
不同大豆蛋白源对黄金鲈蛋白酶及淀粉酶活力的影响
2016-01-10 02:02吴莉芳瞿子惠杨欢欢王洪鹤
饲料工业 2016年16期
■吴莉芳 周 锴 瞿子惠 杨 兰 杨欢欢 黄 权 王洪鹤
(1.吉林农业大学动物科技学院,吉林长春 130118;2.厦门海博丰水产科技有限公司,福建厦门 361026)
鱼粉是水产动物饲料中必需氨基酸和蛋白质的主要来源。但随着高密度集约化水产养殖业的迅猛发展,鱼粉的需求量增加,而鱼粉资源短缺,寻求鱼粉蛋白源替代品已成为国内外水产养殖工作者的主要研究课题[1]。大豆蛋白源是优质的植物蛋白源,在水产动物饲料中被广泛应用。国内外学者在大豆蛋白源替代鱼粉蛋白源方面,进行了大量研究,如:虹鳟(Oncorhynchus mykiss)[2-3]、大西洋鲑(Salmo salar L)[4-5]等鱼类研究的较为深入。但是,大豆蛋白源因其加工工艺的不同,对水产动物的影响也存在较大的差异。刘兴旺等[6]研究了大豆浓缩蛋白(Soy protein concentrate,SPC)替代鱼粉对大菱鲆(Scophthatmus maximus L)摄食、生长及体成分的影响,结果表明,在大菱鲆的配合饲料中,随着大豆浓缩蛋白替代水平的升高,大菱鲆的摄食率、特定生长率均显著降低,但是,大豆浓缩蛋白替代鱼粉蛋白质20%、40%、60%时,对其蛋白质效率和饲料效率的影响不明显。吴莉芳等[7]在埃及胡子鲇(Clarias lazera)的配合饲料中,利用去皮豆粕不同比例替代鱼粉蛋白质,研究了去皮豆粕替代鱼粉对其消化酶活力和肠道组织的影响,结果表明,当去皮豆粕替代鱼粉蛋白质的45%和60%时,埃及胡子鲇的前肠和后肠蛋白酶活力显著低于对照组,且肠道结构完整性被破坏,部分肠绒毛脱落。徐奇友等[8]研究了大豆分离蛋白(Soy protein isolate,SPI)替代鱼粉对哲罗鱼(Hucho taimen)稚鱼生长、体成分和血液生化指标的影响,结果表明,随着饲料中大豆分离蛋白替代比例的增加,哲罗鱼死亡率迅速增加,增重率下降。Barnes等[9]在虹鳟的研究中发现,当发酵豆粕替代35.0%以上鱼粉时会影响虹鳟的生长性能。向枭等[10]在齐口裂腹鱼(Schizothorax prenanti)幼鱼的饲料中,利用豆粕不同比例(0、20%、40%、60%、80%、100%)替代鱼粉蛋白质,研究发现,当豆粕替代鱼粉蛋白质的60%时,齐口裂腹鱼的增重率、特定生长率、饲料效率均达到最高。
