
液态发酵制备菜籽粕多肽的动力学研究
2016-01-03 10:59何荣海王治平马海乐
中国粮油学报 2016年9期
何荣海 邢 欢 刘 磊 王治平 马海乐
液态发酵制备菜籽粕多肽的动力学研究
何荣海 邢 欢 刘 磊 王治平 马海乐
(江苏大学食品与生物工程学院;江苏省农产品物理加工重点实验室江苏省农产品生物加工与分离工程技术研究中心,镇江 212013)
研究了枯草芽孢杆菌液态分批发酵菜籽粕的代谢特征,在分批发酵过程中细胞生长曲线呈S形,蛋白消耗曲线则呈反S形变化,多肽浓度在稳定期32 h达到最高10.2 mg/mL。通过SPSS拟合Logistic方较好地描述了枯草芽孢杆菌正常液态发酵过程中菌体生长规律,拟合Luedeking-Piret方程1.068ln(0.99 + 0.01e0.226t)描述了多肽生成的规律,S = 14.430-+ 0.566ln(0.99 + 0.01e0.266t)描述了底物蛋白消耗的规律,模型验证中计算值与实测值的相对误差绝大部分低于10%,较好反映了枯草芽孢杆菌分批发酵过程的动力学特征。
枯草芽孢杆菌 动力学模型 液态发酵 菜籽粕
菜籽粕是菜籽榨油后剩下富含优质蛋白的大宗副产品,但由于在菜籽热榨或溶剂提取制油的过程中,菜籽蛋白严重变性,制约了对菜籽粕的深度利用,目前一般仅用作饲料或肥料,造成蛋白资源的浪费[1-2]。研究表明,以菜籽粕为底物,通过微生物发酵产生的蛋白酶水解菜籽粕中的蛋白质,并降解菜籽粕中的抗营养因子,可提高变性菜籽蛋白的生物利用度,制得具有良好功能特性且粗蛋白含量高的菜籽多肽[3-7],是一种极具前景的菜籽粕深加工技术。枯草芽孢杆菌被广泛应用于医药、农业和科学研究[8],利用枯草芽孢杆菌液态发酵菜籽粕的研究报道不多,而有关枯草芽孢杆菌液态发酵生产多肽过程的生长特性、发酵动力学分析及模型研究文献报道更少,这方面研究的不足阻碍了发酵菜籽粕制备多肽的技术应用。
微生物反应动力学包括细胞生长动力学、代谢产物合成动力学和基质消耗动力学。通过发酵动力学的研究和发酵模型的建立,能够深入了解发酵过程中菌体生长和产物形成的机制。对实现试验数据的模拟放大、发酵条件优化与发酵过程自动化控制,提高产物发酵指标具有重要意义[9-11]。
本研究以实验室保存的枯草芽孢杆菌为试验菌株,进行菌株液态发酵条件优化研究,在此基础上进行了该菌的生长规律、多肽生成及基质蛋白消耗等动力学研究,建立了描述发酵过程变化的动力学曲线。通过动力学曲线了解枯草芽孢杆菌发酵过程的代谢变化,获得其动力学数学模型和模型参数,为进一步放大试验,优化和工业化生产的优化与控制提供参考。
1 材料与方法
1.1 材料及试剂
枯草芽孢杆菌W1-3(Bacillus subtilis),本实验室从镇江恒顺醋业股份有限公司提供的醋醅分离所得,北京诺赛基因公司鉴定;菜籽粕:江苏省丹阳市正大油脂有限公司,粗蛋白质量分数33.14%;Gly-Gly-Tyr-Arg:Sigma公司;其他试剂均为分析纯。
1.2 仪器及设备
BS124S型电子天平:赛多利斯科学仪器有限公司;YX280A型灭菌锅:上海三申医疗器械有限公司;SW-CJ-2FD型双人单面净化工作台:苏州净化设备有限公司;PHS-25型数显pH计:上海精密科学仪器有限公司;UV-1601型紫外可见分光光度计:北京诺利分析仪器公司;LRH-250型生化培养箱:上海一恒科学仪器有限公司;TGL-16型高速台式冷冻离心机:长沙湘仪离心机仪器有限公司;QYC-200型全温空气摇床:上海福玛实验设备有限公司。
1.3 培养基
基础培养基:含1.0 g/100 mL 葡萄糖、1.0 g/100 mL酵母粉、0.5 g/100 mL 氯化钠和2 g/100 mL 琼脂,pH 7.0。
种子培养基:含1.0 g/100 mL 葡萄糖、1.0 g/100 mL 酵母粉、0.5 g/100 mL 氯化钠,pH 7.0。
发酵培养基:含5.0 g/75 mL菜籽粕,pH 7.0。
1.4 培养条件
将试验用菌种接到基础培养基,于30℃条件下恒温静止培养24 h,使菌种转接活化。挑取3环活化后的枯草芽孢杆菌接入装有100 mL种子培养基的三角瓶中,棉塞封口,置于空气摇床中进行扩大培养,温度30 ℃,转速180 r/min,培养24 h,得到试验用种子液。由培养好的菌悬液(菌体浓度为1×108个/mL),每瓶接种量为每瓶1.5 mL,培养基装量75 mL/250 mL 三角瓶,36 ℃,180 r/min 摇床培养。
