
热带雨林林内小生境对木奶果和染木种子萌发与幼苗存活的影响
2016-10-14 05:22:41刘明航
西北植物学报 2016年8期
刘明航,叶 娟,文 彬
(1 中国科学院西双版纳热带植物园综合保护中心,云南勐腊 666303; 2 中国科学院大学,北京 100049)
热带雨林林内小生境对木奶果和染木种子萌发与幼苗存活的影响
刘明航1,2,叶 娟1,2,文 彬1*
(1 中国科学院西双版纳热带植物园综合保护中心,云南勐腊 666303; 2 中国科学院大学,北京 100049)
森林生态系统中小生境的差异可能影响种子萌发与幼苗的建立,并形成物种不同的分布格局。为探讨生境异质性对植物分布的影响,该研究在西双版纳热带季节性雨林中选取5个具有不同海拔和地形的小生境,人工散布木奶果与染木种子,定期监测种子萌发与幼苗存活,同步记录小生境的近地面(距地面3 cm)温度、近地面(3 cm)空气湿度和表层土壤(5 cm)含水量变化,并在实验室开展种子脱水和不同水势条件下的种子萌发实验。结果表明:(1)实验所选的5个小生境中,沟底地带的土壤水分含量在一年中始终都高于其他小生境;5个小生境的空气相对湿度在雨季与雾凉季都接近饱和,只有在干热季出现明显的差别,即随海拔的升高,空气湿度下降。同时,在旱季,近地面温度随海拔的升高而升高, 出现1~2个月的山地逆温现象。(2)木奶果与染木具有不同的种子传播与萌发规律,而且染木种子在不同生境下,其种子萌发率和幼苗存活率有显著的差异。其中:木奶果种子在雨季成熟散布并快速萌发,以幼苗的形式度过干热季,其种子可以在相对较低的水势下萌发;染木种子在雨季末期成熟并进入休眠期,以种子的形式度过干热季,其种子相对较抗脱水。研究认为,地形是决定西双版纳热带季节性雨林生境异质性的重要因子,小生境在温度和水分方面的差异在干热季时尤为明显,并对木奶果和染木种子的萌发或幼苗存活产生了重要影响, 而木奶果与染木种子自身的特性使其对于西双版纳季节性的干旱具有不同的适应对策。
生境异质性;热带雨林;地形;种子萌发;幼苗存活
生境是土壤、地形、温度、降水等诸多环境因子的综合体,是植物生存的载体。生境的空间异质性是生态系统的一个普遍特征[1]。由于生境异质性的存在,不可避免地导致群落内物种组成结构的小尺度差异。生境异质性程度越高,能形成的小生境就越多,从而在更大范围内满足不同物种的需求。自然界的生物分布既不是均匀的,也不是随机的,而是聚集成斑块,形成梯度或其他形式的空间结构[2]。生境过滤与生物间相互作用作为群落构建的2个基本驱动力,影响特定群落结构的形成[3-5]。多数的研究表明,生境过滤是塑造物种共存的主要动力[6]。种子是有花植物生命过程中唯一可以被远距离传播的、独立的生命个体。作为植物生活史中的重要环节,种子萌发与幼苗阶段通常是植物更新的瓶颈期,此阶段死亡率高,幼苗长成为成年植株后,其对不良环境的耐受性提高,因此种子萌发成功与否和早期幼苗能否存活能影响种群数量和物种分布格局。有研究表明,种子萌发与幼苗存活阶段所处的微环境对于植物分布具有重要影响[7]。以往关于环境异质性、物种和生境的关联性对物种多样性影响的研究大都仅限于胸径≥1 cm 的木本植物(非幼苗个体)[8-10],鲜有研究探讨这些过程对树木生活史早期阶段的影响[11]。
西双版纳地处热带亚洲的北缘,这里的热带雨林是地球上位置最靠北端的热带雨林,同时又是中国目前面积最大、保存最为完好的热带雨林。西双版纳热带雨林具有明显的季节性,称为热带季节性雨林,但具有与赤道低地热带雨林几乎一样的群落结构和生态外貌特征,是亚洲热带雨林的一个类型。由于发生在季风热带北缘纬度和海拔的极限条件下,受到季节性干旱和热量不足的影响,这些特征又有别于赤道低地的热带雨林[12]。不同于赤道地区典型的热带雨林,西双版纳热带雨林明显地受到地形的影响,主要分布在低海拔的沟谷地区,称为“沟谷雨林”,而且物种的斑块分布现象十分突出。
木奶果(Baccaurearamiflora)和染木( Saprosmaternata)均为西双版纳热带季节性雨林中广泛分布的物种,且具有相似的分布格局,即绝大多数个体集中分布在沟底和低坡、中坡,很少分布在坡顶。基于西双版纳季节性雨林具有的季节性干旱与热量不足的特点,本实验探讨了林内不同的小生境下,温度与水分条件对木奶果和染木种子萌发与幼苗存活的影响,以期在生境异质性-种子萌发-分布格局三者之间建立联系,从而为西双版纳季节性雨林植物组成结构提供一定的解释。
1 材料和方法
