
利用RAPD、ISSR分子标记分析野牡丹属亲缘关系
2016-05-30 15:30:31陈振东郑涛林秀香林艺华郑少缘苏金强
热带作物学报 2016年9期
陈振东 郑涛 林秀香 林艺华 郑少缘 苏金强
摘 要 采用ISSR和RAPD分子标记技术对9个种44份野牡丹属种质资源进行了亲缘关系分析。结果表明:11条ISSR引物共扩增出91条谱带,其中多态性条带90条,多态性条带的比率为98.9%;16 条RAPD引物共扩增出113条谱带,其中101条多态性条带,多态性比率为89.38%。2种分子标记相比较,ISSR分子标记检测出的多态性比率更高。44份野牡丹属种质明显聚为2组,种质资源遗传相似系数在0.61~0.88,平均相似系数0.75。基于不同引物的条带组合构建了供试的44份野牡丹属种质资源的ISSR和RAPD指纹图谱,采用这2种指纹图谱可对供试的所有野牡丹属种质资源进行鉴定。
关键词 野牡丹属;种质;分子标记;亲缘关系;指纹图谱
中图分类号 S682.39 文献标识码 A
野牡丹科(Melastomaceae)植物全世界约240个属,共3 000余种[1]。野牡丹属(Melastoma)属于野牡丹科,大约有100个种,分布中心处于东南亚一带,中国有9个种和1个变种,主要分布带处于长江以南各个省区[2]。野牡丹属植物花色有白色系、粉色系、紫色系,其花期长、花量大,株型从地被、灌木到小乔木均有分布,园林运用上可做到周年有花(花期:地菍、印度白花野牡丹四季开花,展毛野牡丹3~4月,紫毛、多花、细叶野牡丹、毛菍5~6月份,野牡丹、毛菍7~8月),具备良好的观赏性状;此外,该属植物的药用价值近来也有深入研究[3-4],发展前景广泛。目前,绝大多数的野牡丹属植物仍未被人工开发利用,已在野牡丹属种质资源的收集及评价[5-9]、栽培技术[10]、脱毒育苗[11-13]、传粉特性[14-15]以及种子发育[16]等方面进行了研究。
目前,针对野牡丹属的分类研究主要集中于宏观形态学[16]、细胞遗传学[17]、孢粉学[18]、细胞学[19]、表型多样性[20-21]等方面,利用分子标记对野牡丹种质资源的遗传性进行研究还比较少。郑涛等[22]利用ISSR分子标记对福建省33份野牡丹属种质资源的遗传多样性进行研究,聚类分析表明,34份材料可划分为3个类群5个亚类,主坐标散点分析可分为4个类群,这与根据形态学划分的结果基本一致。野牡丹属因为分布范围广、自然杂交类型丰富,一些野生种质的分类尚未十分明确,给人工杂交亲本选择带来了不少困扰。