
辽河源自然保护区油松群落特征及优势种群结构
2016-10-14 05:22:47王泺鑫韩海荣程小琴康峰峰
西北植物学报 2016年8期
王泺鑫,韩海荣,程小琴,康峰峰
(北京林业大学省部共建森林培育与保护教育部重点实验室,北京 100083)
辽河源自然保护区油松群落特征及优势种群结构
王泺鑫,韩海荣*,程小琴,康峰峰
(北京林业大学省部共建森林培育与保护教育部重点实验室,北京 100083)
通过实地踏查,选取辽河源自然保护区内成片的不同年龄的油松天然次生林群落,采用样方法进行群落学特征信息调查,分析研究区的群落区系特征、外貌特征和优势种群结构特征,为油松天然林的保护和抚育管理提供理论依据。结果表明:(1)辽河源自然保护区油松天然次生林群落中共有种子植物41科90属144种,植物种类丰富,具有明显的温带针叶林特征,以地面芽植物为主(60.27%),中叶植物占优势(82.19%);区系组成与温带的成分关系最为密切。(2)优势种油松在研究区构成成片的不同林龄的纯林,幼龄林的群落中小径级(Ⅰ~Ⅲ)个体所占比例较高(75%),随着年龄增加,大径级个体所占比例逐渐增加,成熟林径级呈正态分布,各龄组高度级和冠幅的变化趋势与径级相似。(3)4个龄组幼苗储备都很丰富,均占更新层的58%以上,成熟林中幼苗所占比例达90%,但幼树比例较少,群落的更新状况不佳,但潜在更新能力较强。
油松;群落特征;种群结构
植物群落的基本特征是植物与植物之间、植物与环境之间的相互关系,这些相互关系的可见标志是群落中各种植物在空间、时间上的配置状况,包括群落的区系成分、外貌特征、物种组成、结构特征及多样性特征等[1-3]。优势种作为植物群落中重要的组成部分,对群落物种、群落环境及群落发展有明显的控制作用[4-5]。研究群落特征及优势种群结构可以反映群落现存状况,对了解群落未来发展趋势也有重要意义。
油松属于松科松属植物,是暖温带森林最重要的建群种之一,在辽河源自然保护区有较大面积分布。该区油松林是辽河源自然保护区典型的森林群落类型之一,在海拔950~1 350 m的阳坡、阴坡、半阴坡形成成片的不同林龄的纯林[6]。辽河源自然保护区地理位置特殊,该区的油松天然次生林不仅对辽河流域的水源涵养、水土保持起到重要作用,同时也为京津地区的生态安全提供非常重要的绿色屏障。近些年来,有关学者从景观格局[7]、生物量和分配格局[8-9]、土壤理化性质[10]、土壤微生物特征[11]等方面对辽河源自然保护区内的油松天然次生林进行了研究,但是对该区油松林群落特征及优势种群结构的研究较少,因此,选取辽河源自然保护区内成片的不同年龄的油松天然次生林群落为研究对象,通过对群落区系特征、外貌特征、优势种群结构特征进行研究探讨,为油松天然林的保护和抚育管理提供一定的依据。
1 研究地区与研究方法
1.1 研究区概况
研究区位于河北省平泉县辽河源自然保护区,地理位置为118°22′39″ ~118°37′21″ E,41°01′30″~41°21′15″ N,海拔625~1 738 m,平均海拔1 180 m,属于内蒙古高原与冀北山地的过渡地带,是温带暖温带交错带中的典型地区,该区气候属于半湿润半干旱大陆性季风型的山地气候,四季分明,雨热同季,昼夜温差大,年平均气温7.3 ℃,年平均降雨量550 mm,年平均蒸发量1 800 mm,湿润度0.4,年日照时长2 000~2 900 h,无霜期110~125 d。土壤种类以山地棕壤土和淋溶褐土为主。森林植被主要有油松天然次生林,山杨林(Populusdavidiana),白桦林(Betulaplatyphylla)及其混交林,灌木层主要有土庄绣线菊(Spiraeapubescens)、西伯利亚杏(Prunussibirica)、照山白(Rhododendronmicranthum),草本层有薹草(Carextristachya)、深山堇菜(Violaselkirkii)、大丁草(Leibnitziaanandria)、玉竹(Polygonatumodoratum)和野青茅(Calamagrostisarundinacea)等
1.2 研究方法
1.2.1 样地设置与调查 对辽河源自然保护区进行全面踏查,了解森林分布生长情况,通过生长锥钻取树木木芯样本和轮枝计数法发现该地区有成片的不同林龄的油松纯林。《国家森林资源连续清查技术规定》(2003)中对油松天然林的龄组进行了划分(划分标准:幼龄林年龄为≤30 a、中龄林年龄为31~50 a、近熟林年龄为50~60 a、成熟林年龄为61~80 a)[12]。选择林相整齐、具有代表性的油松天然次生林地段设置面积为20 m×30 m的标准样地,共18块 (幼龄林4块、中龄林6块、近熟林5块、成熟林3块),在每个标准样地的4个角及中心布设5个2 m×2 m灌木样地和25个1 m× m草本样地,记录样地基本信息(表1)和群落学特征信息,包括对样地内胸径≥3 cm的乔木进行每木检尺,记录乔木的名称、株数、树高、胸径、冠幅、等;对胸径3 cm以下的幼苗、幼树,记其株数并测量株高;灌木和草本样地,记录各物种的名称、高度、叶面积等。
