
大黄鱼养殖群体和野生群体骨骼micro CT扫描显微结构比较
2016-04-10 01:50赵金良柯巧珍刘家富翁华松韩坤煌
海洋科学 2016年12期
王 映, 赵金良, 柯巧珍, 刘家富, 陈 佳, 翁华松, 韩坤煌
(1. 上海海洋大学 农业部淡水水产种质资源重点实验室, 上海 201306; 2. 大黄鱼育种国家重点实验室,福建 宁德 352103; 3. 宁德市富发水产有限公司, 福建 宁德 352103; 4. 国家级官井洋大黄鱼原种场, 福建宁德 352103)
大黄鱼养殖群体和野生群体骨骼micro CT扫描显微结构比较
王 映1, 赵金良1, 柯巧珍2,3,4, 刘家富2,3,4, 陈 佳2,3,4, 翁华松2,3,4, 韩坤煌2,3,4
(1. 上海海洋大学 农业部淡水水产种质资源重点实验室, 上海 201306; 2. 大黄鱼育种国家重点实验室,福建 宁德 352103; 3. 宁德市富发水产有限公司, 福建 宁德 352103; 4. 国家级官井洋大黄鱼原种场, 福建宁德 352103)
为了了解大黄鱼(Larimichthys crocea)野生群体和养殖群体脊椎骨骨骼显微结构的差异, 本研究运用了Micro CT技术对大黄鱼全鱼骨骼系统进行了扫描与三维重建, 选取野生群体和养殖群体背部和尾部脊椎骨进行了显微结构参数差异分析。结果显示, 大黄鱼脊椎骨与头骨、尾骨相连, 脊椎骨26个,肋骨附着在第1~11腹椎, 无肌间刺, 胸鳍位于头骨后, 背鳍、腹鳍、臀鳍和尾鳍与脊椎骨并不相连。大黄鱼野生群体背部和尾部脊椎骨的组织矿物质密度和骨矿物质密度均显著高于养殖群体(P<0.01),骨体积分数较养殖群体大(P<0.05), 骨小梁间隔较养殖群体小(P<0.05), 骨小梁数量、骨小梁厚度和结构模型指数上没有差别。推测是由于野生群体和养殖群体食物组成、生活环境以及活动情况等差异造成的。
大黄鱼(Larimichthys crocea); 养殖群体; 野生群体; 骨骼; micro CT扫描
大黄鱼(Larimichthys crocea)隶属于鲈形目(Perciformes)、石首鱼科(Sciaenidae), 是中国海水重要经济鱼类之一, 享有“国鱼”之称[1]。自20世纪70年代以来, 由于过度捕捞, 导致大黄鱼野生资源严重衰竭[2]。面对大黄鱼天然捕捞资源的丧失, 为恢复大黄鱼渔业资源, 中国水产科技工作者对大黄鱼人工繁殖与苗种培育进行了大量研究与试验, 取得了一系列显著性成果, 推动了大黄鱼养殖业快速发展[3]。目前, 大黄鱼渔业资源已经转变为人工养殖群体为主、野生群体为辅的状况。但在渔业生产与管理过程中, 对大黄鱼野生群体与养殖群体鉴别尚缺乏科学可行的判别依据。