因此,本研究在前期研究的基础上扩大了取样范围,以中国的44份野牡丹属种质为研究对象, 利用ISSR和RAPD分子标记技术,再次从分子水平上探讨野牡丹属内的亲缘关系,为有效利用分子标记评价野牡丹属种质资源遗传多样性及遗传基础提供理论依据,同时,利用筛选出的通用引物构建44份野牡丹属野生种质资源的RAPD、ISSR指纹图谱,为野牡丹属野生种质资源快速鉴定及后期人工选育种亲本选择提供理论参考。
1 材料与方法
1.1 材料
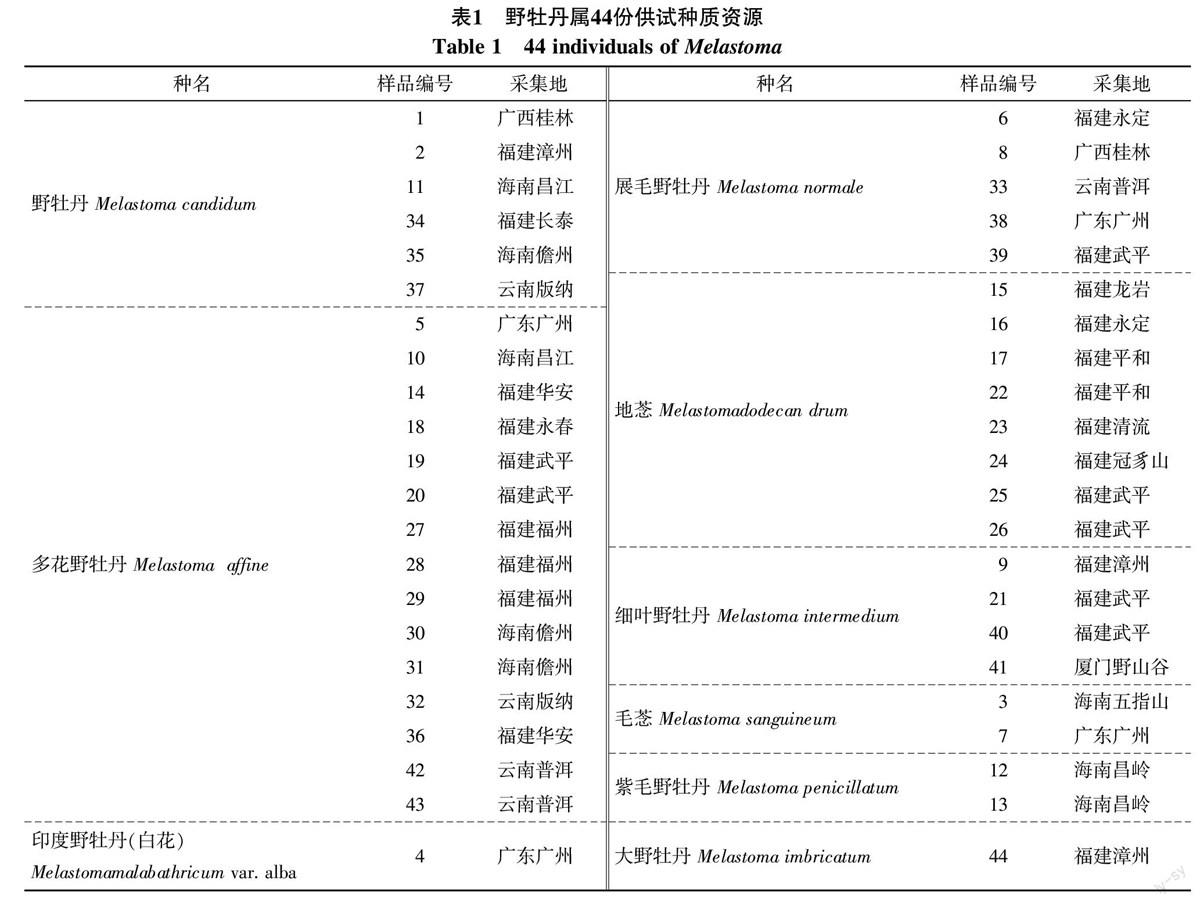
本研究供试材料为野牡丹属野生种质资源共9个种44份材料,来源于福建、广东、广西、海南、云南5个省份。采集新鲜叶片冻于液氮,保存在-80 ℃。
1.2 方法
1.2.1 DNA提取 采用前期试验确定的改良CTAB法提取野牡丹基因组DNA,通过1%琼脂糖凝胶电泳检测,并用分光光度法定量后,-20 ℃保存备用。
1.2.2 PCR扩增 ISSR-PCR反应体系的建立:DNA模板(30 ng/μL)0.7 μL,10×buffer 2 μL,dNTPs(10 mmol/L)0.6 μL,引物(10 μmol/L)0.8 μL,Taq酶(5 U/μL)0.1 μL,ddH2O定容20 μL。反应程序:反应程序:94 ℃预变性5 min;94 ℃变性1 min;53 ℃退火40 s;72 ℃延伸2 min。
PAPD-PCR反应体系的建立:DNA模板(20 ng/uL)1.5 μL,10×Taq Green Buffer(含Mg2+)2 μL,引物(10 μmol/L)0.7 μL,dNTPs(10 mmol/L)0.4 μL,Taq DNA聚合酶(5 U/μL)0.3 μL,ddH2O定容20 μL。反应程序:94 ℃预变性5 min;94 ℃变性1 min,38 ℃退火40 s,72 ℃延伸2 min,45个循环;最后72 ℃延伸5 min,4 ℃保存。
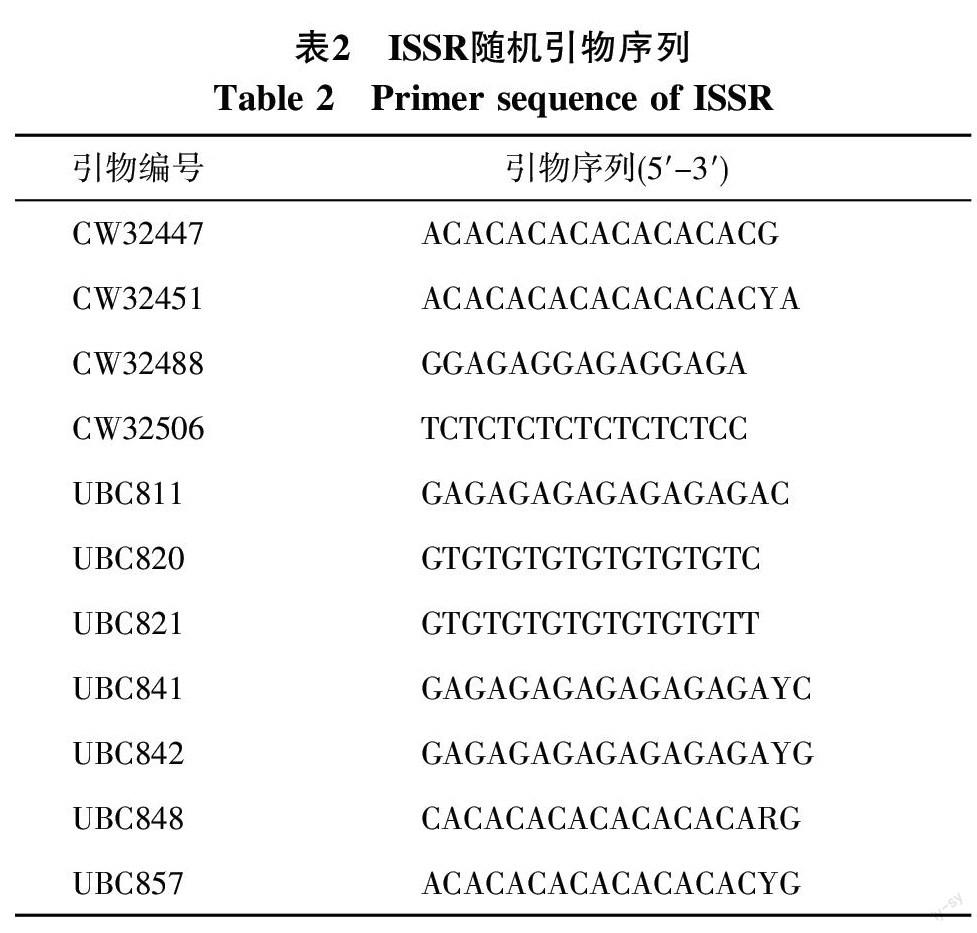
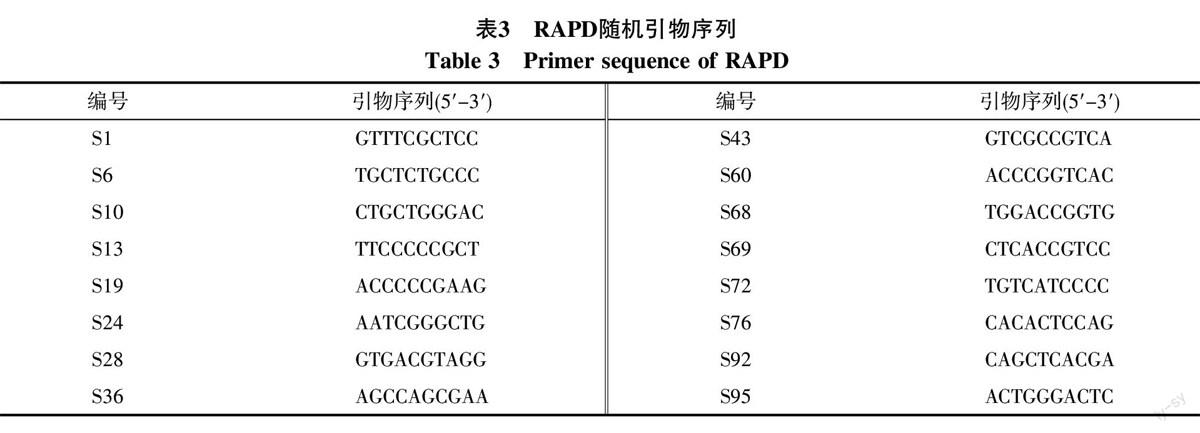
1.2.3 引物筛选 从116条ISSR、100条RAPD随机引物中分别筛选出11条、16条多态性好、条带清晰的引物供PCR反应(表2、表3)。
1.3 数据处理
