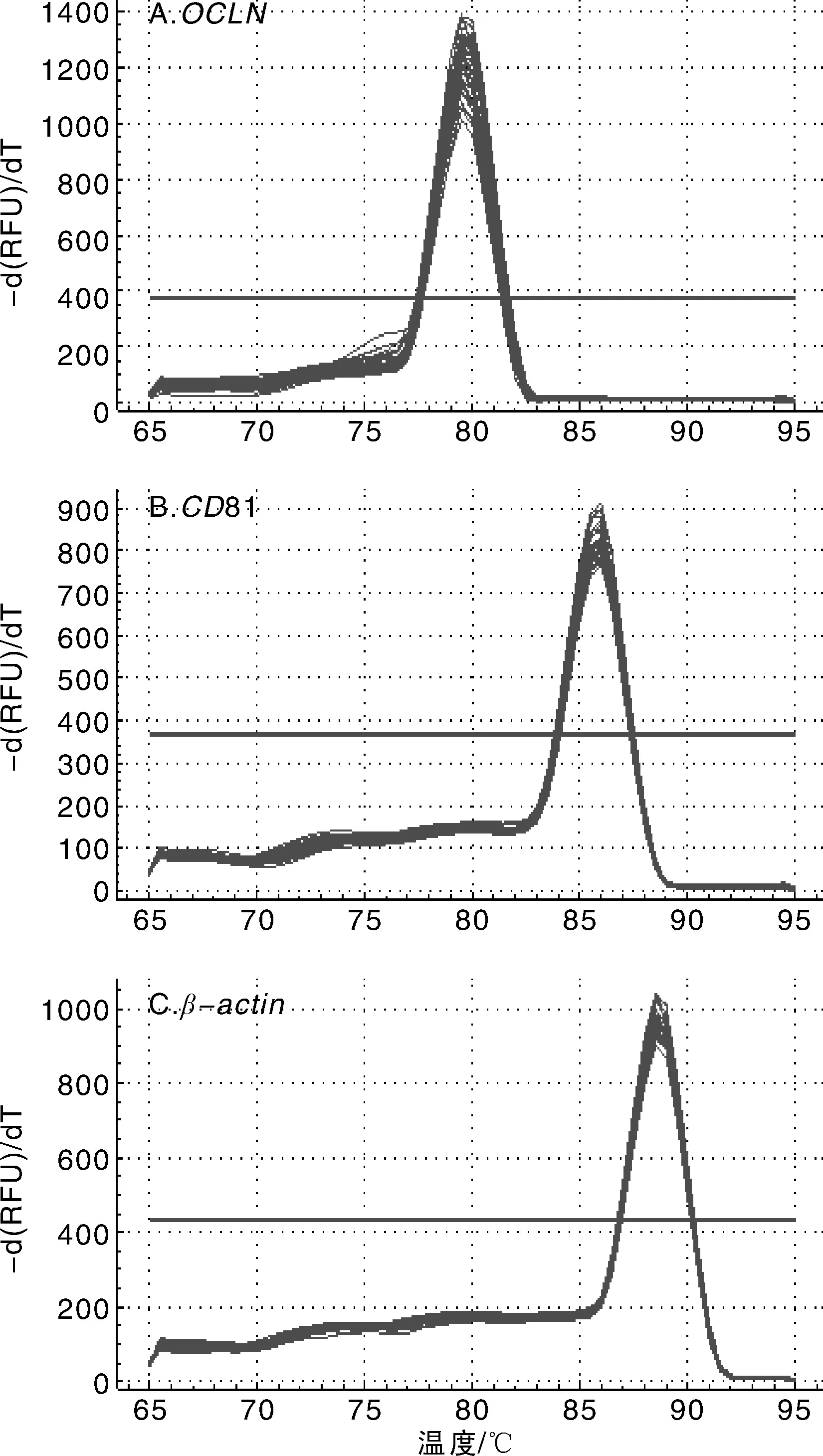
图3 OCLN,CD81和β-actin熔解曲线Fig.3 Melting curve of OCLN, CD81 and β-actin

图4 Huh 7.5.1中OCLN与CD81的表达量Fig.4 The expression of OCLN and CD81 in Huh 7.5.1

图5 OCLN和CD81在Huh7.5.1与不同猕猴组织中的表达量Fig.5 The expression level of OCLN and CD81 in Huh7.5.1 and different macaque tissues

图6 不同猕猴个体肝脏中OCLN和CD81的表达量Fig.6 The expression level of OCLN and CD81 in macaque liver
对比发现,猕猴组织中OCLN与CD81的表达量均远低于Huh7.5.1细胞中的基因表达量。
3 讨论
OCLN蛋白是介导丙型肝炎病毒(HCV)入侵细胞并且完成完整增殖周期的关键蛋白,其异常表达会降低HCV以及HCVpp对人体细胞的感染[3,10]。丙型肝炎由于其多突变型而难以治愈,疫苗研制困难。由于HCV具有感染物种特异性,从而大大限制了实验动物模型种类[11],故寻找可替代的染病状态与人类尽可能接近的新的模型动物是近年来研究的热点。普遍认为黑猩猩模型是最为理想的动物模型,因为黑猩猩的HCV感染、致病和免疫等过程与人类相似,但是由于伦理争议而被限制使用。尽管已有的树鼩模型和人鼠嵌合肝模型等均可用于HCV感染的研究,但是由于感染阳性率低,肝脏病理变化轻微等原因而限制其使用[12-13]。
Kohaar等[6]研究发现,OCLN在人体不同组织的表达具有明显的差异。通过实验我们发现,猕猴川西亚种也具有该特点,但是组织中OCLN的表达量均显著低于Huh 7.5.1中OCLN的表达量。除此之外,OCLN在猕猴川西亚种不同组织之间表达量明显不同,这可能与HCV感染的组织特异性有关。本研究结果表明,OCLN在猕猴组织中表达量大小为脾<肝<肺,该结果也与Kohaar等[6]测得人类的OCLN表达规律相符合。Sourisseau等[14]研究发现,豚尾猴OCLN与猕猴OCLN的E2区氨基酸排布一致,两者与人类OCLN的E2区仅有一个氨基酸差别,但是该差别并不影响HCV入胞过程,OCLN的表达量低则是限制HCV感染豚尾猴的原因。因此推测,猕猴OCLN可能具有介导HCV感染细胞的能力,但是由于其表达量极低,HCV感染可能受到限制。