骨骼是鱼体的基本结构之一, 起支持躯体作用,鱼类骨骼生长发育与遗传、营养、环境等因素密切相关[4], 因此, 骨骼系统不仅成为鱼类分类学和亲缘进化关系研究的重要材料, 同时也是许多鱼类年龄鉴定和生长发育状况分析的重要依据。关于大黄鱼骨骼系统的研究, 丘书院[5]采用腐烂法、鲜煮法对大黄鱼离体骨骼结构组成和形态进行了观察, 刘家富[6]和方家仲等[7]先后观察了大黄鱼胚胎发育及仔稚鱼期骨骼形态特征, 王秋荣等[8]采用二重染色法描述了3~28日龄人工培育大黄鱼骨骼系统发育, 但对大黄鱼骨骼显微结构特征研究还未有报道。
显微CT(Micro-CT)又称小动物CT, 是一种非破坏性的3D成像技术, 可以在不破坏样本的情况下观察动物体内骨骼形态结构, 同时还能获取骨骼内部显微结构特征[9]。在鱼类中, 国内学者仅张宁等[10]利用该技术描述了鲤鱼(Cyprinus carpio)骨骼空间分布结构以及各部位骨骼显微结构间差异。由于营养状况、生活环境存在明显差异, 对大黄鱼野生群体和养殖群体骨骼生长发育状况也会产生不同程度影响,为此, 作者首次采用Micro CT技术对大黄鱼骨骼系统进行显微扫描和三维重构, 比较了野生群体和养殖群体脊椎骨显微结构差异, 旨在为大黄鱼群体种质鉴定、资源管理与利用提供新的证据与技术。
1 材料与方法
1.1 实验材料
大黄鱼野生群体源自2014年10月从福建东冲口海域, 通过声波驱赶鱼群, 将其集中后, 采用小围网撒网捕捞获得。野生个体唇部为胭脂红, 体色金黄,体型细长, 暂养于宁德市国家级大黄鱼原种基地海区鱼排。养殖群体来源于宁德市富发水产有限公司2013年6月人工繁育鱼苗于室内养殖而来, 养殖水温11.8~29.5℃、盐度22.7~30.7、透明度50~150 cm、溶解氧4.7~8.8 mg/L、pH值7.9~8.2, 养殖饲料为大黄鱼膨化颗粒料和软颗粒饲料。2014年11月18日, 随机取野生群体与养殖群体各13尾, 年龄均为1龄(经耳石磨片进行鉴定), 养殖群体体长为6.9~7.3 cm、体质量为57.2~60.7 g, 野生群体体长为7.5~7.9 cm、体质量为54.6~59.4 g。
1.2 micro CT分析
将大黄鱼野生个体和养殖个体分别编号, 逐尾放入Micro CT μ80显微CT仪(SCANCO公司, 瑞士)中, 对全鱼进行CT扫描。扫描条件: 电压(70 kVp),电流(114 μA), 360°旋转扫描, 扫描时间为180 min,分辨率14 μm, 帧平均4帧, 角度增益0.4°。对每个样品扫描获得5500张不同截面的1024×1024像素图片, 采用μCT Ray v4.0-1图像处理软件对大黄鱼全鱼骨骼系统进行三维重建。