根据2种分子标记方法通过PCR反应扩增出的条带图片,对同一引物所扩增出迁移率相同的条带计为1个位点,有条带的位置计为“1”,无条带位置计为“0”,所得数据输入Excel 2003工作表建成原始“0/1”数据矩阵,利用NTSYS聚类分析软件进行聚类分析。针对所有供试品种,对11条ISSR引物和16条RAPD引物扩增出的条带进行比对,选出鉴别能力较强的引物,然后对由这些引物经PCR反应跑出的电泳条带进行形象化处理,同一位点上记为“1”的电泳条带用黑色方块表示,记为“0”的条带用白色方块表示,据此构建出供试的44份野牡丹属种质资源的DNA指纹图谱。
2 结果与分析
2.1 ISSR分子标记结果与分析
本研究以前期试验筛选出的11条ISSR随机引物对供试的44份野牡丹属种质资源进行DNA多态性检测[16],总共扩增出91条清晰度好,重复性高的谱带,其中多态性谱带有90 条,多态性条带比例为98.9%(表2)。扩增结果表明,扩增位点最多的引物为UBC857 、UBC811和CW32506,位点数均为12个。其中引物UBC857对44份野牡丹属种质DNA的扩增图谱见图1。
2.2 RAPD标记结果与分析
16条RAPD引物共扩增出113条谱带,其中101条多态性条带,多态性比率为89.38%。扩增产物大小集中在2 00~2 000 bp。扩增位点最多的引物是S1、S19、S43,位点数均为9。其中引物S19对44份野牡丹属种质DNA的扩增图谱见图2。
2.3 44份野牡丹属种质亲缘关系分析
综合44份野牡丹属种质ISSR和RAPD扩增条带数做聚类分析,聚类结果见图3。可将供试野牡丹属种质资源分为2大类群。3号毛菍与其他各地的地菍聚为Ⅰ类;其他36份野牡丹属种质聚类Ⅱ类。Ⅱ类野牡丹属种质较多,可细分为3组,1组为1号、2号、4号、5号、6号、7号、8号、10号、9号、12号、11号、18号、13号、19号、21号、20号、33号;2组为30号、31号、32号、34号、35号、36号、37号、38号、39号、40号、41号、43号、44号、;3组为14号、27号、28号、29号福建多花野牡丹与42号云南普洱的多花野牡丹。44份野牡丹属种质资源遗传相似系数在0.61~0.88,平均相似系数为0.75,表明44份野牡丹属种质之间的亲缘关系较近。
2.4 44份野牡丹属种质指纹图谱的建立
由于ISSR引物的多态性,同一引物扩增出的44份野牡丹属种质资源的条带有明显差异。比对结果表明:利用引物UBC811可以鉴别出38份野牡丹属种质资源,利用引物UBC851可以鉴别出42份野牡丹属种质资源,而采用引物UBC811和UBC857组合可以鉴别出供试的44份野牡丹种质资源。