1.1 研究地区概况
西双版纳地区一年可以分为3个季节:5~10月份为雨季;11月至翌年2月为雾凉季,降雨较少,但有大雾,可以部分弥补降雨的不足;3~4月为干热季,气温升高,干旱少雨[13]。西双版纳地区年平均降水量为1 560 mm,约80%的降水集中在5月到10月的雨季[14]。野外观测点设置在勐腊县补蚌村西双版纳热带森林动态监测样地附近,该样地的地理位置为21°36′42″~58″N,101°34′26″~47″E。样地沟谷底部及两侧坡面是望天树(Shoreawantianshuea)占优势的热带季节性雨林[15],中下层乔木主要有绒毛番龙眼(Pometiatomentosa)、假海桐(Pittosporopsiskerrii)、云树(Garciniacowa)、蚁花(Mezzettiopsiscreaghii)、染木(Saprosmaternata)等[16]。5个观测点都是在林冠郁闭(故可以忽略光照条件的差异),受人为扰动较少的自然保护区内,其地理信息如表1,与雨林动态监测样地的相对位置如图1。
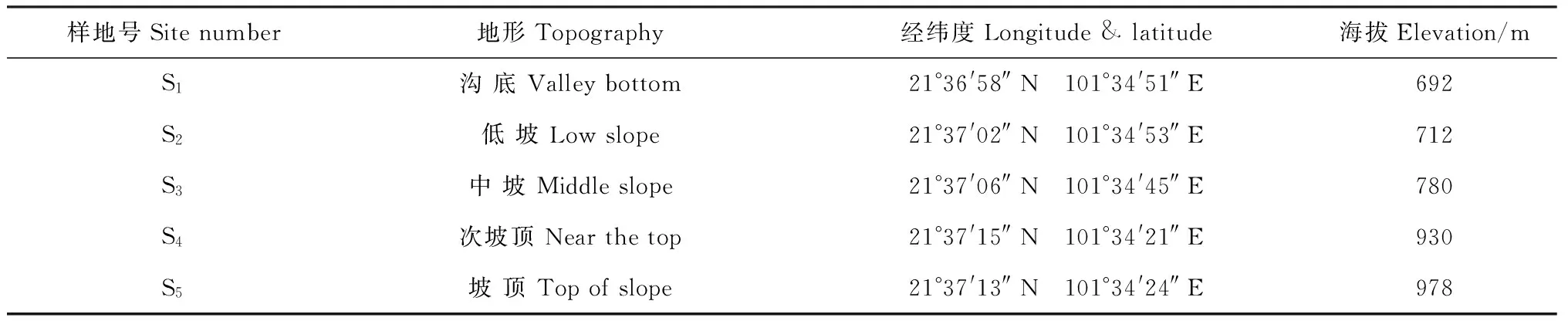
表1 观测点的地理信息
1.2 种子采集与处理
种子采集于观测点附近自然分布的木奶果、染木植株。种子来源覆盖了较多植物个体与较广阔的分布范围,使种子充分代表了此区域内种群,避免遗传因素对实验的影响。实验于2014年进行,木奶果种子8月成熟,染木种子10月成熟,种子成熟后立即采摘。木奶果与染木种子的初始含水量分别为58.3%±0.01%和54.6%±2.61%(鲜重为基础计算)。种子散布前去除种子外附属物如果肉、假种皮,处理好后尽快散布于实验点。用于实验室实验的种子置于封口的聚乙烯袋内,10 ℃储藏,在1周之内使用。由于染木种皮对其种子萌发有抑制作用,用于实验室实验的种子在实验前先剥除种皮。
1.3 小生境温度、湿度和土壤含水量监测
在每个观测点距近地面(高3 cm处)放置1枚ibutton温湿度记录仪(DS1923, Dallas公司,温度精度为0.5 ℃,湿度精度为0.6%),以15 min 1次自动记录,每月月底下载数据1次。同时在每个观测点采集土样5份,即在每个观测点4个角5 m外及正中各选 1个取样点,先去除近地面落叶,再用环形刀取5 cm深度土样,置于专用的铝盒内,盖紧,密封,带回实验室,挑除小石子、枯枝落叶等,103 ℃烘箱内烘干48 h至恒重,利用重量差值计算土壤含水量,以湿土重量为基数表示土壤含水量。
1.4 野外种子萌发与幼苗存活实验
每个观测点每种种子设置6个重复实验点,即面积为40 cm×40 cm的6个小样方,小样方之间距离为20 cm,每个小样方150粒种子,总计900粒。先疏松表层土壤,再将种子平铺于地面,为防止萌发阶段的动物取食,用铁丝网罩住种子并用钢钉固定。待大部分种子成苗后移除铁丝网,每月底记录种子萌发数及幼苗存活数。
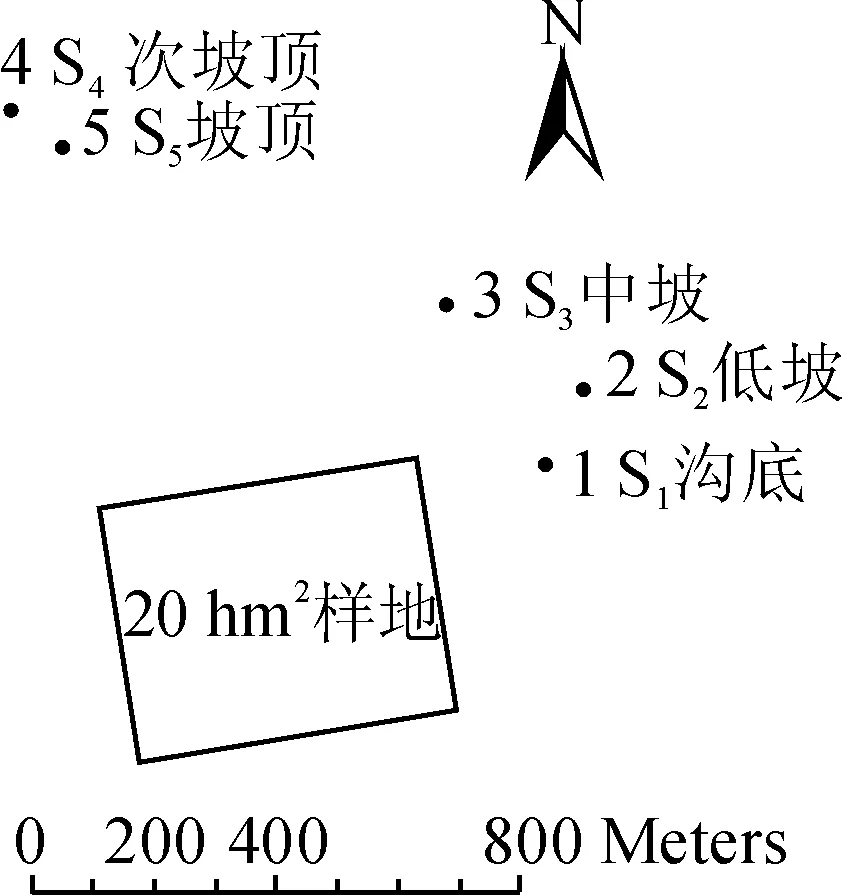
图1 观测点与样地相对位置Fig. 1 Observation sites in relation to the Xishuangbanna tropical seasonal rainforest dynamics plot
1.5 不同水势条件下种子萌发实验
按Michel 的方法[17],将聚乙二醇PEG8000(Sigma公司)溶解于蒸馏水,配成水势0、-0.2至-1.0 MPa的PEG溶液,该溶液可模拟种子萌发过程中外界的水势环境。取广口玻璃瓶,瓶底铺1层脱脂棉,把种子放置于脱脂棉上,用不同水势的PEG溶液浸润脱脂棉,溶液量为10 mL,用含透气口的封口膜封口。按每个水势下设置5个(染木)或6个(木奶果)重复,每个重复20粒种子。种子置于人工气候箱(宁波东南仪器厂)内培养,条件为:25 ℃、光照强度20 μmol·m-2·s-1(24 h)、相对空气湿度(RH)80%,每3 d更换1次溶液及脱脂棉,并记录萌发率与生长情况。
1.6 种子在室内条件下的脱水处理
将种子单层平铺于室内阴凉处,缓慢阴干,定期取样,以监测种子含水量与生命力的变化。(1)含水量测定:根据国际种子检验协会(ISTA,2004)推荐的方法,以烘干(103 ℃,17 h)前后种子重量差异计算种子的含水量。每处理含8个重复,每个重复1粒种子。含水量(%)=(鲜重-干重)/鲜重×100%。(2)生命力测定:每处理100粒种子,分成5个重复,播种于含有1%琼脂的培养皿,盖上培养皿盖后,放入人工气候箱内培养(条件同上)。每周记录1次种子萌发情况。萌发周期90 d,90 d后仍未萌发的种子,用刀片解剖开,检验其是否死亡,若未死亡,继续培养,直至确认种子死亡。