黄金鲈(Perca flavescens)属鲈形目(Perciformes),鲈科(Percidae),鲈属(GenusPerca),又称“丝绸鲈”,是北美地区最受欢迎的淡水名贵鱼类之一[11]。由于具有经济价值高、体形优美、适应性强、病害少等特点,是网箱、池塘、工业化养鱼的主要对象。目前,关于黄金鲈肌肉营养成分与品质评价[12]、温度和pH值对黄金鲈消化酶活力的影响[13]、黄金鲈的人工繁殖[14]、苗种培育[15]及食用鱼养殖[16]等方面已有报道,而关于黄金鲈配合饲料方面,目前尚未见报道。消化酶活力的研究能够为鱼类配合饲料的研制提供一定的理论依据。因此,本研究通过饲养试验,在黄金鲈配合饲料中,利用豆粕和全脂豆粉部分替代鱼粉蛋白质,探讨不同大豆蛋白源对黄金鲈蛋白酶及淀粉酶活力的影响,旨在为合理开发利用大豆蛋白源及优化黄金鲈饲料配方提供参考。
1 材料与方法
1.1 试验饲料
饲料配方及主要营养成分如表1所示,共配制3种等氮(粗蛋白质42.0%)、等能(总能为20.0 MJ/kg)的配合饲料,其中Diet 1为对照组,以鱼粉为主要蛋白源,Diet 2是利用豆粕替代20%鱼粉蛋白质,Diet 3是利用全脂豆粉替代20%鱼粉蛋白,Diet 1、Diet 2组饲料分别以鱼油和玉米油[m(鱼油)∶m(玉米油)=1∶1]为脂肪源,Diet 3组没有添加鱼油和玉米油。面粉和糊精为糖源,纤维素为填充物,将所有原料粉碎后,过60目筛,各原料按配方比例称重后混合均匀,加适量的水,利用电动绞肉机,挤压成粒径为2.0 mm的颗粒饲料,于60℃烘箱中烘干后,置于-4℃冰柜中,保存备用。
1.2 试验鱼
试验黄金鲈鱼种购自浙江省台州市天台县龙溪淡水养殖场,平均体质量(28.71±0.08)g/尾。为了使试验鱼逐渐适应的实验室生态系统的环境条件及试验所用饲料,在正式试验分组前,进行为期15 d的预饲试验,在预饲试验过程中,以对照组饲料,饱食投喂黄金鲈鱼种。
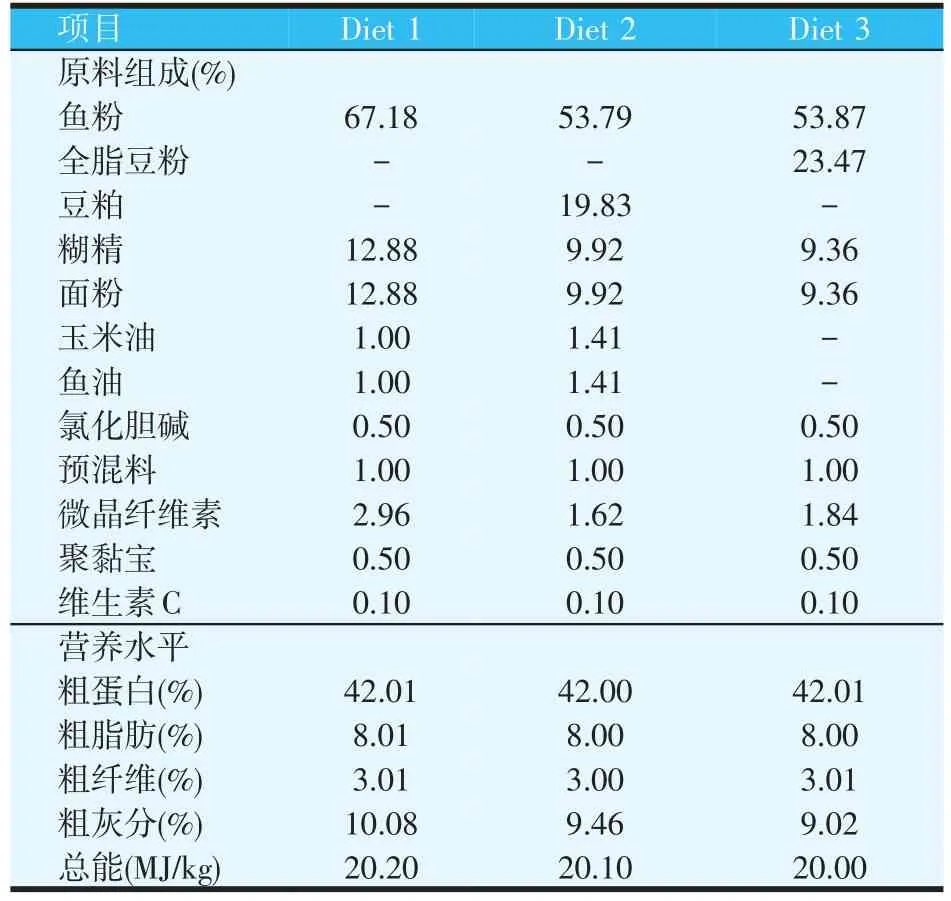
表1 饲料配方及主要营养水平(风干基础)
1.3 试验设计及试验条件
设置3个处理组,Diet 1为对照组,Diet 2为豆粕组,Diet 3为全脂豆粉组,每组设3个重复,每个重复放养黄金鲈鱼种20尾。试验于2013年9月26日~11月20日在吉林农业大学动物科技学院水产养殖实验室的生态系统内进行,共计8周。
试验所用玻璃缸规格为80 cm×60 cm×50 cm,水温控制在22~24℃,pH值为 7.0~8.0,利用增氧气泵连续充气,DO在5.0 mg/l以上,氨氮浓度小于0.3 mg/l,亚硝酸盐浓度小于0.08 mg/l。
1.4 鱼体消毒