从筛选16条ARAPD引物中选择多态性好、鉴别效率高的核心引物S19和S43,建立44份野牡丹属种质资源的指纹图谱(图5)。利用引物S19可以鉴别出30份野牡丹属种质资源,利用引物S43可以鉴别出29份野牡丹种质资源,而采用引物S19和S43组合可以鉴别出供试的44份野牡丹属种质资源。由图5可知,利用2条RAPD核心引物可建立44份野牡丹种质资源的指纹图谱,每个品种都有唯一的指纹图谱。
3 讨论与结论
3.1 分子标记用于野牡丹种质的遗传变异研究
采用ISSR分子标记研究44份野牡丹种质的遗传变异情况,10条ISSR引物共扩增出91条带,其中多态性条带90条,多态性比率98.9%;采用RAPD分子标记对44份野牡丹种质的遗传变异进行研究,16条PAPD引物共扩增出113条带,其中多态性条带101条,多态性比率89.38%。可见,2种分子标记均可检测出野牡丹种质资源间较为丰富的遗传变异;但比较起来,ISSR分子标记检测出的多态性比率比RAPD分子标记检测出的高。采用ISSR分子标记研究野牡丹属种质间的遗传变异,其多态性位点的百分率为98.9%,高于新疆红花(Canluumts tinctorius)[21]品种间的93.77%、菊花(Dendranthema×grandi flora)[22]品种间的92.5%和君子兰[23](Clivia)品种间的81.95%;采用RAPD标记研究野牡丹种质的遗传变异,其多态性位点的百分率仅为89.38%,接近于亚麻(Linum usitatissimum)[24]品种间的90.49%,高于石蒜(Lycoris Herb.)[25]品种间的70%,低于红花(Canluumts tinctorius)[21]品种间92.31%。2种分子标记遗传相似性分析结果表明,野牡丹属种质间的遗传相似系数0.61~0.88之间,表明44份野牡丹属种质资源之间亲缘关系较近。
3.2 野牡丹属种质的遗传变异分析
与之前的研究结果[22]对比,本研究扩大了取样范围,从外观形态上判断,增加了《福建植物志》以外的3个种(紫毛、印度、大野牡丹)进行比较分析,得到较为全面的分析数据。本研究2种分子标记方法都将地菍单独聚为一类,这与先前的研究结果[22]相同,表明地菍与其它种间的亲缘关系相对较远,在自然杂交过程中的参与度较低;4号印度白花野牡丹是印度野牡丹(Melastoma malabathricum,粉色花)的变异种,研究结果将其与5号广州多花野牡丹聚在一起,说明二者在亲缘关系上非常接近。