分别选取背部第4~5段、尾部第22~23段脊椎骨进行骨骼显微结构分析, 采用μCT Evaluation Program V6.5-2软件处理, 定量分析参数包括: 组织体积、骨体积、骨体积分数、骨小梁厚度、骨小梁数量、骨小梁分离度、骨矿物质密度(bone mineral density, BMD, mg/mL), 组织矿物质密度(tissue mineral density, TMD, mg/mL), 结构模型指数(SMI)。上述指标的分析见Pinto等[11]的报道。
上述数据经Excel 2007初步整理后, 使用SPSS 19.0软件分析群体间差异显著性水平[12]。
2 结果
2.1 野生大黄鱼骨骼的三维重构图像与脊椎骨显微结构
大黄鱼骨骼Micro CT扫描后三维重构图见图1-1和1-2。大黄鱼脊椎骨与头骨、尾骨相连, 脊椎骨26个, 肋骨附着在腹椎上, 从1~11腹椎向下成弧形分开排列, 无肌间刺。胸鳍位于头骨后, 背鳍、腹鳍、臀鳍和尾鳍与脊椎骨并不相连。背鳍分布于第3脊椎骨至第21~22脊椎骨之间, 臀鳍连接在腹棘间骨的下端, 分布于第17~20脊椎骨之间, 尾鳍与尾鳍支鳍骨呈扇形连接状。背部第4、5段脊椎骨见图1-3、1-4、尾部第22-23段脊椎骨见图1-5、1-6。
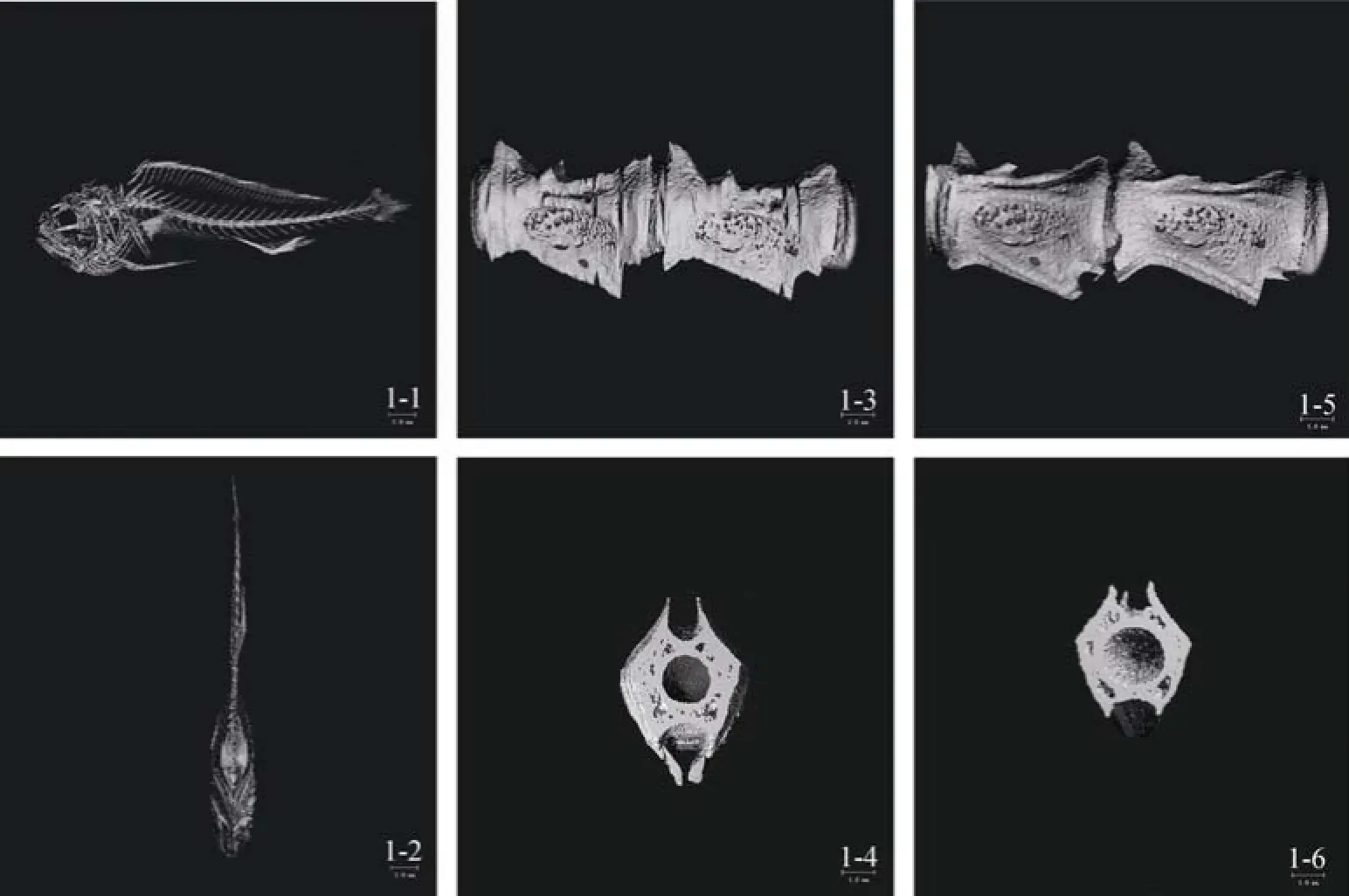
图1 大黄鱼骨骼系统三维重建Fig. 1 Three-dimensional reconstruction of skeleton of Larimichthys crocea
2.2 脊椎骨显微结构参数比较
大黄鱼野生群体与养殖群体背部、尾部脊椎骨显微结构参数见表1, 野生群体背部、尾部脊椎骨骨体积分数分别为46%、41%, 均显著大于养殖群体40%、35% (P<0.05)。野生群体背部、尾部骨小梁分离度分别为0.74、1.04 μm, 显著小于养殖群体0.96、1.28 μm(P<0.05)。野生群体背部组织矿物质密度(150.76 mg/mL)和骨矿物质密度(240.67 mg/mL)极显著高于养殖群体(TMD=128.77 mg/mL、BMD=208.43 mg/mL) (P<0.01); 尾部组织矿物质密度(127.69 mg/mL)和骨矿物质密度(214.77 mg/mL)极显著高于养殖群体(TMD=109.18 mg/mL、BMD=187.65 mg/mL)(P<0.01)。在骨小梁数量、骨小梁厚度和结构模型指数参数上, 野生群体和养殖群体背部、尾部脊椎骨间没有显著差异。
两个群体尾部脊椎骨骨体积分数、骨矿物质密度、组织矿物质密度略低于背部脊椎骨, 骨小梁数量减少, 骨小梁间隔增加, 骨小梁厚度和结构模型指数间无显著差别。