根据Samarah等(2009)的方法计算脱水指数来度量脱水速率,公式如下:
SMLR为种子脱水速率指数,n为脱水的梯度次数,SMCi为第i个脱水梯度的含水量,dayi为脱水的天数。
1.7 数据统计分析
以上实验均以胚根长出2 mm为萌发成功,子叶展开为成苗,分别统计萌发率和成苗率。
SPSS17.0在0.05水平上,运用非参数检验中的两个独立样本检验(two-independent sample test)对表层土壤含水量、种子萌发率、幼苗存活率或死亡率进行观测点间两两比较;概率分析(probit analysis)计算缓慢脱水种子50%致死率时的含水量。
2 结果与分析
2.1 观测点小生境温度、湿度和土壤含水量动态变化特征
表层土壤含水量不仅是表示小生境水分条件的一个重要指标,且能从侧面反映降水情况,对种子萌发具有重要影响。实验期内5个观测点土壤含水量变化趋势都呈U(或V)型曲线(图2,A),符合大背景下的气候与降水趋势,即第一年雨季后逐渐下降,进入干热季都达到最低值,进入第二年雨季后逐渐上升。月平均土壤含水量在5个点间有明显差异。其中,S1观测点(沟底)与其它4个观测点有显著的差异(P<0.05),其土壤含水量约有10个月能维持在30%以上,而其余4个点土壤含水量变化曲线互有交错,不存在显著差异(P>0.05);2月至3月份,是土壤水分条件最严峻的时期,S4观测点土壤含水量最低,仅有11.9%,而S1观测点最高,为22.7%。
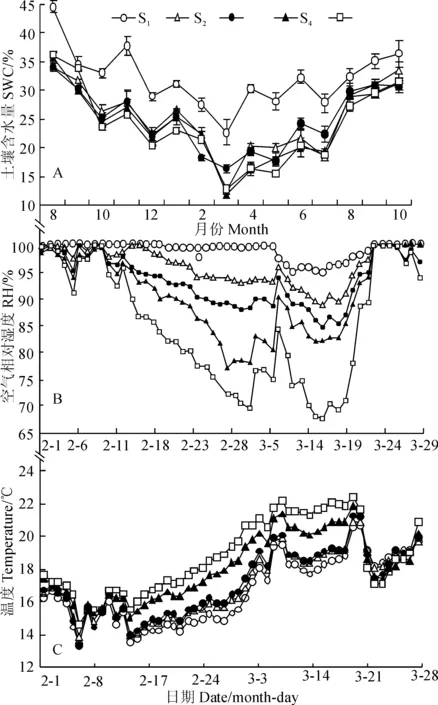
A.每月土壤含水量(平均值±标准误);B. 2月~3月日平均湿度;C.2月~3月日平均温度;S1~S5.观测点(S1.沟底;S2.低坡;S3.中坡;S4.次坡顶; S5.坡顶);下同。图2 各观测点小生境土壤含水量、空气相对湿度和气温动态变化特征A. Monthly soil water content(Means±SE);B. Daily mean relative humidity from Feb. to Mar.;C. Daily mean temperature from Feb. to Mar.;S1-S5 .Observation sites (S1.Valley bottom;S2.Low slope;S3.Middle slope;S4.Near the top;S5.Top of slope); Other figures follow the same diagram formFig. 2 The soil water content, air relative humidity and air temperature at each observation site
同时,近地面的空气相对湿度是衡量生境水分条件的另一个指标,它直接关系到处于地表种子的水分散失速率,进而影响种子生命力。总体而言,热带季节性雨林生境中观测区,除了2~4月份(雾凉季后期和干热季)外,其它时间空气湿度都较为饱和,5个点的每月平均湿度都在90%以上(数据未展示),雨季常达到100%;只有最干旱的2月~3月份2个月时间内,5个观测点日平均湿度呈现明显的分离。随着海拔的上升,从S1到S5观测点的空气相对湿度逐渐下降。且S5观测点最低为67.5%,S1观测点虽然有下降,但最低值仍高达95.0%(图2,B)。
另外,从雨季至雾凉季前期,5个观测点间的日平均气温差异极小(数据未展示),在雾凉季末期与干热季前期和中期(2月~3月),各观测点气温表现出较为明显的分离(图2,C)。观测点在干热季的日平均气温呈现出随海拔升高而升高的山地逆温现象,最高的日平均温度差值为4.5 ℃。翌年雨季,高海拔处温度下降幅度大,逆温现象消失,此后5个观测点间温差较小,大部分时间都在1 ℃以内(数据未展示)。
2.2 木奶果和染木野外小生境种子萌发与幼苗存活特征
本实验所选用的两种种子成熟期存在差别,木奶果在雨季中期(约8月份)果实成熟并散布种子。野外观测表明,散布到5个实验点的种子均萌发迅速,2个月内即可成苗。而染木在雨季末期(约10月份)果熟,种子散布后并不立即萌发,翌年2月开始萌发(S1点),S1至S5观测点染木种子在萌发时间上有明显的“滞后”现象(图3)。