研究结果也发现,通过形态学观测很难准确的将野牡丹属中的几个种进行区分,甚至发生错误分类,例如,44号大野牡丹叶片、花瓣显著大于其他样品,形态特征与大野牡丹类似,通过形态学分类很容易将其归为大野牡丹类,而标记结果则表明其应为多花野牡丹类群;32号云南版纳的多花野牡丹应为野牡丹类群;37号云南版纳的野牡丹,与38、39亲缘关系最近,兼有野牡丹、展毛野牡丹的特征,因此将其归为展毛野牡丹更为准确。另外,野牡丹属亲缘关系远近不但与种的类别有关也与地域分布存在一定关系,相同来源地的野牡丹属植物因种间差异可能亲缘关系较远,如10号与12、/13号,而不同来源地的植物因种间差异较小可能被聚为一类,如30、/31号,这也说明多花野牡丹种内存在比较明显的基因分离现象,遗传背景比较复杂。从本研究的样品分析来看,地菍具有较强的遗传稳定性,其余野牡丹属种间、不同来源地的同种植物存在明显的基因交叉,再次证明了前期研究得到的野牡丹属植物“遗传背景复杂、自然杂交类型丰富、亲缘关系总体较近”的结论。
本研究分别筛选出2条特异引物建了供试的44份野牡丹属种质资源的ISSR和RAPD指纹图谱,采用这2种指纹图谱可对供试的所有野牡丹属种质行鉴定,为利用分子标记技术快速、准确地鉴定野牡丹属资源奠定基础。
参考文献
[1] 陈 介. 中国植物志, 第53卷第1分册[M]. 北京: 科学出版社, 1984: 137-190.
[2] 胡松梅, 蒋道松, 龚泽修. 野牡丹属植物研究进展[J]. 现代园艺, 2007(5): 3-6.
[3] 梁春玲, 周玖瑶, 吴俊标, 等. 野牡丹抗炎镇痛作用及其对小白鼠免疫功能影响的研究[J]. 中国药师, 2012, 15(11): 34-36.
[4] 刘 惠, 沈毅华, 刘 文. 野牡丹提取物对血小板聚集的影响[J]. 广东医学院学报, 2012, 30(5): 12-15.
[5] 彭东辉, 张启翔, 黄俊婷. 中国野牡丹科观赏植物种质资源及其在福建省的分布初步调查[J]. 中国园林, 2008, 23(11): 83-88.
[6] 林秀香, 苏金强, 黄阿凤. 福建野牡丹科植物资源初步调查及评价[J]. 福建热作科技, 2003, 28(4): 17-19.
[7] 马国华, 林有润. 华南野牡丹科野生花卉种质资源的收集和繁殖[J]. 中国野生植物资源, 2001, 20(6): 72-73.
[8] 范建红, 冯志坚. 广东野牡丹科野生观赏植物资源[J]. 中国野生植物资源, 2002, 21(4): 19-21.