1.2.2 植物区系和群落特征划分
植物区系划分按吴征镒[13]中国种子植物属的分布区类型划分,通过野外调查记载和室内鉴定样方标本,统计科属种数及其组成并归类。
植物生活型划分按照Raunkiaer[14]提出的生活型分类系统,按更新芽距土壤表面的位置划分,分为5大类:高位芽植物(Ph)、地上芽植物(Ch)、地面芽植物(H)、地下芽植物(G)和一年生植物(Th),将野外调查记载的维管植物统计分类,计算各类生活型种数和所占比例,绘制生活型谱。叶型划分按照Raunkiaer[14]创立的叶型分类系统进行统计分析。以叶面积(复叶叶面积为所有小叶叶面积之和)大小将植物的叶分为6级,各级间相差9倍,分别为微叶(E),叶面积:0~25 mm2;细叶(D),叶面积:26~225 mm2;小叶(C),叶面积:226~2 025 mm2;中叶(B),叶面积:2 026~18 225 mm2;大叶(A),叶面积:18 226~164 025 mm2;巨叶(S),叶面积:>164 025 mm2。统计野外调查记载的维管植物的叶面积,计算各类种数和所占比例,绘制叶型谱。
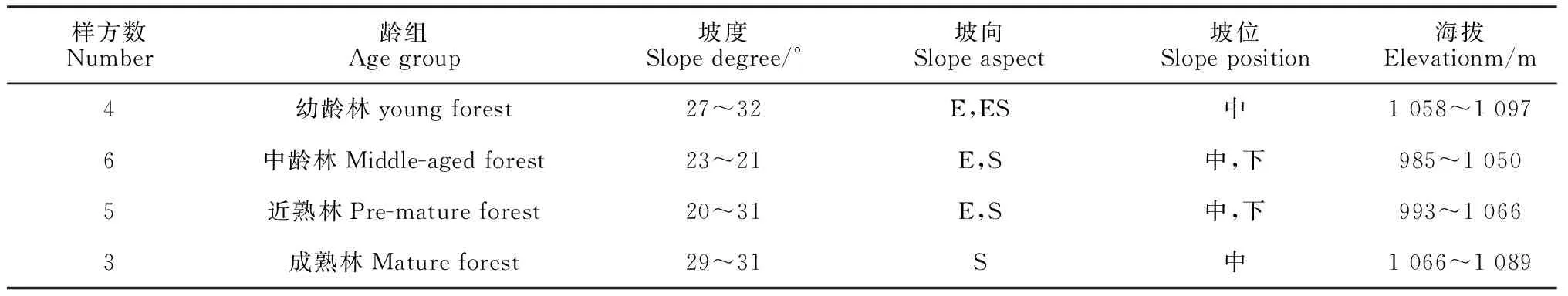
表1 辽河源油松天然次生林样地基本情况
1.2.3 优势种群的径级、高度及冠幅划分
研究区群落中乔木层物种组成单一,优势种和建群种均为油松,仅个别样地伴生有几株山杨、蒙古栎(Quercusmongolia),但其所占比例不足1%,因此按照不同等级利用胸径、高度及冠幅指标对不同年龄段油松种群进行结构划分。胸径(DBH):以4 cm作为级差将胸径划分为Ⅰ~Ⅻ12个级别,其中Ⅰ、3 cm≤DBH<7 cm,Ⅻ、DBH≥47 cm;树高(Height):以3 m作为级差将树高划分为Ⅰ~Ⅸ9个等级,其中Ⅰ、H<3 m,Ⅸ、H≥21 m;冠幅以东西和南北冠幅乘积(Canopy)表示:以5 m2作为级差将冠幅划分为Ⅰ~Ⅻ12个级别,其中Ⅰ、C<5 m2,Ⅻ、C≥55 m2。乔木幼苗幼树以株高(height)作为划分标准分为5级,Ⅰ(幼苗)、h≤0.2 m,Ⅱ(幼树)、0.2
2 结果与分析
2.1 油松天然次生林的群落特点
2.1.1 植物的科、属、种数组成 根据样地调查数据统计(表2),辽河源油松天然次生林群落中共有蕨类植物2科2属2种,银粉背蕨(Aleuritopterisargentea)和中华卷柏(Selaginellasinensis),裸子植物1科1属1种,即油松,种子植物40科89属143种,种子植物中含属较多的科有:菊科(10属)、蔷薇科(6属)、毛茛科(6属)、禾本科(6属)、石竹科(5属)、伞形科(5属),合计占种子植物所有属数的42.22%;含种较多的科为菊科(24种),其次是蔷薇科(10种)、豆科(9种)、禾本科(7种)、毛茛科(7种)、石竹科(6种)、伞形科(6种),以上7科所有植物种占本群落种子植物种数的47.92%。样地内乔木层几乎为油松纯林,偶尔伴生有蒙古栎、山杨等;灌木层的优势种有蔷薇科的土庄绣线菊、西伯利亚杏、杜鹃花科的照山白、迎红杜鹃(Rhododendronmucronulatum)等;草本层主要有菊科、禾本科、石竹科植物。
2.1.2 区系特征 油松天然次生林群落植物组成较为复杂,种子植物(共90属)中以北温带分布的种类占绝对优势(表3),其所占百分比达到47.78%,包括松属(Pinus)、绣线菊属(Spiraea)、孩儿参属(Pseudostellaria)、紫菀属(Aster)、薹草属(Carex)等43个属,其次是世界分布、旧世界温带分布的种类,其所占比例均达到10%以上,其中世界分布有铁线莲属(Clematis)、鼠李属(Rhamnus)、繁缕属(Stellaria)、银莲花属(Anemone)等14个属;旧世界温带分布有13个属,包括:风毛菊属(Saussurea)、白屈菜属(Chelidonium)、败酱属(Patrinia)、沙参属(Adenophora)、菊属(Dendranthema)等。泛热带分布仅有2属:白茅属(Imperata)和狗尾草属(Setaria);热带亚洲至热带非洲分布也仅有香茶菜属(Rabdosia)和水杨梅属(Geum)2个属;属于热带亚洲和热带美洲间断分布、旧世界热带分布、热带亚洲至热带非洲分布的仅有1个属,分别为地榆属(Sanguisorba)、天门冬属(Asparagus)、苦荬菜属(Ixeris)。研究区油松天然次生林群落的植物与温带的成分关系最为密切,显示了该群落的温带性。