在种子萌发率方面,由于S1观察点地处沟底,雨天可能出现较大的地表径流,少许木奶果种子被雨水冲走(数量不详),观测到成苗率为87.3%±5.8%(以人工散布种子数为基础计算),S2~S5观测点木奶果成苗率分别为91.7%±2.4%、91.8%±2.1%、96.6%±0.5%、92.0%±2.1%;且排除受到干扰的S1观测点,S2~S5观测点木奶果种子初始萌发率无显著差异(P>0.05)。木奶果在水分、温度条件都好的雨季萌发,且萌发迅速而整齐,此时不同小生境下环境因素的差异被抹平,因此萌发率在各生境下无差异。染木种子在S1~S5观测点的萌发率分别为47.3%±4.2%、61.1%±2.3%、48.9%±6.6%、28.8%±7.4%和7.3%±1.9%,且除S1与S3观测点、S3与S4观测点间差异不显著外(P>0.05),其他各点之间萌发率两两互相差异显著(P<0.05),与木奶果不同,在不同生境下,其在萌发阶段即出现幼苗数量的巨大差异。
在幼苗存活率上,木奶果种子萌发整齐且出苗率高,以各观测点实际成苗数量为基数(计为100%),每阶段结束时幼苗死亡的数量除以这一阶段开始时幼苗数量即为该阶段幼苗死亡率(表2)。其中,雨季末期的2个月与雾凉季是木奶果幼苗死亡的高峰期,S3和S4观测点在雨季死亡率超过50%,5个观测点在雾凉季都有一年中最高的死亡率;进入干热季,木奶果所剩存活幼苗已经较少,死亡数较少;监测1年后,各实验点木奶果幼苗死亡量分别占各自总数的91.4%±3.8%、97.9%±1.3%、97.0%±1.3%、96.1%±1.1%和95.6%±4.0%,5个点间幼苗最后存活率无显著差异(P>0.05),只是在沟底(S1)处存活的木奶果数量较其他生境多。对于染木种子而言,野外萌发率差异较大,故以播种量为基数计算其存活率(图3)。其中,至实验第2年1月份,染木幼苗的存活率在S2观测点最高(37.9%±1.7%), S1(25.2%±2.9%)、S3(26.1%±3.6%)和S4(20.0%±5.3%),S5点存活率最低(2.2%±0.9%),S1、S3、S4观测点间两两之间无显著差异(P>0.05),其余观测点的两两间存在差异显著(P<0.05)。可见染木种子萌发率在不同生境下出现差异,且萌发数量强烈影响了幼苗的数量。
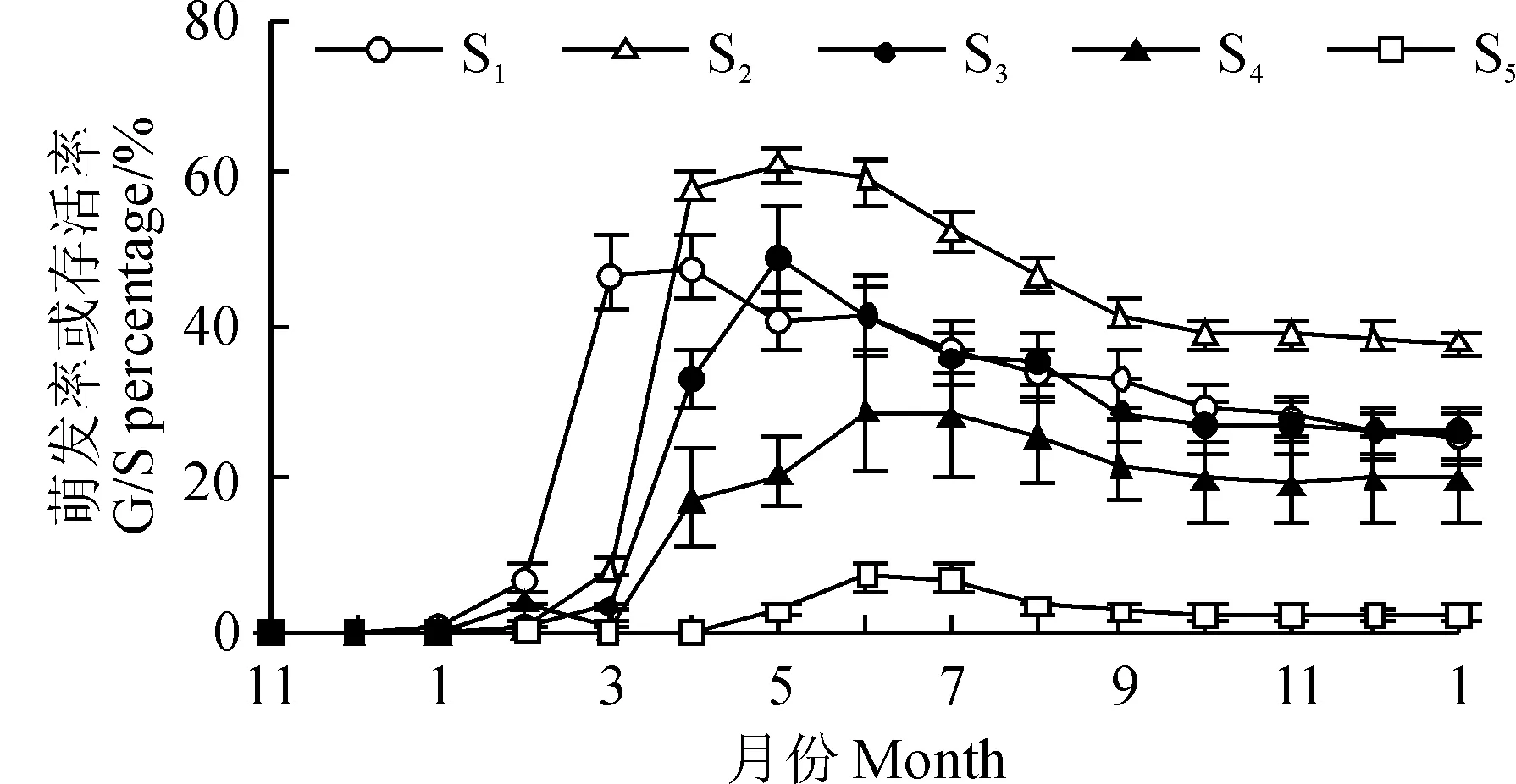
图3 染木种子萌发率或幼苗存活率(平均值±标准误)Fig. 3 S. ternata seed germination percentage and seedling survival percentage(Means±SE)
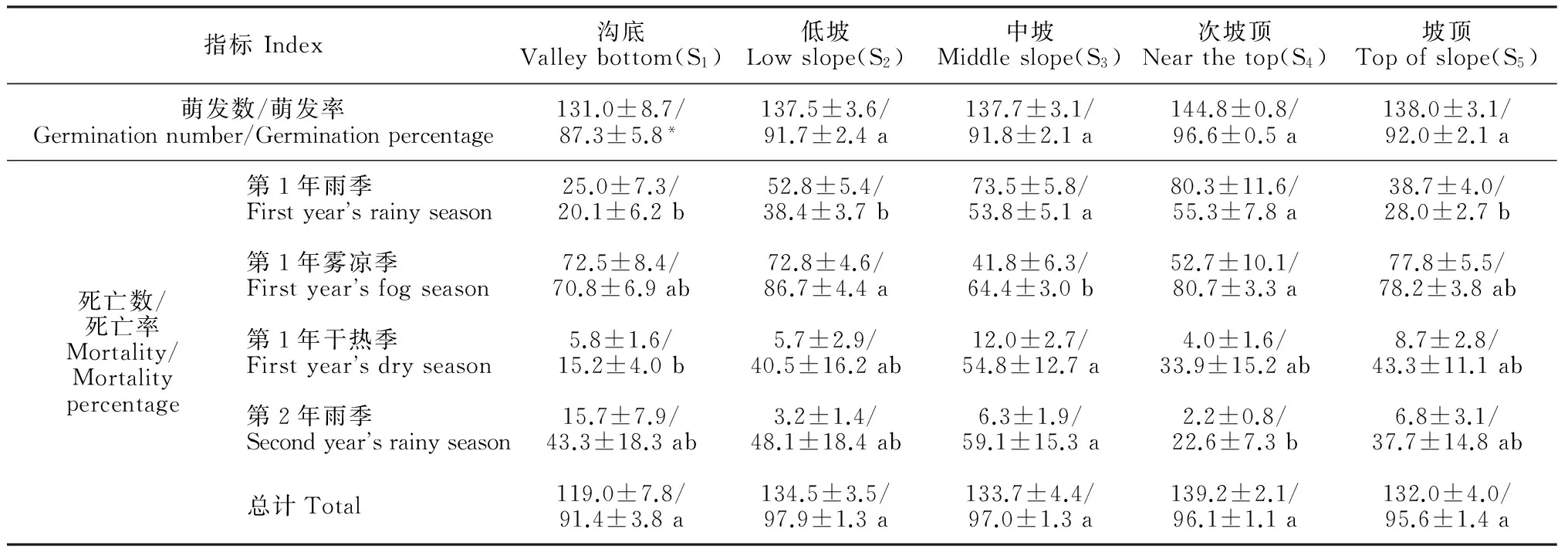
指标Index沟底Valleybottom(S1)低坡Lowslope(S2)中坡Middleslope(S3)次坡顶Nearthetop(S4)坡顶Topofslope(S5)萌发数/萌发率Germinationnumber/Germinationpercentage131.0±8.7/87.3±5.8*137.5±3.6/91.7±2.4a137.7±3.1/91.8±2.1a144.8±0.8/96.6±0.5a138.0±3.1/92.0±2.1a死亡数/死亡率Mortality/Mortalitypercentage第1年雨季Firstyearsrainyseason25.0±7.3/20.1±6.2b52.8±5.4/38.4±3.7b73.5±5.8/53.8±5.1a80.3±11.6/55.3±7.8a38.7±4.0/28.0±2.7b第1年雾凉季Firstyearsfogseason72.5±8.4/70.8±6.9ab72.8±4.6/86.7±4.4a41.8±6.3/64.4±3.0b52.7±10.1/80.7±3.3a77.8±5.5/78.2±3.8ab第1年干热季Firstyearsdryseason5.8±1.6/15.2±4.0b5.7±2.9/40.5±16.2ab12.0±2.7/54.8±12.7a4.0±1.6/33.9±15.2ab8.7±2.8/43.3±11.1ab第2年雨季Secondyearsrainyseason15.7±7.9/43.3±18.3ab3.2±1.4/48.1±18.4ab6.3±1.9/59.1±15.3a2.2±0.8/22.6±7.3b6.8±3.1/37.7±14.8ab总计Total119.0±7.8/91.4±3.8a134.5±3.5/97.9±1.3a133.7±4.4/97.0±1.3a139.2±2.1/96.1±1.1a132.0±4.0/95.6±1.4a