Sourisseau等[14]发现,豚尾猴与人类CD81共有4个氨基酸的差别,而该差别导致的结构差异则是限制HCV入胞以及限制HCV感染物种特异性的重要因素,其表达量低也是影响HCV感染豚尾猴肝细胞的因素。而猕猴与豚尾猴CD81的HCV结合区域氨基酸排序一致,因此推测猕猴CD81与HCV结合过程可能受到限制。我们发现,CD81在猕猴组织中均有表达,但表达量显著低于Huh 7.5.1中的表达量,CD81在肝脏的表达量相比其他组织最高,也仅为4%~14%。我们推测,猕猴CD81表达量低以及其结构变异可能阻碍HCV结合过程,可能是导致猕猴无法直接感染HCV的因素之一。Bitzegeio等[15]发现,小鼠CD81与HCV的结合效率低,但是突变型HCV则可以有效地与小鼠CD81结合,进而感染细胞。因此,可在提高猕猴CD81表达量的同时,培养出适应猕猴CD81的突变型HCV,用以感染猕猴进行造模。
Dorner等[16]利用转基因技术构建了能够表达人类OCLN,CD81分子的转基因小鼠模型,Ouyang等[17]建立了具有完整免疫功能的人鼠嵌合肝模型,Wu等[18]通过免疫组化证明了在该模型中有人类蛋白的存在,并且支持HCV复制及感染。但是上述两种模型均不能用于免疫机制方面的研究[19-21]。与小鼠等实验动物相比,猕猴拥有与人更加相近的基因背景和生理状态,是良好的用于人类疾病造模的动物。针对OCLN在猕猴川西亚种的组织中表达量相比于Huh 7.5.1极低,可能不易感染HCV这一问题,或许可以从基因角度进行研究,通过提高OCLN的表达量来解决。由于结构差异导致的猕猴CD81可能无法有效与HCV结合这一问题,可以通过转基因等技术解决,从而得到可以直接感染HCV的猕猴个体。
[1] ANDO-AKATSUKA Y, SAITOU M, HIRASE T, et al. Interspecies diversity of the occludin sequence: cDNA cloning of human, mouse, dog, and rat-kangaroo homologues[J].TheJournalofCellBiology, 1996, 133(1): 43-47.