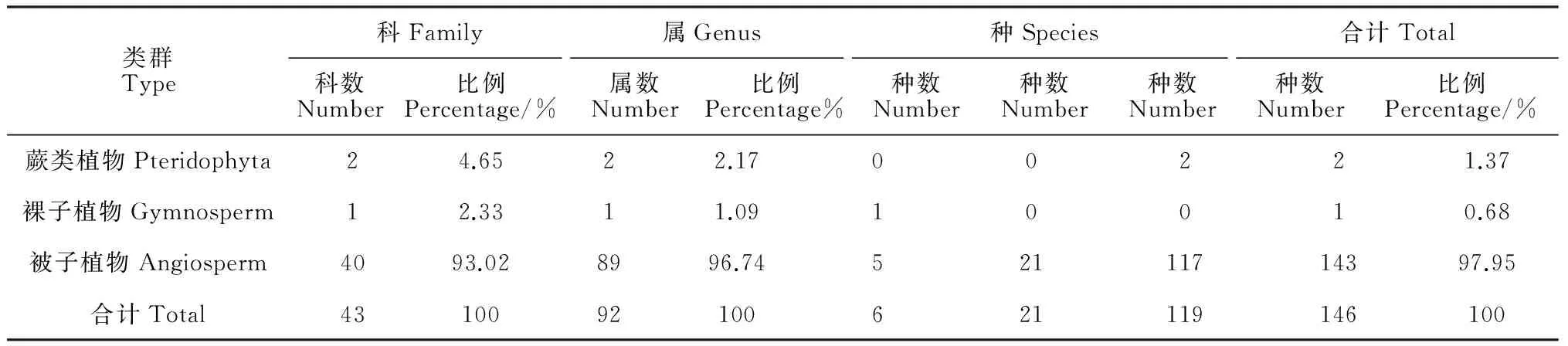
表2 辽河源油松天然次生林群落科、属、种数的统计

表3 辽河源油松天然次生林群落区系组成
2.1.3 群落外貌特征 辽河源地区油松天然次生林群落为常绿针叶林群落,自上而下分为明显的3层乔木层、灌木层和草本层且季相明显,冬季群落进入休眠期,乔木层油松枝叶不再生长,灌木层植物叶片及部分枝稀疏、脱落,草本层植物地上部分枯萎,凋落物层丰厚,植物处于休眠状态;春季气温回升,乔木层油松小枝末端冬芽萌发,灌木层、草本层植物发芽、长叶、开花;夏季植物生长旺盛,群落郁闭度、植被盖度均最大,秋季乔木层油松第二年生球果成熟播种,灌木层植物叶片及部分枝条枯萎,草本层植物地上部分枯黄,向冬季季相过渡。
生活型和叶型是研究群落外貌的重要参数,调查样地中高位芽植物(Ph)有31种(图1),所占比例为21.23%,均为乔木和灌木植物,有油松、核桃楸(Juglansmandshurica)、三裂绣线菊(Spiraeatrilobata)、东陵八仙花(Hydrangeabretschneideri)、照山白等;地上芽植物(Ch)9种,所占比例为6.16%,包括矮小半灌木、匍匐植物和垫状植物;地面芽植物(H)88种,所占比例为60.27%,包括堇菜属植物、地榆(Sanguisorbaofficinalis)等;地下芽植物(G)6种,占总数的4.11%,主要种类有玉竹、角盘兰(Herminiummonorchis)等;一年生植物(Th)12种,占总数的8.22%。按照Raunkiaer的生活型谱,在研究区的油松天然次生林群落中地面芽植物占优势,高位芽植物次之,地上芽植物、地下芽植物和一年生植物较少。表明研究区生长季温度适宜,降水充沛,冬季虽然寒冷,但雪层能保护大部分植物。
调查样地内植物属于大叶的只有核桃楸和糙苏(Phlomisumbrosa)2种植物,属于中叶的植物种类最多,共有120种,占调查样地植物种数的82.19%,其次是小叶,有15种,主要有地榆、水杨梅(Geumaleppicum)、龙芽草(Agrimoniapilosa)等,细叶和微叶所占比例也很小,主要植物有细叶苔草(Carexrigescens)、猪毛菜(Salsolacollina)、蓬子菜(Galiumverum)、附地菜(Trigonotispeduncularis)等。叶子的大小形态与气候有密切的关系,中叶植物占明显优势反映出该区夏季比较温和湿润的特点。
2.2 油松天然次生林种群径级、树高、冠幅结构特征
对研究样地内油松的生长状况(表4)进行调查发现幼龄林到中龄林油松生长迅速,胸径、树高、冠幅都增长较快,随着林龄增加,胸径仍逐渐增大而树高和冠幅生长则较缓慢。自然生长状态下,幼龄林密度和郁闭度都最大,随着油松林龄的增加,种群的自疏作用明显,样地油松株数迅速减少,郁闭度降低,到成熟林油松密度维持在一定的水平上。胸高断面积是计算蓄积量的重要指标,其值同时也可以反映样地的蓄积量,随着林龄增加,样地蓄积量逐渐增大,近熟林到成熟林蓄积量增加最快。植物体的各器官间在生长上表现出相互依赖和相互制约的相关性,植物的营养生长期,随着年龄的增长,树高、胸径、冠幅都表现出相应的特征。
2.2.1 油松种群径级结构 由图2可知从4个龄组油松天然次生林群落乔木胸径分布的统计结果来看,幼龄林(a)的径级结构近似于倒J型分布,随着胸径的增加,个体株数逐渐减少,小径级林木(胸径属于Ⅰ、Ⅱ、Ⅲ径级)占75%,其中属于Ⅱ径级(胸径为7 cm~11 cm)的个体株数最多,占40%左右,大径级林木(胸径≥30 cm)只占0.5%,个体数仅有7株。中龄林(b)中中径级(胸径属于Ⅳ、Ⅴ、Ⅵ径级)个体数显著增加,群落内小径级林木和中径级林木占该群落80%以上,其中小径级林木占33%以上,中径级林木大约占47%,同时也有一定数量的大树分布,表明种群表现出稳定增长趋势。近熟林(c)的径级结构为峰型(中径级个体储备型),个体主要集中在中径级林木,约占60%,其中属于Ⅵ径级(27~31 cm)个体数最多,大约占30%,小径级林木与大径级林木则相对较少,分别约占20%,种群处于稳定发育阶段。成熟林(d)的胸径分布比较均匀,小、中、大径级林木所占比例分别约为29%、43%、28%,近似正太分布,种群仍表现增长趋势。
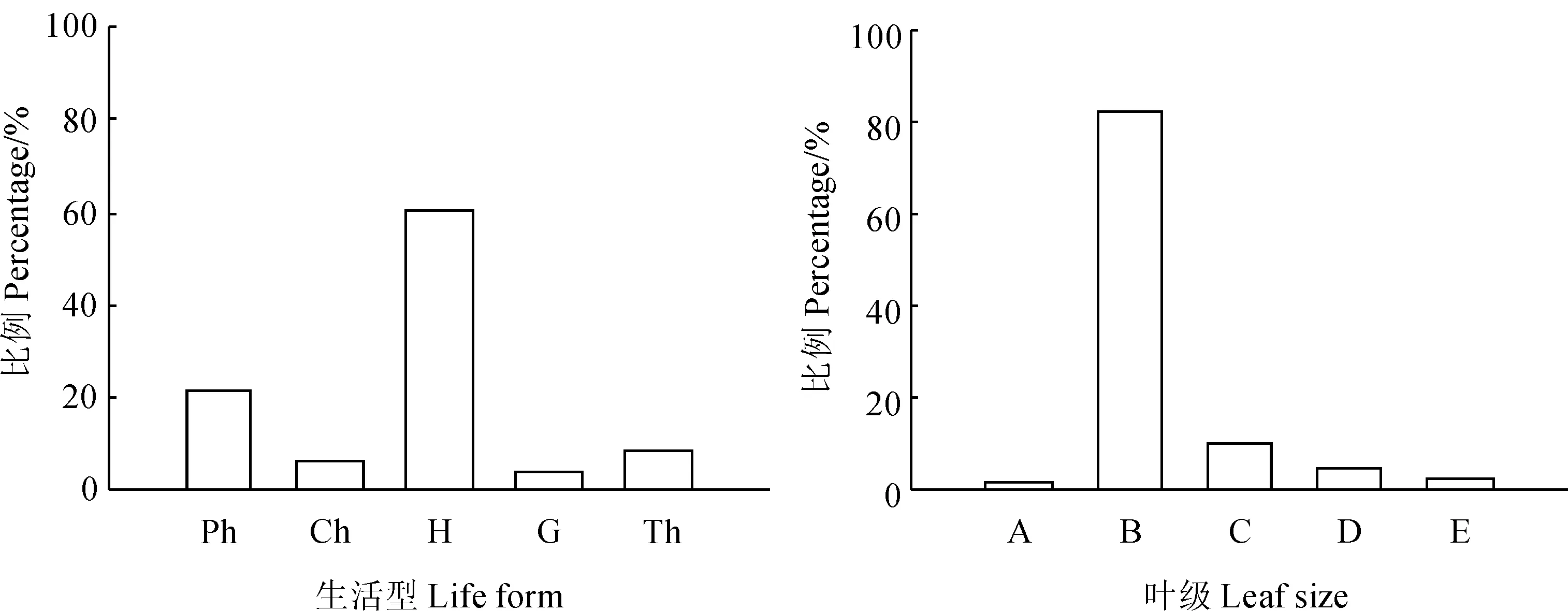