预饲试验前,利用浓度为4%的食盐水溶液,药浴黄金鲈鱼种5 min。
1.5 饲养管理
1.5.1 饲料的投喂
每天投喂3次,投喂时间为07:00、12:00、17:00;以4%~6%的投饵率,采用人工手撒的方法,进行饲料的投喂。
1.5.2 水质的管理
试验过程中,每天检测水体的温度、pH值、DO,每周检测氨氮和亚硝酸盐的含量。隔天换水1次,每次换水1/4,在整个养殖过程中,观察鱼的活动情况。
1.6 样品的采集与测定
生长试验结束后,空腹24 h,然后从每个重复中取黄金鲈10尾,用于消化酶活力测定。蛋白酶和淀粉酶活力测定参照吴莉芳等[17]方法进行。
1.7 统计分析
采用SPSS17.5软件,对所得数据进行方差分析,多重比较采用Duncan′s分析组间差异显著性。试验数据用“平均值±标准差”表示,显著性水平设定为P<0.05。
2 结果与分析
2.1 不同大豆蛋白源对黄金鲈蛋白酶活力的影响(见表2)

表2 饲料中不同大豆蛋白源对黄金鲈蛋白酶活力的影响(U/mg protein)(平均值±标准差)
表2表明,本试验条件下,黄金鲈胃蛋白酶活力Diet 2和Diet 3显著低于Diet 1(P<0.05),而Diet 2黄金鲈幽门盲囊、肝脏、前肠及中后肠蛋白酶活力与Diet 1差异不显著(P>0.05);Diet 3黄金鲈的胃、幽门盲囊、肝脏、前肠及中后肠蛋白酶活力显著低于Diet 1(P<0.05)。
2.2 不同大豆蛋白源对黄金鲈淀粉酶活力的影响(见表3)
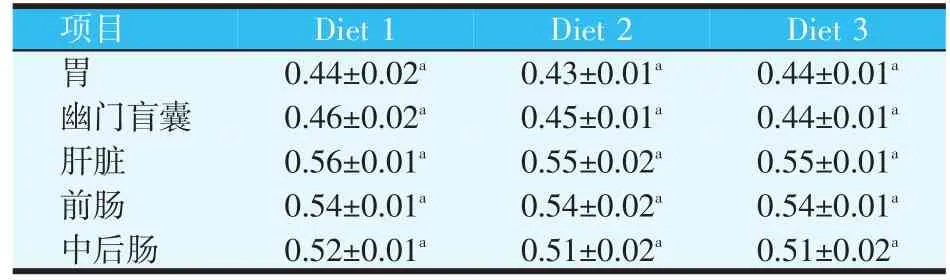
表3 饲料中不同大豆蛋白源对黄金鲈淀粉酶活力的影响(U/mg)(平均值±标准差)
从表3可以看出,Diet1和Diet2和Diet 3黄金鲈的胃、幽门盲囊、肝脏、前肠及中后肠淀粉酶活力差异不显著(P>0.05)。
3 讨论
3.1 不同大豆蛋白源对黄金鲈蛋白酶活力的影响
在黄金鲈配合饲料中,豆粕替代鱼粉蛋白的20%组的幽门盲囊、肝脏、前肠及中后肠蛋白酶活力与对照组差异不显著;而在黄金鲈配合饲料中,当全脂豆粉替代鱼粉20%蛋白时,其幽门盲囊、胃、肝脏、前肠及中后肠蛋白酶活力显著低于对照组。这主要是由于全脂豆粉和豆粕中抗营养因子种类和含量不同及饲料的适口性不同所致。本试验所需全脂豆粉是实验室自行磨制,而豆粕是大豆提取豆油后得到的副产物,在其加工过程中,经过膨化和湿粕脱溶等加工工艺,导致对热敏感的大豆抗营养因子(如:蛋白酶抑制因子、大豆凝集素等)钝化效果较好。