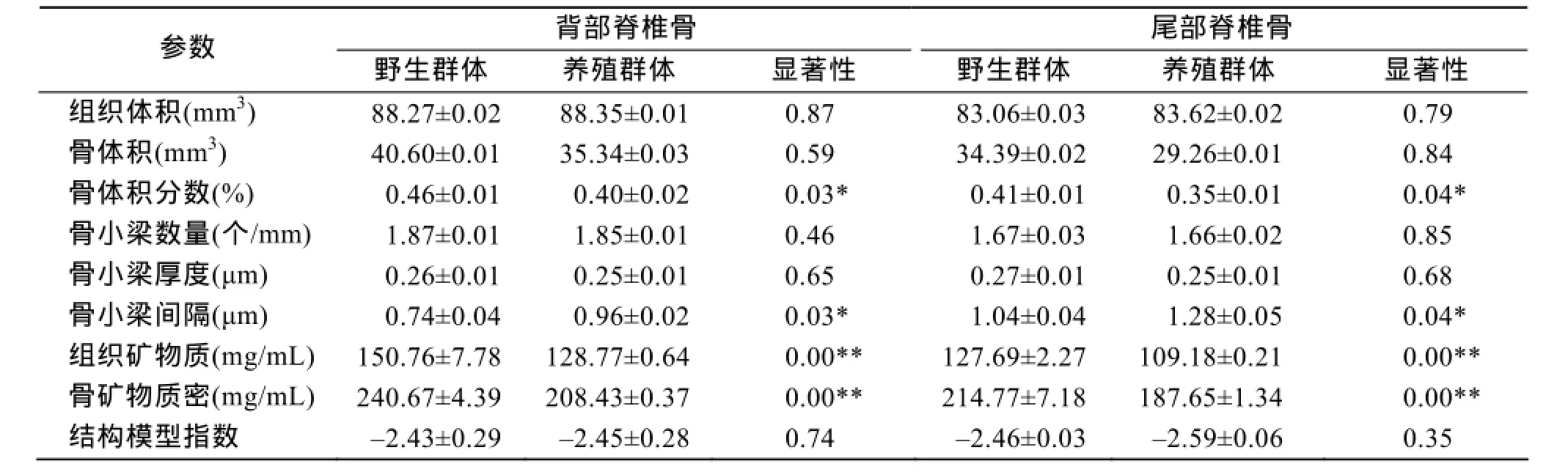
表1 大黄鱼野生群体和养殖群体脊椎骨显微结构参数比较Tab. 1 Comparison of micro structural vertebra parameters in wild and cultured stocks of Larimichthys crocea
3 讨论
骨骼生长发育与遗传、营养、环境因素密切相关, 其中, 营养不仅是骨骼发育的物质基础, 同时,还直接参与骨骼结构建成。本研究中, 大黄鱼野生群体背部、尾部脊椎骨骨体积分数、骨矿物质密度和组织矿物质密度均显著高于养殖群体, 这可能与野生群体、养殖群体的食物营养成分差异、食物量丰廉程度直接关联。野生群体从开口仔鱼起到稚鱼、幼鱼、成鱼, 其适口饵料依次为轮虫(Rotifer)、甲壳类无节幼体、桡足类(Copepods)、磷虾(Euphausia)、糠虾(Mysis)、莹虾(Lucifer)、毛虾(Acetes)和其他小型鱼、虾、蟹等[13], 饵料生物种类多、营养组成全面,有利于骨骼发育与矿化, 因而, 骨矿物质密度和组织矿物质密度相对较高; 而养殖群体鱼苗、鱼种培育过程完全摄食人工配合颗粒饲料, 目前, 由于对不同发育时期大黄鱼的营养需求研究尚不全面[14], 人工配合饲料在营养成分组成及比例上与天然饵料存在差异, 尤其是对骨骼发育的营养物质, 如矿物质、维生素、添加剂等。张春晓[15]研究发现, 大黄鱼鳞片、脊椎骨和全鱼体中磷含量随饲料可利用磷的升高而显著升高, 脊柱骨和全鱼磷含量在饲料可利用磷为0.91%时达到稳定, 磷缺乏或不足时, 不仅抑制大黄鱼生长, 还会影响骨骼矿化。张璐[16]研究发现, 当饲料中维生素D的含量在32.7~1524.5 IU/kg时, 大黄鱼脊柱骨、鳃盖骨、鳞片灰分以及脊柱骨钙含量表现出显著上升趋势, 维生素D过高或过低时均会导致骨骼矿化异常。张佳明[17]研究发现, 饲料中锌、铁含量显著影响大黄鱼脊椎骨中铁、锌的含量, 大黄鱼骨骼中铁、锌含量的数值随饲料锌水平的上升而增大, 在大黄鱼铁、锌满足需求后趋于平稳, 铁、锌缺乏均会降低大黄鱼骨骼矿化程度。由于人工饲料中营养物质的不全面, 养殖群体骨骼的发育程度会受到一定程度的限制, 因而, 出现脊椎骨骨体积分数低于野生群体。此外, 在大黄鱼肌肉营养成分比较研究中,野生群体与养殖群体在肌肉脂类和脂肪酸组成上存在明显的差异, 这主要与它们的生活环境、运动状况和食物丰富程度差异有关[18]。在不同养殖模式、投喂不同饵料及不同品系大黄鱼营养成分比较中, 饵料被认为是影响鱼类全鱼和肌肉氨基酸组成差异的主要原因之一[19]。这些从另一方面佐证了营养组成对大黄鱼野生群体与养殖群体骨骼发育差异存在显著影响。