Ph.高位芽植物;Ch.地上芽植物;H.地面芽植物;G.地下芽植物;Th.一年生植物;A.大叶(18 226~164 025 mm2);B.中叶(2 026~18 225 mm2);C.小叶(226~2 025 mm2);D.细叶(26~225 mm2);E.微叶(0~25 mm2)图1 辽河源油松天然次生林群落生活型谱和叶级谱Ph. Phanerophyte;Ch. Chamaephytes;H. Hemicryptophyte;G. Geophyte;Th. Therophyte;A. Large Leaf (18 226~164 025 mm2);B. Middle Leaf(2 026~18 225 mm2);C. Leaflet(226~2 025 mm2);D. Fine Leaf (26~225 mm2);E. Leptophyll(0~25 mm2)Fig. 1 Species life form spectrum and leaf size classification in the P. tabuliformis nature forest of Liaoheyuan
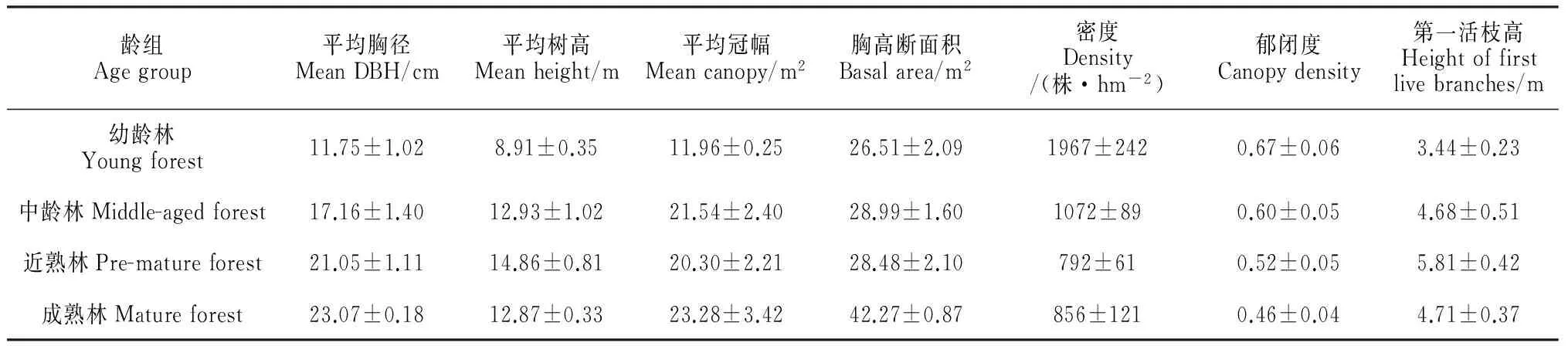
龄组Agegroup平均胸径MeanDBH/cm平均树高Meanheight/m平均冠幅Meancanopy/m2胸高断面积Basalarea/m2密度Density/(株·hm-2)郁闭度Canopydensity第一活枝高Heightoffirstlivebranches/m幼龄林Youngforest11.75±1.028.91±0.3511.96±0.2526.51±2.091967±2420.67±0.063.44±0.23中龄林Middle-agedforest17.16±1.4012.93±1.0221.54±2.4028.99±1.601072±890.60±0.054.68±0.51近熟林Pre-matureforest21.05±1.1114.86±0.8120.30±2.2128.48±2.10792±610.52±0.055.81±0.42成熟林Matureforest23.07±0.1812.87±0.3323.28±3.4242.27±0.87856±1210.46±0.044.71±0.37
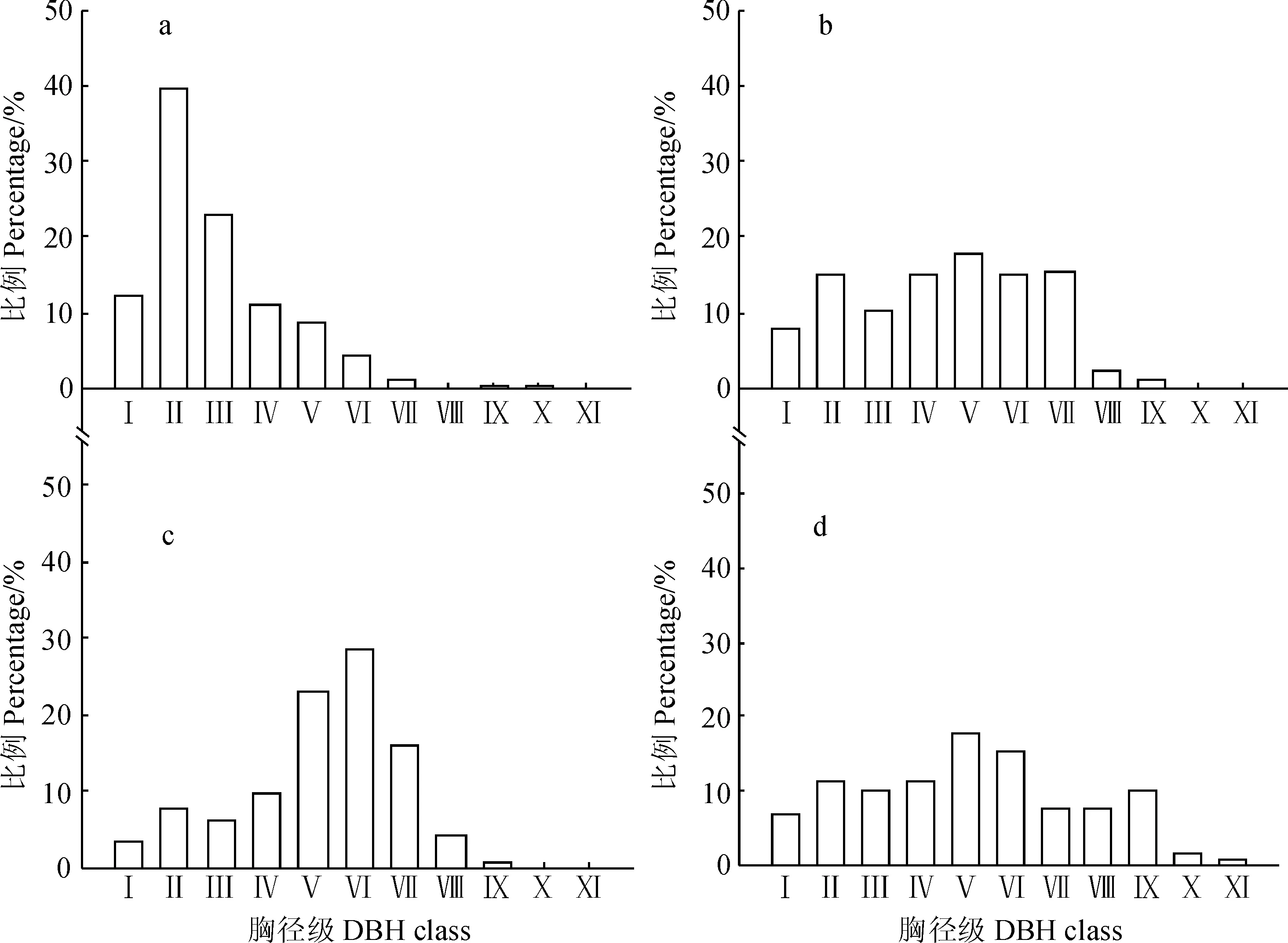
a .幼龄林;b. 中龄林;c. 近熟林;d. 成熟林;Ⅰ. 3 cm≤胸径<7 cm;Ⅱ. 7 cm≤胸径<11 cm;Ⅲ~Ⅺ. 11 cm≤胸径<47 cm,每级相差4 cm;Ⅻ. 胸径≥47 cm图2 不同龄组的油松径级分布a. Young forests;b. Middle-aged forests;c. Immature forests;d. Mature forests;Ⅰ. 3 cm≤DBH<7 cm;Ⅱ. 7 cm≤DBH<11 cm;Ⅲ-Ⅺ. 11 cm≤DBH<47 cm,4 cm increased per level;Ⅻ. DBH≥47 cm.Fig. 2 Diameter class distribution in the each age group of P. tabuliformis nature forests
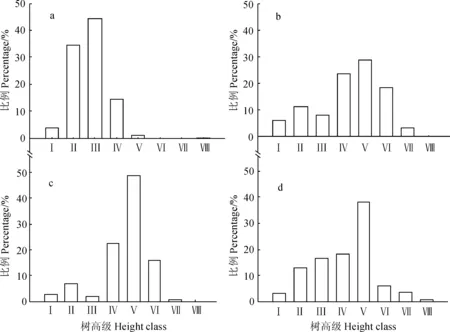
a .幼龄林;b. 中龄林;c. 近熟林;d. 成熟林;Ⅰ. 树高<3 m; Ⅱ.3 m≤树高<6 m;Ⅲ~Ⅷ. 6 m≤树高<21 m,每级相差3 m;Ⅸ. 树高≥21 m图3 不同龄组的油松高度级分布a. Young forests;b. Middle-aged forests;c. Immature forests;d. Mature forests;Ⅰ.H<3 m;Ⅱ.3 m≤H<6 m;Ⅲ-Ⅷ. 6 m≤H<21 m,3 m increased per level;Ⅸ.H≥21 mFig.3 Height class distribution in the each age group of P. tabuliformis nature forests
2.2.2 油松种群高度结构 由4个龄组油松天然次生林群落乔木树高分布的统计结果(图3)可以看出,幼龄林(a)的树高分布集中在Ⅱ、Ⅲ高度级(3~9 m)上,占80%左右,很少有树高在12 m以上的植株,林木个体比较矮小;中龄林(b)、近熟林(c)的树高分布均集中Ⅴ高度级(12~15 m),分别占各群落的28.74%、48.95%,表明从幼龄林到中龄林是油松树高增长最快的时期,中龄林之后,大部分油松个体的树高维持在平均树高左右,树高生长缓慢,成熟林(d)的树高仍集中在Ⅴ高度级,但是树高在20 m以上的个体数在4个龄组中最多。
2.2.3 油松种群冠幅结构 冠幅的大小决定树木光合作用的能力,显示植株占有空间的能力,也反映了树木的生产力,由图4可以看出,油松幼龄林(a)的冠幅分布也近似呈现倒J型,集中在Ⅱ级(5~10 m2)上,占36.39%,中龄林(b)、近熟林(c)的个体在Ⅱ~Ⅴ级上分布较均匀,成熟林(d)的冠幅分布均集中Ⅳ级(15~20 m2),和树高生长相似,幼龄林到中龄林是油松的冠幅增大最快的时期,中龄林之后,冠幅生长减缓,成熟林冠幅分布趋于正太分布,较大冠幅的植株所占比例显著增加,大冠幅个体在群落中所占比例与其他龄组相比也是最多。