注:季死亡率=季幼苗死亡数/季初始幼苗存活数 × 100%;* 表示沟谷(S1点)一些种子被雨水冲走,数量不详,计为遗失; 只对死亡率进行统计,同行的不同字母表示差异显著(P<0.05)
Note: Mortality percentage on the season = Mortality on the season / Seedling number at the beginning of the season × 100%; * means some seeds at valley bottom (S1) were washed away and the number is unknown. Those seeds was counted as missing; Only mortality percentage is analyzed and different letters in the same row indicate significant difference (P<0.05)
2.3 木奶果和染木在不同水势条件下种子萌发特征
图4显示,木奶果和染木种子萌发率在整个实验过程中均有随着水势的降低而下降的趋势,且相同水势条件下表现为木奶果萌发率高于染木。其中,在30 d内,木奶果萌发率于0 MPa水势时达到99.2%±0.8%,而于-1.0 MPa时仅为 1.7%±1.7%;在90 d的萌发周期里,木奶果萌发率于0~-0.5 MPa水势下都达到90%以上,于-0.6 MPa水势下萌发率为80.8%±3.3%,在-0.8MPa水势下萌发率为29.2%±7.9%。同时,染木种子萌发率在0 MPa水势下最高为84.0%±5.7%,在-0.2 MPa水势下萌发率降至55.0%±3.2%,在-0.5MPa的水势下仍能萌发14.0%±4.5%,但在-0.6MPa水下萌发率仅为2.0%±1.2%。可见,木奶果和染木种子萌发均不同程度受到水分胁迫的影响,木奶果种子对PEG造成的较强水分胁迫有较强的耐受性,而染木种子萌发对外界水分胁迫比木奶果更敏感。
2.4 室内自然脱水对木奶果和染木种子萌发的影响
木奶果种子在室内自然阴干的条件下(24.5 ℃,88.9%RH)的脱水速率指数(SMLR)为17.9,其萌发率在前期脱水过程中始终保持高值,直到阴干96 h后、种子含水量为29.8%以下时萌发率才出现明显的下降(P<0.05)(图5,A)。木奶果种子半致死量(LD50)含水量为25.4%,95%的置信区间为24.0%~27.1%。同时,染木种子在类似的室内自然阴干条件下(22.7 ℃, 85.9%RH)的脱水速率指数为5.4,其萌发率在种子含水量较高时便随着含水量下降而下降(图5,B)。染木种子最大萌发率只有78.3%,半致死量(相当于39%的萌发率)含水量为38.7%,95%的置信区间为32.7%~49.3%。可见,木奶果种子与染木种子相比较具有不抗脱水、但耐脱水的特性,即种子易失水,但在较低含水量下仍能保持高萌发率;染木种子具有抗脱水,但不耐脱水的特性,即种子不易失水,能在野外保持较高含水量,但在含水量下降时不能保持高萌发率。
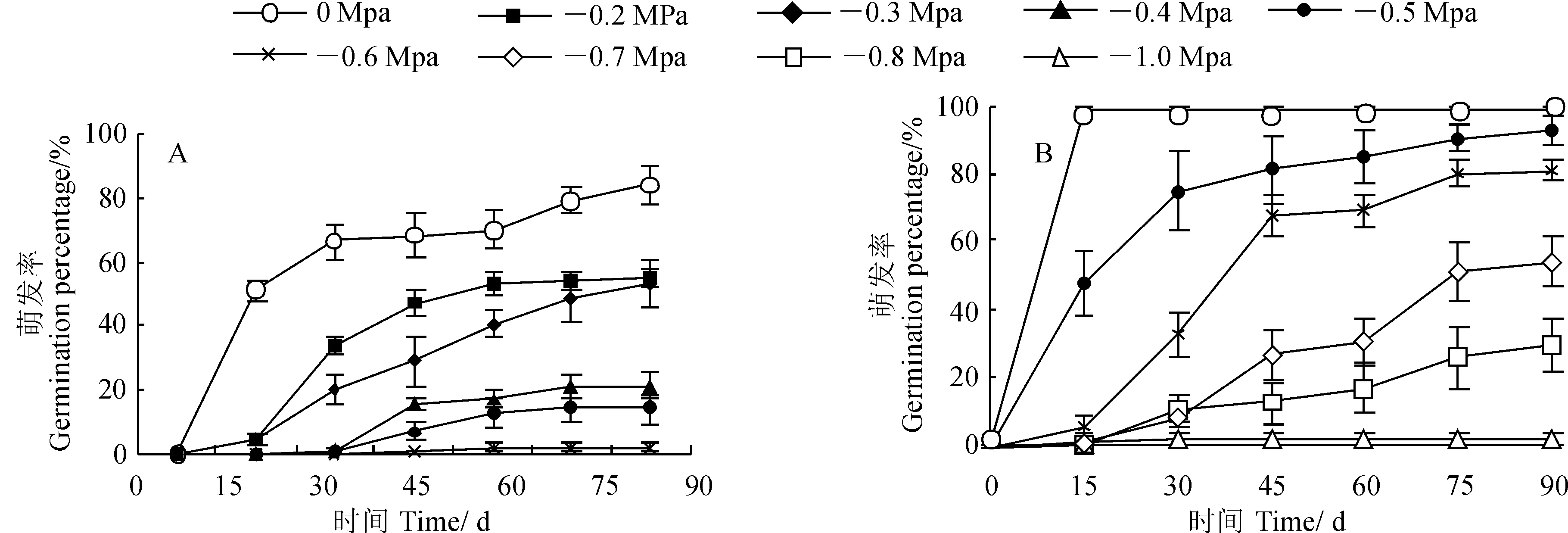
图4 染木(A)和木奶果(B)在不同水势下萌发率(平均值±标准误)Fig. 4 S. ternata (A) and B. ramiflora (B) seed germination percentage under different water potentials(Means±SE)
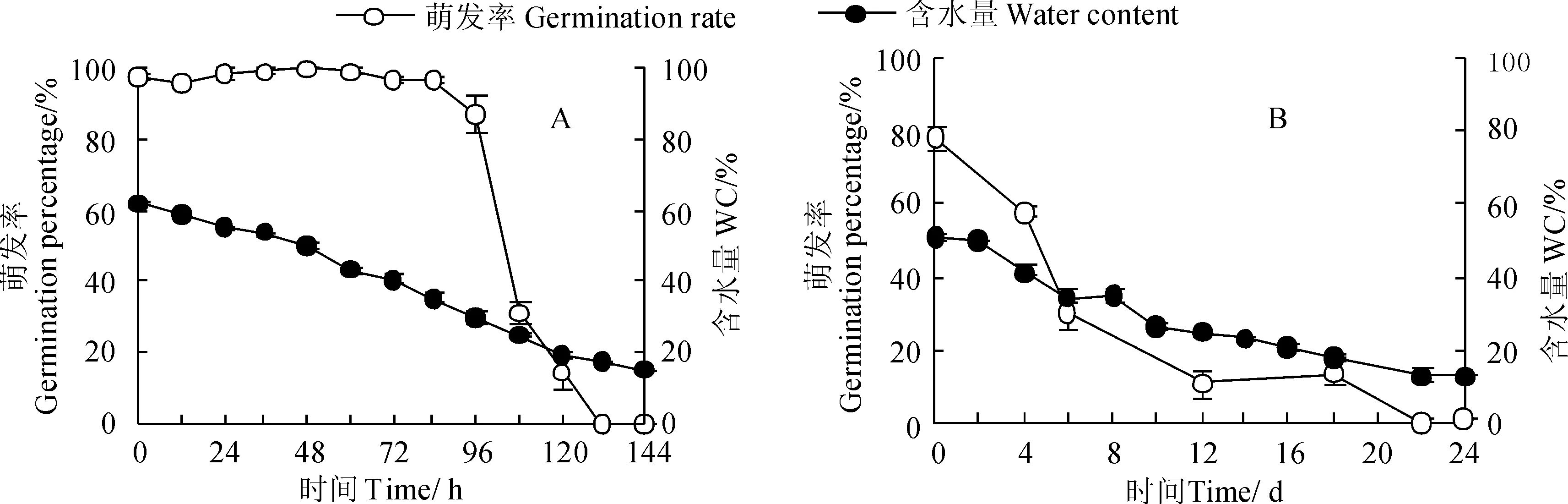
图5 木奶果(A)和染木(B)室内自然脱水下的萌发率(平均值±标准误)Fig. 5 B. ramiflora (A) and S. ternata (B) seed germination rate after dehydration under ambient regimes(Means±SE)
3 讨 论