除营养外, 养殖环境对鱼类骨骼发育也存在一定影响。本实验中, 养殖群体是在养殖车间内进行全程饲养, 水体空间狭小, 水体更换低, 水质条件与化学成分与野生群体生活的自然海区间存在明显差异。这在一定程度上也会影响大黄鱼野生群体、养殖群体骨矿化程度差异。
目前普遍认为, 运动有利于骨骼发育以及钙磷代谢, 加速矿物质的骨内沉积, 并促进骨的生长、骨密度增大。本研究中, 野生群体脊椎骨骨小梁分离度显著小于养殖群体, 表明野生群体骨小梁排列紧密,这可能与野生群体和养殖群体的活动差异有关。野生群体生活在海洋中, 活动范围大, 主动索饵以及躲避敌害攻击时需要长途游泳, 运动强度大、运动量大, 骨小梁分离度低。而养殖群体生活在空间相对狭窄的室内养殖池中, 饲料来源于人工定时投喂、饲料充足, 无需长距离索饵和躲避敌害, 其运动强度和运动量要远低于野生大黄鱼, 骨小梁分离度高。
结构模型指数(SMI)是用于评价骨小梁板状和杆状化程度, 从理论上讲, 完整杆状骨小梁SMI理论值是3, 板状骨小梁的SMI理论值是0, 致密结构可为负值。本研究中, 大黄鱼背部、尾部脊椎骨SMI均为负值, 表明脊椎骨骨小梁形态为致密结构型,这与脊椎骨是躯干的主要支撑系统、承载鱼体大部分重力、致密结构有利于骨小梁承受较强压力作用有关。此外, 本研究中野生群体和养殖群体在骨小梁数量和骨小梁厚度上无明显差异, 这可能与鱼体骨骼结构和力学性能有关, 其具体原因还有待进一步研究。大黄鱼野生群体和养殖群体背部脊椎骨骨小梁数量较尾部数量多, 说明背部脊椎骨部分与非骨组织的焦点较多, 这也与它们骨矿物质密度值大小分布趋势较为一致。背部脊椎骨属于鱼体的主应力区, 可保持身体整体性和自身重量, 因而, 该重载荷区部位骨质结构参数与尾部间存在差异[10]。
[1] 刘家富, 韩坤煌. 我国大黄鱼产业的发展现状与对策[J].福建水产, 2011, 33(5): 4-8. Liu Jiafu, Han Kunhuang. Current development situation and countermeasure of large yellow crocker industry in China[J]. Journal of Fujian Fisheries, 2011, 33(5): 4-8.
[2] 张其永, 洪万树. 官井洋大黄鱼资源的兴衰演变及其修复对策[J]. 海洋渔业, 2015, 37(2): 179-186. Zhang Qiyong, Hong Wanshu. Resource status and remediation strategy for large yellow croaker in Guanjingyang Bay[J]. Marine Fisheries, 2015, 37(2): 179-186.