对4个龄组油松天然次生林的冠幅和胸径及树高进行Pearson相关性检验,结果显示幼龄林的冠幅和胸径及树高的相关性都不显著,中龄林、近熟林、成熟林的冠幅和树高没有显著的相关关系,而和胸径存在显著的相关关系,通过线性方程进行拟合,中龄林的冠幅胸径拟合方程为y=-2.753+1.329x(P<0.05,R=0.526),近熟林的冠幅胸径拟合方程为y=-4.794+1.250x(P<0.05,R=0.463)成熟林的冠幅胸径拟合方程为y=-1.753+1.087x(P<0.05,R=0.422)。以上数据说明:中龄林之后,油松冠幅与胸径的相关性比与树高的相关性更密切,这种相关性随着年龄的增加而降低。
2.3 油松种群更新
森林更新是一个重要的生态学过程。研究区内油松天然次生林所受人为干扰较少,多为天然下种。从油松幼苗幼树高度级结构(图5)可以看出,4个龄组油松天然次生林群落中优势种油松的种子萌发率均较高,林内实生幼苗较多,幼龄林、中龄林中株高≤0.2 m的幼苗的数量大约为3 000株·hm-2,近熟林和成熟林的林下幼苗数量则能达到6 000株·hm-2,4个龄组幼苗的数量在更新层所占比例均达到58%以上,成熟林则高达93%。幼龄林中幼树的生长状况良好,随着高度级增大,个体株数也逐渐增加;中龄林幼树和近熟林林下的幼树在2 m(4个龄组灌木层植被高度范围为0.2~2.5 m)以上的个体数骤减,可能原因是中龄林和近熟林的林下灌木层竞争激烈,不利于幼树生长;成熟林中高度>0.2 m(4个龄组草本层植被高度范围为0.01~0.2 m)的油松幼树个体很少,仅有几株,大部分个体分布在草本层,更新状况不佳。油松天然次生林更新层幼苗、幼树的分布反映出林下灌木层和草本层是影响幼苗更新的一个重要的生物因素。幼龄林尚未形成稳定的林下环境,油松更新状况较好,随着油松林龄的增加,林下环境逐渐稳定,灌木层植物对油松幼树的生长产生影响,油松幼树很难长成大树,成熟林中灌木层和草本层植物分布均比较稳定,油松幼苗在草本层竞争中优势不明显,长成幼树的个体数很少。
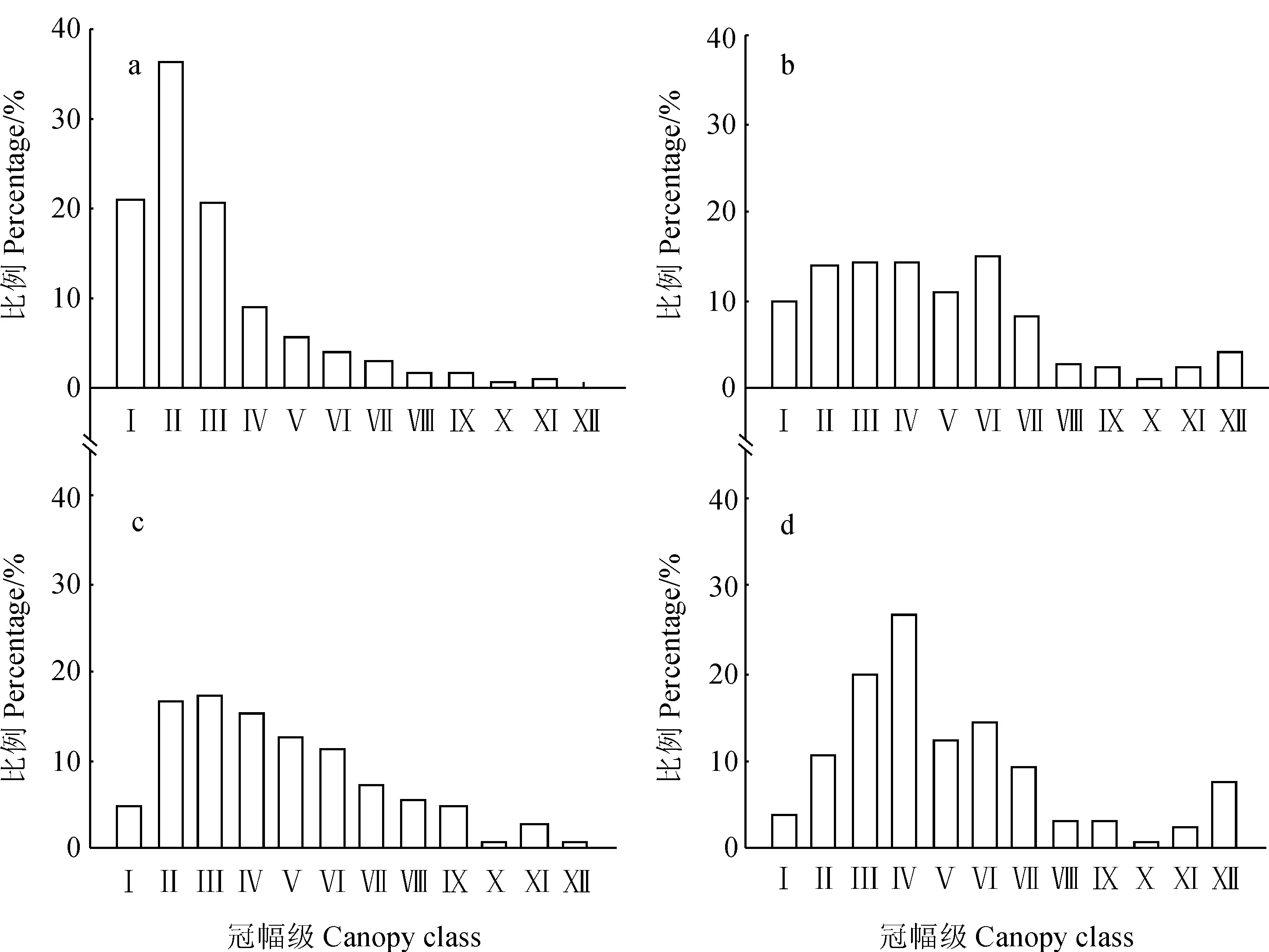
a.幼龄林;b. 中龄林;c. 近熟林;d. 成熟林;Ⅰ. 冠幅<5 m2;Ⅱ.5 m2≤冠幅<10 m2;Ⅲ~Ⅺ. 10 m2≤冠幅<55 m2,每级相差5 m2;Ⅻ. 冠幅≥55 m2图4 不同龄组的油松冠幅分布a. Young forests;b. Middle-aged forests;c. Immature forests;d. Mature forests;Ⅰ.C<5 m2;Ⅱ.5 m2≤C<10 m2;Ⅲ-Ⅺ. 10 m2≤C<55 m2,5 m2 increased per level;Ⅻ.C≥55 m2Fig. 4 Canopy class distribution in the each age group of P. tabuliformis nature forests
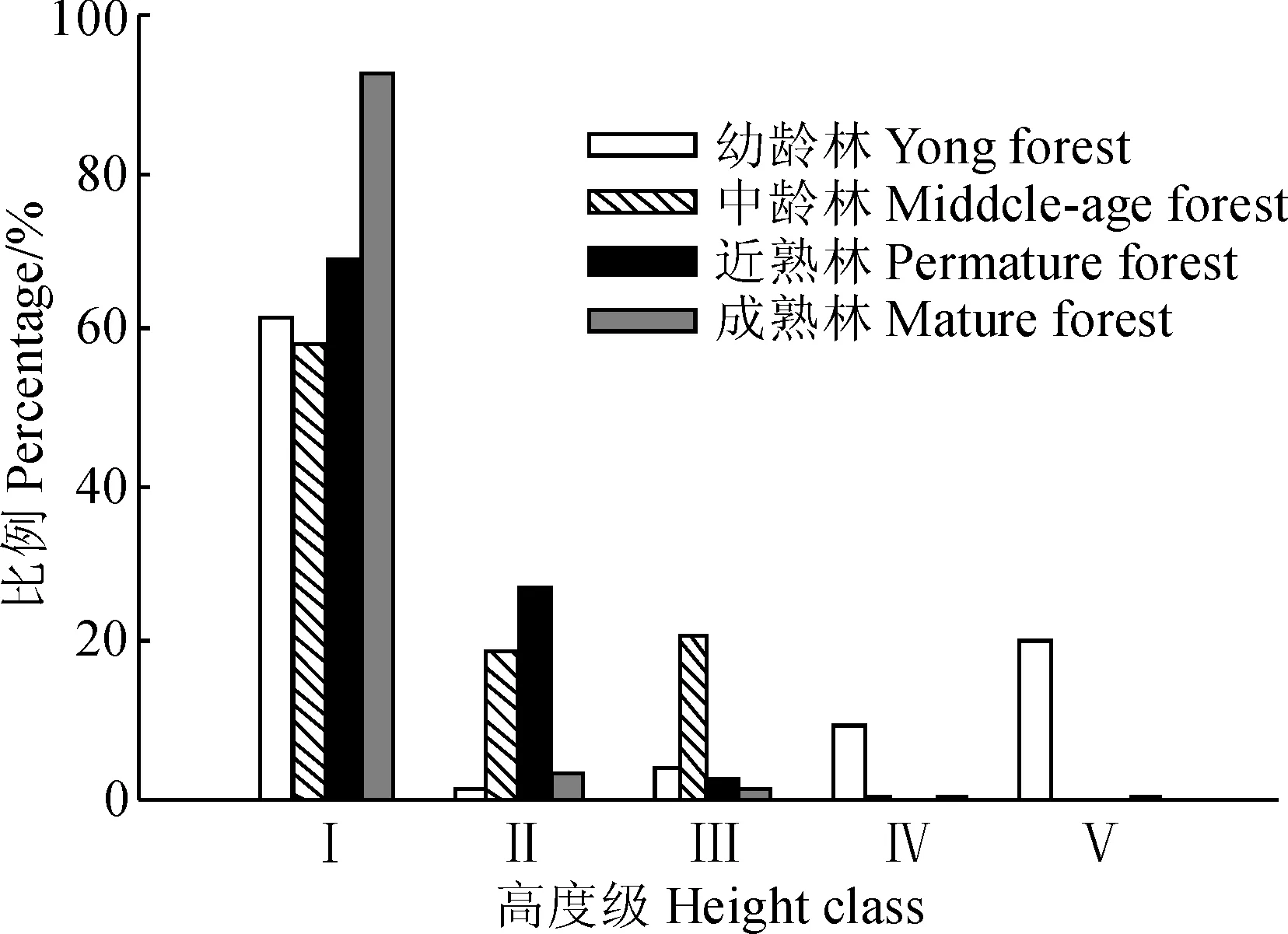
Ⅰ.幼苗(H≤0.2 m);Ⅱ.幼树(0.2
3 讨 论
水热条件的不同导致各地区植物分布的差异[15],这些差异通过植物区系、群落外貌,如物种组成、生活型组成、植物叶的特征反映出来[3]。辽河源地处内蒙古高原和冀北山地的过渡地带,是典型的蒙古植物区系和华北植物区系的交汇地带,吴征镒曾把该地区划为泛北极植物区的中国-日本森林亚区,华北地区,华北平原、山地亚地区[16]。调查发现相比于其他油松天然林[17],研究区群落中植物种类丰富,裸子、被子、蕨类植物均有分布,并且以温带性质成分的种类占绝对优势,热带性质成分的种类很少;生活型上,一般而言,热带雨林中高位芽植物占绝对优势;而在四季分明的温带,地面芽植物是对这种气候条件适应最成功的生活型[18],研究区植物的生活型谱中地面芽植物所占比例达50%以上,处于绝对优势地位,与温带地区群落的生活型谱特征相符合,也反映出该地区冬季虽然寒冷但是降雪丰沛的特点;相关对叶级分布的研究表明,大叶片植物经常出现在热带气候温暖潮湿的地区,而小叶片植物则常出现在气候十分干燥或寒冷地区,从湿润到干燥,从温暖到寒冷,植物叶面积通常表现出由大到小逐渐缩小的现象[19],调查样地中以中叶植物为主,大叶和微叶植物所占比例很少,反映出该区生长季温热、降雨适宜的特点。区系组成、生活型组成及植物叶的大小这些特征综合显示了辽河源自然保护区的暖温带性,同时也反映出该区夏季温和湿润,冬季寒冷多雪的气候特点。