幼苗生存及生长可能是一个同时受多因素影响的过程[18],地形的坡向和坡度能引起植被在小尺度上呈“虎皮状”的分布格局[19]。水分在很多热带雨林也是影响幼苗存活生长的重要因素[20-21],尤其是幼苗根系只在土壤表层,无法与成年植物竞争[22]。作者据本研究结果推测西双版纳热带季节性雨林林内生境异质性可能主要是由非生物因子主导的,这一点与赤道低地热带雨林主要由生物因子主导的生境异质性存在很大不同。作者猜想,地形可能通过改变小生境的水分条件和温度,从而影响西双版纳热带雨林群落中物种的分布。本研究发现,虽然和其他4个观测点一样,位于沟底的观测点也存在土壤含水量的季节性变化,即雨季结束后各观测点土壤含水量逐渐降低,第二年进入雨季后又逐渐升高,但是比较全年相同时段的数据发现,沟底的土壤含水量始终都是最高的,而且干热季各观测点的差别尤为明显,相差幅度可达15个百分点;不同小生境在湿度与温度方面的差异则主要表现在干热季,位于沟底的观测点在干热季仍有接近饱和的近地面空气相对湿度,而位于坡顶的观测点则空气相对湿度偏低,而气温偏高,日平均空气湿度可低至65%,日平均气温可比沟底高出4.5 ℃。这样,位于坡顶的小生境一方面可能因为较低的土壤含水量限制了种子的萌发,另一方面又可能因为较低的空气相对湿度和较高的温度而加快种子水分的丧失,从而造成种子丧失生命力。
本实验所用的木奶果和染木都是西双版纳热带季节性雨林中的优势种,且具有相似的分布格局,即绝大多数个体都分布在沟底和低坡、中坡,较少有个体在坡顶分布[23];同时,这两种植物的种子都具有对脱水敏感的特点[24-25]。本研究的野外种子萌发实验中,播种到沟底和中坡地段染木种子的萌发率显著高于坡顶,并以低坡观测点最高,有61.1%的种子萌发并成苗,而坡顶观测点最低,成苗率仅7.3%。染木种子的这种野外萌发模式与该物种在热带季节性雨林中的分布格局相吻合,说明在不同地段的种子萌发差别可能是该物种分布格局的成因:染木于雨季末期落果,种子散播后有约3~4个月的休眠期,在野外萌发时正值西双版纳的干热季,而在本实验样地周边范围内的其他实验表明,干热季土壤中水势最低可以到达-1.5 MPa[26]。因此,散布在坡顶的染木种子可能因为偏低的土壤含水量限制了种子的萌发,因为水势胁迫实验证明,该种子对水分胁迫敏感,当水势降低到-0.5 MPa时,绝大部分种子萌发就已经被限制;同时,在干热季坡顶具有低湿、高温的特点,散布在坡顶的染木种子长期暴露在这样的条件下会加快种子水分的散失,脱水有可能导致种子在萌发条件满足前就失去了活力。与此不同,木奶果种子可以在相对较低的水势条件下萌发,野外散布到5个观测点的种子均有很高的萌发率(普遍达90%以上),野外种子萌发不是该物种分布格局形成的原因,但种子萌发后大量幼苗在不到一年的时间内死亡,沟底略高的幼苗存活率可能是形成木奶果物种分布格局的重要原因。
在西双版纳热带季节性雨林中,木奶果与染木具有相似的分布格局,而且两个物种都具种子脱水敏感的特点,但对于西双版纳季节性的干旱,这两个物种具有不同的适应对策:木奶果在雨季中期果实成熟,种子散布快速萌发,以幼苗的形式度过干热季,尽管大部分幼苗已经在雨季和雾凉季死亡;而染木是在雨季末期果实成熟,种子散布后并不萌发,而是进入3~4个月的休眠期,以种子的形式度过土壤含水量最低的2月、3月。种子在室内自然脱水下的实验表明,相对于木奶果种子,在相类似的温、湿度条件下染木种子具有较高的脱水抗性,这一特点有利于该种子避免因在野外长期存留导致脱水伤害。当染木种子开始萌发的时候,木奶果种子早已经萌发结束,且大部分幼苗已经死亡。从生态位的角度分析,这两个物种在空间上是重叠的,但它们种子萌发和幼苗建立的时间互相错开,因而较好地避免了直接竞争。
[1] FOWLER N. Effects of environmental heterogeneity in space and time on the regulation of populations and communities[J].SymposiumoftheBritishEcologicalSociety, 1988,(1): 249-269.
[2] LEGENDRE P, FORTIN M J. Spatial pattern and ecological analysis[J].Vegetatio, 1989, 80(2): 107-138.