[3] 张祖兴, 李明云. 大黄鱼种质资源研究进展[J]. 水产科学, 2006, 25 (7): 376-378. Zhang Zuxing, Li Mingyun. Advancement in research on germplasm resources of large yellow croaker Pseudosciaena crocea[J]. Fisheries Science, 2006, 25 (7): 376-378.
[4] 刘康, 张璐, 刘丽燕, 等. 简述鱼类骨骼畸形及其诱因[J]. 广东饲料, 2011, 20(4): 46-48. Liu Kang, Zhang Lu, Liu Liyan, et al. Description the incentives of fish skeletal deformities. [J]. Guangdong Feed, 2011, 20(4): 46-48.
[5] 丘书院. 大黄鱼骨骼的研究[J]. 厦门大学学报, 1957, 2: 100-109. Qiu Shuyuan. On the skeleton of the great croaker, Pseudsciaena crocea Richardson[J]. Journal of Xiamen University(Natural Science), 1957, 2: 100-109.
[6] 刘家富. 人工育苗条件下的大黄鱼胚胎发育及仔、稚鱼形态特征与生态习性的研究[J]. 海洋科学, 1999, 23(6): 61-65. Liu Jiafu . Study on the development of Pseudosciaena crocea (Richardson) embryo and its morphological characteristics and the ecology of its larval juvenile fish [J]. Marine Sciences, 1999, 23(6): 61-65.
[7] 方家仲, 褚茂兵, 肖勤, 等. 大黄鱼早期发育的形态学研究[J]. 海洋科学, 2003, 27(6): 1-6. Fang Jiazhong, Chu Maobing, Xiao Qin, et al. Morphological studies on the early development of largeyellow croaker Pseudosciaena crocea (Richardson) [J]. Marine Sciences, 2003, 27(6): 1-6.