优势种是群落的重要组成部分,其结构不仅对群落结构有直接影响,而且能客观地体现群落的发展趋势[5]。通过划分林龄研究优势种油松的种群结构,可以深入了解辽河源自然保护区各年龄阶段油松的生长状况,结果表明:4个龄组油松天然林的胸径分布、树高分布和冠幅分布符合油松的生长规律,随着油松年龄的增大,大部分油松个体的胸径、树高、冠幅都增大,但各林龄油松的生长发育也存在差异。幼龄林密度最大,为1967株·hm-2,油松个体的胸径、树高、树冠较小,大部分集中在Ⅰ、Ⅱ、Ⅲ上,处在快速生长期,种群呈现增长趋势。牛丽丽认为油松天然林群落中优势种个体集中在同一林龄,会使该种群的竞争加剧,随着种群个体的生长,大部分小树将在竞争中被淘汰[20]。本研究中,中龄林的密度显著小于幼龄林,且大树所占的比例也显著增加,与其研究一致。中龄林和近熟林均为中间个体储备型,但和中龄林相比,近熟林的胸径和树高分布的峰值向更大的径级和高度级推移,胸径分布的峰值出现在Ⅵ(23~27 cm)径阶上,树高分布的峰值出现在Ⅴ(12~15 cm)高度级上,而冠幅分布的变化不明显。成熟林中,胸径和冠幅的分布趋于正太分布,表明成熟林油松种群比较稳定,油松属于增长型种群,林龄较长,处于旺盛发育的个体可能维持50-100年[21],因此可以推测在未来一段时间内,油松仍会是该群落的优势种;胸径和冠幅较大的油松个体所占比例显著增加,表明近熟林到成熟林是油松蓄积量增加最快的阶段。在经营管理中,可以对幼龄林进行适当间伐,促进种群发展;中龄林、近熟林和成熟林采取适当抚育管理,使种群更加稳定。
优势种幼苗在群落中的定居、生长与该物种天然更新密切相关,同时决定了群落的演替发展方向[22-24]。油松天然林主要通过种子繁殖,因此种子和幼苗的存活、生长状况成为更新的关键。辽河源自然保护区油松天然次林各龄组内实生幼苗数量很大,可以推断该区成年个体的繁殖能力和种子的萌发能力均较高,但只有幼龄林的更新状况较好,中龄林、近熟林及成熟林中只有少数幼苗可以成长为成年个体,尤其是成熟林幼苗和幼树所占比例差异很大,更新状况不佳。韩文娟等对油松人工林幼苗更新的研究表明30年生油松人工林中影响幼苗萌发、生长的因素为林内郁闭度大,光照不足和乔木间较强的竞争力[25],田丽等在研究黄土高原马栏山林区油松及辽东栎天然林幼苗更新中得出:灌草层通过直接的种间资源竞争及间接的改变微环境对优势种的更新产生影响,成为林下优势种幼苗长成幼树、大树的生态学滤筛[26]。本研究中对比各龄组油松幼苗幼树比例说明研究区天然林中林下灌草层构成的微环境是乔木更新的主要影响因素。Denslow等的研究指出,草本层会影响乔木幼苗的生长和存活,是森林更新过程一道潜在障碍,但是草本层的作用是非特异性的,可以调节幼苗库的密度[27]。因此辽河源自然保护区油松天然次生林的潜在更新能力较强,对该区植被干扰后恢复具有重要意义。
[1] 李俊清. 森林生态学[M]. 北京:高等教育出版社. 2010: 211-272.
[2] SCHWILK D W, ACKERLY D. Limiting similarity and functional diversity along environmental gradients[J].EcologyLetters, 2005, 8(3): 272-281.
[3] 刘万德, 臧润国, 丁易. 海南岛霸王岭两种典型热带季雨林群落特征[J]. 生态学报, 2009, 29(7): 3 465-3 476.
LIU W D, ZANG R G, DING Y. Community features of two types of typical tropical monsoon forests in Bawangling Nature Reserve, Hainan Island[J].ActaEcologicaSinica, 2009, 29(7): 3 465-3 476.
[4] 田 丽, 王孝安, 郭 华, 等. 红黄土高原马栏林区优势种幼苗与其种群径级结构的演替研究[J]. 西北植物学报, 2006, 26(12): 2 560-2 566.
TIAN L, WANG X A, GUO H,etal. Succession of the seedlings of dominant species and diameter-class structures of their populations in Malan Forest Region of the Loess Plateau[J].ActaBotanicaBoreali-OccidentaliaSinica, 2006, 26(12): 2 560-2 566.
[5] 达良俊, 杨永川, 宋永昌. 浙江天童国家森林公园常绿阔叶林主要组成种的种群结构及更新类型[J]. 植物生态学报. 2004, 28(3): 376-384.