[3] MACARTHUR R, LEVINS R. The limiting similarity, convergence, and divergence of coexisting species[J].TheAmericanNaturalist, 1967, 101(921): 377-385.
[4] WILSON J, GITAY H. Limitations to species coexistence: evidence for competition from field observations, using a patch model[J].JournalofVegetationScience, 1995, 6(3): 369-376.
[5] GRIME J P. Trait convergence and trait divergence in herbaceous plant communities: mechanisms and consequences[J].JournalofVegetationScience, 2006, 17(2): 255-260.
[6] KRAFT N J B, COMWELL W K,etal. Trait evolution, community assembly, and the phylogenetic structure of ecological communities[J].TheAmericanNaturalist, 2007, 170(2): 271-283.
[7] GRUBB P J. The maintenance of species richness in plant communities: the importance of the regeneration niche[J].BiologyReviews, 1977, 52(1): 107-145.
[8] 祝 燕,白 帆,等.北京暖温带次生林种群分布格局与种间空间关联性[J]. 生物多样性, 2011, 19(2): 252-259.
ZHU Y, BAI F,etal. Population distribution patterns and interspecific spatial associations in warm temperate secondary forests, Beijing[J].BiodiversityScience, 2011, 19(2): 252-259.
[9] LAI J S, MI X C, REN H B,etal. Species-habitat associations change in a subtropical forest of China[J].JournalofVegetationScience, 2009, 20(3): 415-423.
[10] YE J, HAO Z Q, XIE P,etal. Habitat associations of saplings and adults in an old-growth temperate forest in the Changbai Mountains, Northeastern China[J].ForestrystudiesinChina, 2011, 13(1): 13-22.
[11] 田 锴, 陈 磊, 米湘成, 等. 亚热带常绿阔叶林木本植物幼苗分布格局及其对生境过滤的响应[J]. 科学通报, 2013, 2013, 58(34): 3 561-3 569.
TIAN K, CHEN L, MI X C,etal. The effect of habitat filtering on tree seedling distribution in a subtropical evergreen broadleaf forest in China[J].Chin.Sci.Bull. (Chin Ver), 2013, 58(34): 3 561-3 569.
[12] 朱 华. 论滇南西双版纳的森林植被分类[J]. 云南植物研究, 2007, 29(4): 377-387.
ZHU H. On the classification of forest vegetation in Xishuangbanna, Southern Yunnan[J].ActaBotanicaYunnanica, 2007, 29(4 ): 377-387.