[2] XU SY. Progress of tight junction associated protein occludin[J].InternationalJournalPediatrics, 2012,39(5): 451-454.
[3] LIU S, YANG W, SHEN L, et al. Tight junction proteins claudin-1 and occludin control hepatitis C virus entry and are downregulated during infection to prevent superinfection[J].JournalofVirology, 2009, 83(4): 2011-2014.
[4] COCCHI F, DEVICO A L, YARCHOAN R, et al. Higher macrophage inflammatory protein (MIP)-1α and MIP-1β levels from CD8+T cells are associated with asymptomatic HIV-1 infection[J].ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica, 2000, 97(25): 13812-13817.
[5] PLOSS A, EVANS M J, GAYSINSKAYA V A, et al. Human occludin is a hepatitis C virus entry factor required for infection of mouse cells[J].Nature, 2009, 457(7231): 882-886.
[6] KOHAAR I, PLOSS A, KOROL E, et al. Splicing diversity of the human OCLN gene and its biological significance for hepatitis C virus entry[J].JournalofVirology, 2010, 84(14): 6987-6994.
[7] TONG Y, ZHU Y, XIA X, et al. Tupaia CD81, SR-BI, claudin-1, and occludin support hepatitis C virus infection[J].JournalofVirology, 2011, 85(6): 2793-2802.
[8] BUKH J. A critical role for the chimpanzee model in the study of hepatitis C[J].Hepatology, 2004, 39(6): 1469-1475.
[9] 丁红方,马科,田德英.丙型肝炎病毒肝外组织感染与肝外肿瘤的关系[J]. 中西医结合肝病杂志, 2008, 18(4): 223-224.
[10] PLOSS A, EVANS M J. Hepatitis C virus host cell entry[J].CurrentOpinioninVirology, 2012, 2(1): 14-19.
[11] MICHTA M L, HOPCRAFT S E, NARBUS C M, et al. Species-specific regions of occludin required by hepatitis C virus for cell entry[J].JournalofVirology, 2010, 84(22): 11696-11708.
[12] PLOSS A, RICE C M. Towards a small animal model for hepatitis C[J].EMBOReports, 2009, 10(11): 1220-1227.
[13] 夏俊珂,王玲,庄辉. 丙型肝炎病毒实验感染动物模型研究进展[J]. 中国病毒病杂志, 2013(5): 321-325.
[14] SOURISSEAU M, GOLDMAN O, HE W, et al. Hepatic cells derived from induced pluripotent stem cells of pigtail macaques support hepatitis C virus infection[J].Gastroenterology, 2013, 145(5): 966-969. e7.
[15] BITZEGEIO J, BANKWITZ D, HUEGING K, et al. Adaptation of hepatitis C virus to mouse CD81 permits infection of mouse cells in the absence of human entry factors[J].PLoSPathogens, 2010, 6(7): e1000978.
[16] DORNER M, HORWITZ J A, ROBBINS J B, et al. A genetically humanized mouse model for hepatitis C virus infection[J].Nature, 2011, 474(7350): 208-211.
[17] OUYANG E C, WU C H, WALTON C, et al. Transplantation of human hepatocytes into tolerizedgenetically immunocompetent rats[J].WorldJournalofGastroenterology, 2001, 7(3): 324-330.
[18] WU G Y, KONISHI M, WALTON C M, et al. A novel immunocompetent rat model of HCV infection and hepatitis[J].Gastroenterology, 2005, 128(5): 1416-1423.