DA L J, YANG Y C, SONG Y C. Population structure and regeneration types of dominant species in an evergreen broadleaved forest in Tiantong National Forest Park, Zhejiang Province, Eastern China[J].ActaPhytoecologicaSinica, 2004, 28(3): 376-384.
[6] 王泺鑫, 韩海荣, 程小琴, 等. 辽河源自然保护区油松群落结构及物种多样性[J]. 生态学杂志, 2016, 35(2): 314-320.
WANG L X,HAN H R,CHENG X Q,etal. Community structure and species diversity ofPinustabuliformisforest in the Liaoheyuan Nature Reserve[J].ChineseJournalofEcology, 2016, 35(2): 314-320.
[7] 于晓文, 康峰峰, 韩海荣, 等. 冀北辽河源森林景观格局特征及破碎化研究[J]. 安全与环境学报, 2014, 14(2): 278-282.
YU X W, KANG F F, HAN H R,etal. Analysis of landscape patterns and forest fragmentation trend in Liaohe Source Area, Hebei[J].JournalofSafetyandEnvironment, 2014, 14(2): 278-282.
[8] 赵金龙, 王泺鑫, 韩海荣, 等. 辽河源不同龄组油松天然次生林生物量及空间分配特征[J]. 生态学报, 2014, 34(23): 7 026-7 037.
ZHAO J L, WANG L X, HAN H R,etal. Biomass and spatial distribution characteristics ofPinustabulaeformisnatural secondary forest at different age groups in the Liaoheyuan Nature Reserve, Hebei Province[J].ActaEcologicaSinica, 2014, 34(23): 7 026-7 037.
[9] ZHAO J L, KANG F F, WANG L X,etal. Patterns of biomass and carbon distribution across a chronosequence of Chinese Pine (Pinustabulaeformis) forests[J].PlosOne, 2014, 9(4): e94966
[10] 赵伟红, 康峰峰, 韩海荣, 等. 冀北辽河源地区不同林龄油松天然次生林土壤理化特征的研究[J]. 西北林学院学报, 2014,29(3): 1-8.
ZHAO W H, KANG F F, HAN H R,etal. Physicochemical properties of the soils ofPinustabulaeformisnatural secondary stands with different ages in Liaoheyuan area of northern Hebei[J].JournalofNorthwestForestryUniversity, 2014, 29(3):1-8.
[11] 立天宇, 康峰峰, 韩海荣, 等. 冀北辽河源油松天然林土壤微生物碳代谢特征研究[J]. 土壤, 2015,47(3): 550-557.
LI T Y, KANG F F, HAN H R,etal. Charateristics of microbial carbolic metabolism in soils ofPinustabulaeformisin Liaohe river of northern Hebei[J].Soil, 2015,47(3):550-557.
[12] 国家林业局. 国家森林资源连续清查技术规定[M]. 北京: 国家林业局. 2003.
[13] 吴征镒. 中国种子植物区系地理[M]. 北京:科学出版社. 2010: 52-291.
[14] RAUNKIAER C. The life forms of plants and statistical plant geography[M]. Oxford: Oxford University Press. 1934:23-210.