从赵元等[18]对不同大豆制品中主要大豆抗营养因子检测结果来看,全脂豆粉中胰蛋白酶抑制因子含量51.00 U/mg,大豆抗原蛋白Glycinin含量167.67 mg/g,β-glycinin含量11.30 mg/g,大豆凝集素含量19.86 mg/g;而豆粕中胰蛋白酶抑制因子含量0.82 U/mg,大豆抗原蛋白Glycinin含量164.71 mg/g,β-glycinin含量18.46 mg/g,大豆凝集素含量5.66 mg/g。因此,豆粕中的胰蛋白酶抑制因子及大豆凝集素的含量低于全脂豆粉。胰蛋白酶抑制因子能够与鱼类消化道蛋白酶结合,形成稳定的复合物,使蛋白酶失活,阻碍鱼类对饲料蛋白质的消化吸收,从而抑制其生长。大豆凝集素与肠上皮细胞结合后,导致其胃肠道结构破坏,使蛋白酶活力下降,生长受阻[19]。吴莉芳等[7]在埃及胡子鲇的配合饲料中,利用去皮豆粕不同比例替代鱼粉蛋白,研究了对其消化酶及肠道结构的影响,从研究结果来看,30%组、45%组和60%组的埃及胡子鲇中肠和肝胰脏蛋白酶活力显著低于对照组,而前肠和后肠蛋白酶活力45%和60%组显著低于对照组,豆粕组胃蛋白酶活力显著低于对照组。
3.2 不同大豆蛋白源对黄金鲈淀粉酶活力的影响
鱼类的各消化器官均存在淀粉酶,鱼类的食性不同,淀粉酶的活力不同,在通常情况下,肉食性鱼类淀粉酶活力低于杂食性鱼类,杂食性鱼类低于草食性鱼类。同种鱼类的不同消化器官、不同生理阶段及同一消化器官的不同部位,淀粉酶活力亦不相同。黄金鲈属于肉食性鱼类,淀粉酶活力较低。本试验的研究结果也证明了这一点。从本试验的研究结果来看,不同大豆蛋白源对黄金鲈的幽门盲囊、胃、肝脏、前肠及中后肠淀粉酶活力无显著影响。本研究结果与吴莉芳等[17]对埃及胡子鲇的研究结果相类似。钱曦等[20]在翘嘴红鲌(Erythroculter ilishaeformis)的配合饲料中,利用豆粕不同比例(0.0%、13.5%、27.0%、40.5%、54.0%)替代鱼粉蛋白,研究了豆粕替代鱼粉对其消化酶活力的影响,从研究结果来看,豆粕替代鱼粉对翘嘴红鲌肠道淀粉酶活力无显著影响。
猜你喜欢
当代水产(2022年8期)2022-09-20
当代水产(2022年6期)2022-06-29
当代水产(2019年11期)2019-12-23
当代水产(2019年7期)2019-09-03
当代水产(2019年6期)2019-07-25
当代水产(2019年6期)2019-07-25
中国粮油学报(2019年4期)2019-07-12
当代水产(2019年1期)2019-05-16
河南畜牧兽医(2017年8期)2017-11-24
食品工业科技(2014年15期)2014-03-11