[8] 王秋荣, 倪玥莹, 林利民, 等. 大黄鱼仔稚鱼脊柱、胸鳍及尾鳍骨骼系统的发育观察[J]. 水生生物学报, 2010, 34(3): 467-472. Wang Qiurong, Ni Yueying, Lin Limin, et al. Development of the vertebral column and the pectoral and caudal fins in larae of the large yellow croaker Larimichthys crocea (Richardson) [J]. Acta Hydrobiologica Sinica, 2010, 34(3): 467-472.
[9] 桂建保, 胡战利, 周颖, 等. 高分辨显微CT技术进展[J]. CT理论与应用研究, 2009, 18(2): 106-116. Gui Jianbao, Hu Zhanli, Zhou Ying, et al. Technology development of Micro-CT with high spatial resolution [J]. CT Theory and Applications, 2009, 18(2): 106-116.
[10] 张宁, 苏胜彦, 董在杰. 基于micro CT扫描技术的鲤骨骼和显微结构分析[J]. 南方水产科学, 2012, 8(6): 44-49. Zhang Ning, Su Shengyan, Dong Zaijie, et al. Study on bone and microstructure of common carp using micro CT scanning [J]. South China Fisheries Science, 2012, 8(6): 44-49.
[11] Pinto M, Jepsen K J, Terranova C J, et al. Lack of sexual dimorphism in femora of the eusocial and hypogonadic naked mole-rat: A novel animal model for the study of delayed puberty on the skeletal system [J]. Bone, 2010, 46(1): 112-120.
[12] Markvart M, Darvann T A, Larsen R, et al. Micro-CT analyses of apical enlargement and molar root canal complexity[J]. International Endodontic Journal, 2012, 45(3): 273-281.
[13] 刘家富. 大黄鱼养殖与生物学[M]. 厦门: 厦门大学出版社, 2013: 1-281. Liu Jiafu. Culture and biology of large yellow croaker[M]. Xiamen: Xiamen University Press, 2013: 1-281.
[14] 何志刚, 艾庆辉, 麦康森. 大黄鱼营养需求研究进展[J].饲料工业, 2010, 31(24): 56-59. He Zhigang, Ai Qinghui, Mai Kangsen. Advances in nutrition and feed for large yellow croaker[J]. Feed Industry, 2010, 31(24): 56-59.
[15] 张春晓. 大黄鱼(Pseudosciaena croces R.)、鲈鱼(Lateolabrax japonicas)主要B族维生素和矿物质-鳞的营养生理研究[D]. 青岛: 中国海洋大学, 2007. Zhang Chunxiao. Studies on nutritional physiology of major B vitamins and mineral-phoshorus for large yellow croaker, Pseudosciaena croces R, and Japanese seabass, Laeolabrax japonicas[D]. Qingdao: Ocean University of China, 2007.
[16] 张璐. 鲈鱼和大黄鱼几种维生素的营养生理研究和蛋白源开发[D]. 青岛: 中国海洋大学, 2007. Zhang Lu. Nutrional physiology of several vitamins and protein sources development for Japanese bass, Laeolabrax japonicas C, and large yollow croaker, Pseudosciaena crocea R.[D]. Qingdao: Ocean University of China, 2007.
[17] 张佳明. 鲈鱼和大黄鱼微量元素-锌、铁的营养生理研究[D]. 青岛: 中国海洋大学, 2007. Zhang Jiaming. Studies on nutritional physiology of Zinc and Iron for Japanese seabass, Laeolabrax japonicas and large yollow croaker, Pseudosciaena crocea R. [D]. Qingdao: Ocean University of China, 2007.