[15] 古丽努尔·沙比尔哈孜, 潘伯荣, 段士民. 塔里木盆地塔里木沙拐枣群落特征[J]. 生态学报, 2012, 32(10): 3 288-3 295
GULNUR S, PAN B R, DUAN S M. The community characteristics ofCalligonumroborowskiiA. Los in Tarim Basin[J].ActaEcologicaSinica, 2012. 32(10): 3 288-3 295.
[16] 吴征镒. 论中国植物区系的分区问题[J]. 云南植物研究, 1979,1(1): 1-20
WU Z Y. Distribution of China flora partition[J].ActaBotanicaYunnanica, 1979,1(1): 1-20.
[17] 张希彪, 郭小强, 上官周平, 等. 黄土丘陵子午岭油松天然林群落特征研究[J]. 植物研究, 2006,26(2): 106-175.
ZHANG X B, GUO X Q, SHANGGUAN Z P,etal. Community characteristics of naturalPinustabulaeformisforest in Hilly Loess Regions[J].BulletinBotanicalResearch. 2006,26(2): 106-175.
[18] 高贤明, 陈灵芝. 植物生活型分类系统的修订及中国暖温带森林植物生活型谱分析[J]. 植物学报, 1998, 40(6): 553-559.
GAO X M, CHEN L Z. The revision of plant life-form system and an analysis of the life-form spectrum of forest plants in the Warm Temperate Zone of China[J].ActaBotanicaSinica, 1998, 40(6): 553-559.
[19] 于顺利, 马克平, 陈灵芝. 蒙古栎群落叶型的分析[J]. 应用生态学报, 2003, 14(1): 151-153.
YU S L, MA K P, CHEN L Z. Analysis on leaf forms inQuercusmongolicacommunity[J].ChineseJournalofAppliedEcology, 2003, 14(1): 151- 153.
[20] 牛丽丽, 余新晓, 岳永杰. 北京松山自然保护区天然油松林不同龄级立木的空间点格局[J]. 应用生态学报, 2008, 19(7): 1 414-1 418.
NIU L L, YU X X, YU Y J. Spatial patterns of different age-class individuals inPinustabulaeformisforest in Songshan Nature Reserve of Beijing, China[J].ChineseJournalofAppliedEcology, 2008, 19(7): 1 414-1 418.
[21] 张 赟, 赵亚洲, 张春雨, 等. 北京松山油松种群结构及空间分布格局[J]. 应用与环境生物学报, 2009, 15(2): 175-179.
ZHANG Y, ZHAO Y Z, ZHANG C Y. Structure and spatial distribution ofPinustabulaeformispopulation in the Songshan Nature Reserve, Beijing, China[J].ChineseJournalofAppliedandEnvironmentalBiology, 2009, 15(2): 175-179.
[22] FYLLASy N M, POLITI P I, GALANIDIS A,etal. Simulating regeneration and vegetation dynamics in Mediterranean coniferous forests[J].EcologicalModelling, 2010, 221:1 494-1 504
[23] NABE-NIELSE J, SEVERICHE W, FREDERICKSEN T, et al. Timber tree regeneration along abandoned logging roads in a tropical Bolivian forest[J].NewForests, 2007, 34(1): 31-40.
[24] 康 冰, 刘世荣, 王得祥, 等. 秦岭山地典型次生林木本植物幼苗更新特征[J]. 应用生态学报, 2011, 22(12): 3 123-3 130.
KANG B, LIU S R, WANG D X,etal. Regeneration characteristics of woody plant seedlings in typical secondary forests in Qinling Mountains[J].ChineseJournalofAppliedEcology, 2011, 22(12): 3 123-3 130.
[25] 韩文娟, 张文辉, 何景峰, 等. 不同坡向油松人工林建群种种群结构及群落特征分析[J]. 西北农林科技大学学报(自然科学版), 2012, 40(3): 47-55.
HAN W J, ZHANG W H, HE J F,etal. Analysis of population structure and plantation community characteristics ofPinustabulaeformisplantations in Loess Plateau in different slope[J].JournalofNorthwestAgricultureandForestryUniversity(Natural Science Edition), 2012, 40(3): 47-55.
[26] PREVOSTO B, RIPERT C. Regeneration ofPinushalepensisstands after partial cutting in southern France: Impacts of different ground vegetation, soil and logging slash treatments[J].ForestEcologyandManagement, 2008, 256(12): 2 058-2 064.
[27] DENSLOW J S, UOWOLO A L, HUGHES R F. Limitations to seedling establishment in a mesic Hawaiian forest[J].Oecologia, 2006,148: 118-128.
(编辑:潘新社)
Community Characteristics and Population Structure of Dominant Species ofPinustabuliformisForest in the Liaoheyuan Nature Reserve
WANG Luoxin, HAN Hairong*, CHENG Xiaoqin, KANG Fengfeng
(Key Laboratory of Forest Cultivation and Conservation of the Province and the Ministry of Education, Beijing Forestry University, Beijing 100083, China)
Researches were carried out in an age sequence ofPinustabuliformisnatural secondary forest through field surveys on the spots in the Liaoheyuan Nature Reserve. The community characteristic information was studied using quadrat investigation, and by analysing the floristic feature, the physiognomy characteristics, and the population structure of dominant species ofP.tabuliformisnatural forest. The study was proposed to provide theoretical basis for the protection and management of naturalP.tabuliformisforest. Results showed that: (1) the floristics was abundant inP.tabuliformisnatural forest community in the Liaoheyuan Nature Reserve, which was composed of 144 species wild seed plants belonging to 90 genera and 41 families. The appearance of community showed obvious temperate coniferous forest, which characterized by hemicryptophyte plants(60.27%) with mesophyll leaf(82.19%). North-temperate element were plentiful, indicating that the community was closely related to the temperate zone. (2) Outstanding dominant species in the community wasP.tabuliformiswhich formed pure forest of different ages. Small-diameter(Ⅰ—Ⅲ) trees were more frequent(75%) than other diameter classes in young forest. The ratio of big-diameter trees increased with the growth ofP.tabuliformis, and the diameter classes had normal distributions in mature forest. Distribution of height classes and canopy classes in each age group were similar as diameter classes. (3) Seedlings in each age group were abundant, especially in mature forest, accounted for 90%, which reflected that potential regeneration ability in the community was strong, while saplings was small, meaning community regenerative condition was not so well.
Pinustabuliformis; community composition; population structure
1000-4025(2016)08-1687-10
10.7606/j.issn.1000-4025.2016.08.1687
2016-06-18;修改稿收到日期:2016-07-23
林业公益性行业科研专项(201204101)
王泺鑫(1992-),女,在读硕士研究生,主要从事森林生态学研究。E-mail: 1587069562@qq.com
*通信作者:韩海荣,教授,博士导师,主要从事生态学研究和规划研究。E-mail: hanhr@ bjfu.edu.cn
Q948.15+7; S718.54
A
猜你喜欢
乡村科技(2023年20期)2024-01-08 07:20:00
广西林业科学(2022年4期)2022-11-07 01:50:34
林业调查规划(2022年3期)2022-06-13 09:31:04
广西林业科学(2021年6期)2022-01-21 07:39:10
森林工程(2018年5期)2018-05-14 13:54:30
现代农业研究(2017年11期)2018-01-12 21:03:03
辽宁林业科技(2017年4期)2017-06-22 10:23:36
辽宁林业科技(2017年4期)2017-06-22 10:23:36
现代园艺(2017年20期)2017-02-03 18:49:29
广西林业科学(2016年2期)2016-03-20 05:53:41