[9] 尹俊梅, 王祝年, 杨光穗, 等. 海南野牡丹科野生观赏植物种质资源及其开发利用[J]. 热带农业科学, 2006, 26(6): 63-66.
[10] 朱 纯, 陈妙贤, 彭狄周, 等. 10种野牡丹科植物引种栽培及应用研究[J]. 中国野生植物资源, 2006, 25(4): 64-67.
[11] 马国华, 林有润, 简踞光, 等. 野牡丹和地稔的离体培养和植株再生[J]. 植物生理学通讯, 2000, 36(3): 233-234.
[12] 马国华, 张静峰, 刘 念, 等. 从多花野牡丹和野牡丹花柄直接诱导出芽[J]. 植物生理学通讯, 2005, 40(6): 719-719.
[13] 何长信, 代色平, 马国华. 毛菍的组织培养和植株再生[J]. 植物生理学通讯, 2009, 45(1): 49-50.
[14] 彭东辉, 张启翔, 黄启堂, 等. 多花野牡丹传粉生物学观察[J]. 福建林学院学报, 2008, 28(2): 115-120.
[15] 彭东辉, 兰思仁, 吴沙沙. 中国特有种枝毛野牡丹传粉生物学及繁育系统研究[J]. 林业科学研究, 2014, 1: 38-42.
[16] 杨利平, 庄 斌, 苏正华, 等. 野牡丹属植物种子特征的初步研究[J]. 植物遗传资源学报, 2008, 9(2): 248-252.
[17] 张新华, 代色平, 蒋建友, 等. 野牡丹科6种植物染色体数目及核型分析[J]. 热带亚热带植物学报, 2010, 18(4): 386-390.
[18] 刘雪凝, 屈 平. 5种野牡丹属植物花粉形态研究[J]. 河北农业大学学报, 2012, 35(5): 63-66.
[19] 张 媛, 李 燕, 杨利平. 野牡丹属植物的细胞学研究[J]. 湖北农业科学, 2014, 11: 121-124.
[20] 戴小红, 孙伟生, 贺军军, 等. 我国野牡丹属植物的表型多样性研究[J]. 热带作物学报, 2014, 10: 23-26.
[21] 陈进燎, 兰思仁, 吴沙沙, 等. 6种野牡丹属植物叶片表面特征及其分类学意义研究[J]. 福建林学院学报, 2013, 2: 54-59.
[22] 郑 涛, 陈振东, 林秀香, 等. 福建省野牡丹属种质资源的 ISSR分析[J]. 热带亚热带植物学报, 2013, 21(5): 406-413.
[23] 葛 娟, 岳庆妮, 王 蕾, 等. 用RAPD和ISSR法研究新疆红花主栽品种的遗传多样性[J]. 新疆农业科学, 2009 , 46(6): 1 164-1 170.
[24] 缪恒彬, 陈发棣, 赵宏波. 85个大菊品种遗传关系的ISSR分析[J]. 园艺学报, 2007, 34(5): 1 243-1 248.
[25] 郑玉红, 钱美华, 李 莹, 等. ISSR和RAPD分子标记技术在不同君子兰品种遗传多样性上的应用[J]. 北方园艺, 2010(23): 136-139.
[26] Prashant Shekhar, Gupta Sangeeta, Srivast ava Manoj, 等. 基于RAPD-PCR标记的印度亚麻品种遗传多样性和指纹图谱分析[J]. 广西农业科学, 2010, 41(3): 201-206.
[27] 袁菊红, 孙 视, 彭 峰, 等. 石蒜属植物遗传多样性的ISSR和RAPD标记比较研究[J]. 中草药, 2007, 38(10): 1 555-1 561.
猜你喜欢
湖北农业科学(2016年21期)2017-03-18 15:09:28
江苏农业科学(2016年8期)2017-02-15 20:03:31
江苏农业科学(2016年8期)2017-02-15 19:07:41
中国中药杂志(2016年22期)2017-02-13 17:22:57
山东农业科学(2016年12期)2017-01-21 14:26:02
安徽农学通报(2016年24期)2017-01-12 20:34:23
天津农业科学(2016年12期)2017-01-11 23:59:33
中国民族民间医药·上半月(2016年12期)2017-01-11 15:05:31
中国中药杂志(2016年20期)2016-11-19 12:57:20
江苏农业科学(2015年7期)2015-08-20 21:57:46