[19] MEULEMAN P, LEROUX-ROELS G. The human liver-uPA-SCID mouse: a model for the evaluation of antiviral compounds against HBV and HCV[J].AntiviralResearch, 2008, 80(3): 231-238.
[20] CEREC V, GLAISE D, GARNIER D, et al. Transdifferentiation of hepatocyte-like cells from the human hepatoma HepaRG cell line through bipotent progenitor[J].Hepatology, 2007, 45(4): 957-967.
[21] MEULEMAN P, LIBBRECHT L, WIELAND S, et al. Immune suppression uncovers endogenous cytopathic effects of the hepatitis B virus[J].JournalofVirology, 2006, 80(6): 2797-2807.
(责任编辑 张 韵)
Distribution of HCV infection related OCLN and CD81 in different tissues of Macaca mulatta lasiotis
HE Liu1,CHEN Zheng-li1,2,LUO Qi-hui1,2,*,DENG Juan1,XIA Yu1,SHI Liang-qin1,CHENG An-chun2
(1.LaboratoryofExperimentalAnimalDiseaseModel,CollegeofVeterinaryMedicine,SichuanAgriculturalUniversity,Wenjiang611130,China; 2.KeyLaboratoryofAnimalDiseaseandHumanHealthofSichuanProvince/CollegeofVeterinaryMedicine,SichuanAgriculturalUniversity,Ya’an625014,China)
The study on hepatitis C virus (HCV) was restricted by lack of suitable animal model.Macacamulattalasiotis, known as macaque which has the similar genetic background with human, may be a suitable animal model for studying HCV infection. By testing the expression level of species tropism geneOCLNandCD81 in different macaque tissues, the possibility of using macaque as an effective animal model was explored with the expression level of HCV-susceptible human liver cancer cell line of Huh 7.5.1 as the control. The results showed thatOCLNandCD81 were expressed in liver, spleen, lungs, lymph nodes and spinal cord of macaque, but the expression levels were significantly lower than that in Huh 7.5.1(P<0.01). The expression ofOCLNin liver was lower than that in lung, butCD81 expression in liver was higher than other tissues. In addition, the expression ofOCLNin different tissues of macaque showed a similar pattern as that in human, with a decreasing order of lung>liver>spleen. It was concluded that the low expression level ofOCLNandCD81 in macaque could lead to the ineffective combination of HCV and target tissue, which was possibly related to the inefficiency of macaque as a suitable animal model for HCV. In future, the expression level of OCLN and CD81 in macaque may be enhanced by genetic manipulation, so as to improve the HCV-infection of macaque.
HCV; OCLN; CD81;Macacamulattalasiotis; animal model
http://www.zjnyxb.cn
10.3969/j.issn.1004-1524.2016.01.08
2015-07-16
国家科技支撑计划课题(2014BAI03B01);国家重大科学仪器设备开发专项(2013YQ49085906);四川省青年科技创新研究团队项目(2013TD0015)
何柳(1994—),女,河南许昌人,在读本科生,研究方向为实验动物疾病模型方向。E-mail:heliu.july@gmail.com
*通信作者,罗启慧,E-mail:lqhbiology@163.com
S852
A
1004-1524(2016)01-0044-07
浙江农业学报ActaAgriculturaeZhejiangensis, 2016,28(1):44-50
何柳,陈正礼,罗启慧,等. 丙肝感染宿主亲嗜性基因OCLN和CD81在猕猴川西亚种组织中的分布[J].浙江农业学报,2016,28(1):44-50.
猜你喜欢
热带亚热带植物学报(2022年4期)2022-08-18
红领巾·成长(2022年6期)2022-07-23
小猕猴学习画刊(2022年10期)2022-01-01
昆虫学报(2021年11期)2021-12-08
——和田盘羊
野生动物学报(2021年4期)2021-11-14
趣味(语文)(2021年11期)2021-03-09
金桥(2020年9期)2020-10-27
食品与发酵工业(2019年18期)2019-10-11
小猕猴智力画刊(2019年4期)2019-05-08
小猕猴智力画刊(2019年3期)2019-04-19