[13] 刘文杰, 李红梅. 西双版纳旅游气候资源[J]. 自然资源, 1997, 2: 62-66.
LIU W J, LI H M. Tourism climatic resources in Xishuangbanna[J].NaturalResources, 1997, 2: 62-66.
[14] CAO M, ZOU X M, WARREN M,etal. Tropical forests of Xishuangbanna, China[J].Biotropica, 2006,38(3): 306-309.
[15] ZHU H. Forest vegetation of Xishuangbanna, South China[J].ForestryStudiesinChina, 2006, 8(2): 1-27.
[16] 兰国玉, 胡跃华, 曹 敏. 等. 西双版纳热带森林动态监测样地—树种组成与空间分布格局[J]. 植物生态学报, 2008, 32(2): 287-298.
LAN G Y, HU Y H, CAO M,etal. Establishment of Xishuangbanna tropical forest dynamics plot: species compositions and spatial distribution patterns[J].JournalofPlantEcology(Chinese Version), 2008, 32(2): 287-298.
[17] MICHEL B E. Evaluation of water potentials of solutions of polyethylene glycol 8000 both in the absence and presence of other solutes[J].PlantPhysiology, 1983, 72(1): 66-70.
[18] BARBERIS I M, Tanner E V J. Gaps and root trenching increase tree seedling growth in Panamanian semi-evergreen forest[J].Ecology, 2005, 86(3): 667-674.
[19] THIERY J M, D’ HERBES J M, VLAENTIN C. A model simulating the genesis of banded vegetation patterns in Niger[J].JournalofEcology, 1995, 83(3): 497-507.
[20] BUNKER D E, CARSON W P. Drought stress and tropical forest woody seedlings: Effect on community structure and composition[J].JournalofEcology, 2005, 93(4): 794-806.
[21] POORTER L, MARKESTEIJIN L. Seedling traits determine drought tolerance of tropical tree species[J].Biotropica, 2008, 40(3): 321-331.
[22] LEWIS S L, TANNER E V J. Effects of above-and belowground competition on growth and survival of rain forest tree seedlings[J].Ecology, 2000, 81(9): 2 525-2 538.
[23] CAO M, ZHU H, WANG H,etal. Xishuangbanna Tropical Seasonal Rainforest Dynamics Plot: Tree distribution Maps, Diameter Tables and Species Documentation[M]. Kunming: Yunnan Science and Technology Press, 2008: 46, 216.
[24] YU Y, BASKIN J M,etal. Ecology of seed germination of eight non-pioneer tree species from a tropical seasonal rain forest in Southwest China[J].PlantEcology, 2008, 197(1): 1-16.
[25] WEN B, CAI Y F. Seed viability as a function of moisture and temperature in the recalcitrant rainforest speciesBaccaurearamiflora(Euphorbiaceae)[J].AnnalsofForestScience, 2014, 71 (8): 853-861.
[26] CHEN Y J, CAO K F, SCHNITZER S A,etal. Water-use advantage for lianas over trees in tropical seasonal forests[J].NewPhytologist, 2015, 205(1): 128-136.
(编辑:裴阿卫)
Seed Germination and Seedling Survival ofBaccaurearamifloraandSaprosmaternatain Xishuangbanna Tropical Rainforests under Habitat Heterogeneity
LIU Minghang1,2, YE Juan1,2, WEN Bin1*
(1 Center for Integrative Conservation, Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences, Mengla, Yunnan 666303, China; 2 University of Chinese Academy of Sciences, Beijing 100049, China)
Difference at micro-habitats in forest ecosystems may affect seed germination and seedling establishment, lead to various species distribution patterns. In order to explore effects of habitat heterogeneity on plant distribution pattern, we dispersed seeds ofBaccaurearamifloraandSaprosmaternataat five microhabitats which have difference in elevation and topography, in a Xishuangbanna tropical seasonal rainforest. The dynamics of seed germination and seedling survival were monitored. The corresponding land surface (3 cm above the ground) temperature, air humidity (3 cm above the ground) and topsoil (5 cm deep) water content at micro-habitat were also recorded. Meanwhile, seed desiccation under ambient regime and germination under water stress were performed. It was found: (1) at five micro-habitats, soil water content at valley bottom kept the highest all the year round; Air relative humidity was saturated on rainy season and fogy season. Only on dry season there is obvious difference, namely with the increase of altitude, the air humidity dropped; Mountain temperature inversion which last one or two months occured on dry season. (2) In addition,B.ramifloraandS.ternataseeds had different dispersal and germination pattern.S.ternataseed had significant difference in seed germination and seedling survival at different habitats.B.ramiflorahad seed dispersed and quickly germinated on rainy season, and its seed had the ability to germinate under relatively lower water potential, so it spent dry season by seedling; WhileS.ternatadispersed seed in the end of rainy season, which was dormant and relatively desiccation resistant, it pass spent through dry season by seed. In conclusion, topographical condition is a critical factor to effect micro-habitats heterogeneity in Xishuangbanna tropical seasonal rainforest, and the difference in temperature and water regime among these five micro-habitats is most obvious on dry season, which had important effect on seed germination and seedling survival ofB.ramifloraandS.ternata.On the other hand,B.ramifloraandS.ternataseeds have different strategy to cope with drought in Xishuangbanna tropical seasonal rainforest because of their seed traits.
habitat heterogeneity; tropical rainforest; topography; seed germination; seedling survival
1000-4025(2016)08-1654-08
10.7606/j.issn.1000-4025.2016.08.1654
2015-12-20;修改稿收到日期:2016-07-06
国家自然科学基金(31170626)
刘明航(1990-),男,在读硕士研究生,主要从事种子生物学研究。E-mail: liuminghang@xtbg.ac.cn
*通信作者:文 彬,副研究员,硕士生导师,主要从事种子生物学研究。E-mail: wenb@xtbg.org.cn
Q948.114
A
猜你喜欢
——以贵阳花溪公园为例
山地农业生物学报(2022年3期)2022-05-13 09:58:54
云南画报(2021年8期)2021-12-02 02:46:16
现代防御技术(2016年1期)2016-06-01 12:13:27
水利科技与经济(2016年3期)2016-04-09 13:57:20
水利科技与经济(2016年4期)2016-04-09 03:49:18
水利科技与经济(2016年2期)2016-04-09 02:09:13
计算机工程(2015年8期)2015-07-03 12:19:20
云南画报(2015年10期)2015-01-09 04:00:26
云南林业(2014年4期)2014-02-28 21:23:01
计算机工程(2014年6期)2014-02-28 01:26:31