[18] 徐继林, 严小军, 罗瑜萍, 等. 岱衢族野生大黄鱼与养殖大黄鱼肌肉脂类和脂肪酸组成的比较研究[J].中国食品学报, 2008, 8(1): 108-114. Xu Jilin, Yan Xiaojun, Luo Yuping, et al. Comparison of lipids and fatty acids composition in muscle between wild and farmed Pseudosciaena crocea from Tai- chu aace[J]. Journal of Chinese Institute of Food Science and Technology, 2008, 8(1): 108-114.
[19] 周飘苹, 金敏, 吴文俊, 等. 不同养殖模式、投喂不同饵料及不同品系大黄鱼营养成分比较[J]. 动物营养学报, 2014, 26(4): 969-98. Zhou Piaoping, Jin Min, Wu Wenjun, et al. Comparison of nutrient components of large yellow croaker (Pseudosciaena crocea Richardson) cultured in different modes, fed different feeds and from different strain [J]. Chinese Journal of Animal Nutrition, 2014, 26(4): 969-998.
Received:Dec. 30, 2015
Comparison of bone microstructures in cultured and wild stocks of large Yellow Croaker (Larimichthys crocea) using micro CT scanning
WANG Ying1, ZHAO Jin-liang1, KE Qiao-zhen2,3,4, LIU Jia-fu2,3,4, CHEN Jia2,3,4, WENG Hua-song2,3,4, HAN Kun-huang2,3,4
(1. Laboratory of Freshwater Fisheries Germplasm Resources, Ministry of Agriculture, Shanghai Ocean University, Shanghai 201306, China; 2. State Key Laboratory of Large Yellow Croaker Breeding (Fujian Fuding Seagull Fishing Food Co., Ltd), Ningde 352103, China; 3. Ningde Fufa Fishery Company Limited, Ningde 352103, China; 4. National Large Yellow Croaker Original Species Field of Fujian, Ningde 352103, China)
Larimichthys crocea; cultured stock; wild stock; bone; micro CT scanning
To understand the differences in the spine microstructures of cultured and wild stocks of Larimichthys crocea, we used the micro CT scanning technique to capture the full fish of L. crocea. Then, we compared the bone structure parameters of the dorsal and caudal vertebra in the cultured and wild stocks. The results show the head and tail to be directly connected with the trunk and 26 vertebrae, ribs were attached to the 1–11thabdominal vertebrae, and the inter-muscular bones, pectoral fin, dorsal fin, ventral fin, anal fin, and caudal fin were not separated from the vertebrae. Bone mineral density, tissue mineral density, and bone volume fraction of the wild stock were all higher than those of the cultured stock, the trabecular spacing of the wild stock was lower than that of the cultured stock, and there was no difference in the wild and cultured stocks in their trabecular number, trabecular spacing, or structure model index. We attribute these differences to different nutritional, environmental, and movement elements of wild and cultured stocks.
S917.4
A
1000-3096(2016)12-0036-05
10.11759//hykx2015123002
(本文编辑: 谭雪静)
2015-12-30;
2016-03-01
福建省引进重大研发机构资助项目(2013I2005); 水产动物遗传育种中心上海市协同创新中心(ZF1206)
[Foundation: Major R & D Projects Funded by Fujian Province , No. 2013I2005; Aquatic Animal Genetics and Breeding Center of Shanghai, No. ZF1206]
王映(1990-), 女, 硕士研究生, 四川泸州人, 主要从事水产动物遗传育种与繁殖学研究, E-mail: 1185334935@qq.com; 赵金良,通信作者, 教授, E-mail: jlzhao@shou.edu.cn
猜你喜欢
陶瓷学报(2021年5期)2021-11-22
陶瓷学报(2021年5期)2021-11-22
音乐教育与创作(2020年9期)2021-01-04
陶瓷学报(2019年6期)2019-10-27
当代水产(2019年6期)2019-07-25
今古传奇·故事版(2017年23期)2018-01-12
特产研究(2016年3期)2016-04-12
中国卫生标准管理(2015年10期)2016-01-15
故事会(2016年1期)2016-01-05
应用海洋学学报(2015年4期)2015-11-24