1.5 发酵液处理
将发酵产物取出,5 000 r/min条件下离心20 min,将上清液过滤后定容到同一体积(100 mL),保存备用。
1.6 分析测定
1.6.1 枯草芽孢杆菌数量的测定[12]
采用血球板计数法测定。
1.6.2 多肽含量的测定[13]
取2 mL样品溶液,加入2 mL 10%的三氯乙酸(TCA)水溶液,于漩涡混合仪上混合均匀,静置30 min,然后在10 000 r/min下离心10 min,将上清液全部转移到50 mL容量瓶中,并用5%的TCA定容至刻度,摇匀。取6 mL上述溶液置于试管中,加入双缩脲试剂4 mL,于漩涡混合仪上混合均匀,静置10 min,2 000 r/min 离心10 min,取上清液于540 nm下测定OD值,与标准曲线(y=0.152x+0.052,R2=0.999)对照计算得出样品溶液中的多肽浓度。
1.6.3 游离氨基酸总量的测定[14]
吸取发酵处理液10 mL 2份,分别置于250 mL的锥形瓶中,加50 mL水,其中1份加入3滴中性红指示剂(0.1%中性红的50%乙醇溶液),用0.1 moL/L NaOH标准溶液滴定至指示剂由红色变成琥珀色(pH=7.4)即为终点,记录消耗NaOH标准溶液体积V1。另1份加入20.0 mL 40%中性甲醛溶液和3滴百里酚酞指示剂(0.1%百里酚酞乙醇溶液),摇匀,静置1 min,用NaOH标准溶液滴定至指示剂变为淡蓝色(pH=10.0)即为终点,记录消耗NaOH标准溶液体积V2。
氨基酸态氮浓度(mg/mL)=c×(V2-V1)×0.014×100
式中:c为NaOH 标准溶液的浓度/mol/L;V1为中性红为指示剂消耗NaOH标准溶液的体积/mL;V2为百里酚酞为指示剂消耗NaOH标准溶液的体积/mL;0.014 为氮的毫摩尔质量/g/mmoL;100 为氨基酸态氮浓度转换系数。
1.6.4 底物剩余蛋白含量的计算
菜籽粕中的蛋白消耗用于菌株生长和维持、多肽生成和游离氨基酸生成,菌株生长而分泌的蛋白酶作用与蛋白质而生成多肽和游离氨基酸,因此菌株生长和维持所消耗的蛋白绝大部分转变成了多肽和游离氨基酸,故蛋白含量可用菜籽粕中总蛋白量减去多肽和游离氨基酸的含量。
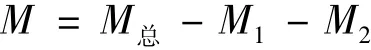
式中:M总为菜籽粕中总蛋白含量/mg;M1为多肽含量/mg;M2为游离氨基酸含量/mg。
2 结果与讨论
2.1 枯草芽孢杆菌发酵过程代谢变化特征
在分批发酵中,枯草芽孢杆菌液态发酵菜籽粕代谢过程变化如图1所示。
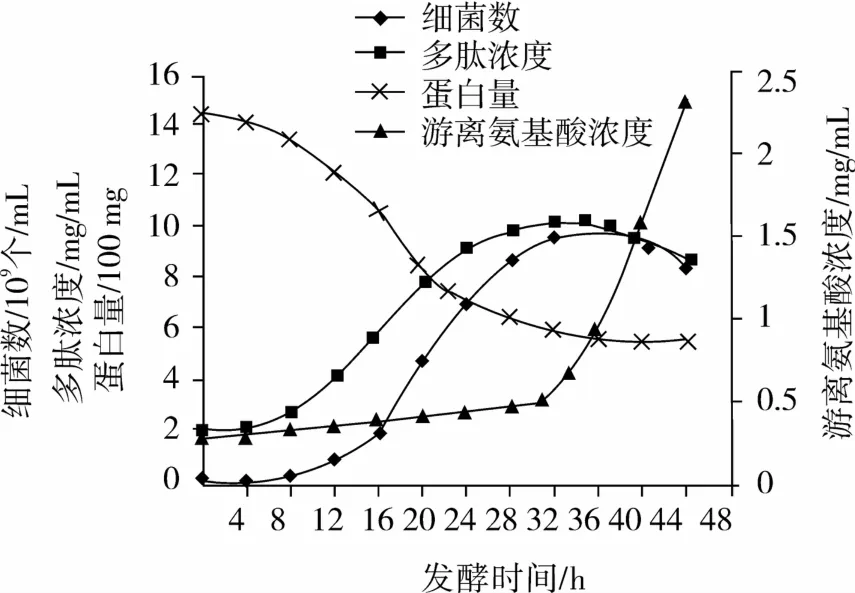
图1 枯草芽孢杆菌分批发酵过程曲线
从图1中可见,该菌生长延迟期较短,发酵0~8 h,菌量增加不明显,多肽基本不生成,蛋白消耗相对较少。8~28 h为生长对数期,菌体数明显上升,多肽产量随着菌体数上升而迅速增加,蛋白消耗明显上升,蛋白量下降到640 mg。28~36 h为平稳期,发酵液中蛋白量变化不明显,菌量也几乎不变,后期会下降,但多肽的浓度还继续上升,32 h达到最高10.2 mg/mL。整个发酵过程从菌体的缓慢生长到达最高的菌体数9.845×109个/mL,多肽浓度从基本不生成到达最高的10.2 mg/mL(多肽得率高达20%),基质蛋白从基本不变到最后基本耗完,整个发酵过程菌体生长代谢正常,多肽浓度达到优化预测值,整个发酵过程没有异常,试验很成功,为以后动力学模型的参数估算提供保障。
总体分析,枯草芽孢杆菌细胞生长曲线为S形,蛋白消耗曲线则呈反S形变化,且多肽为菌体次级代谢产物,8~28 h多肽量随细菌量的增加而增加,而多肽浓度的最高峰却在稳定期中出现,因此多肽的生成属于生长部分偶联型[15]。
2.2 菌体生长动力学
Logistic模型认为微生物生长速率不依赖物质限制[16]。这种动力学的限制因子是细胞浓度或生长时间。Logistic模型是一个典型的S型曲线,能较好地反映分批发酵过程中因菌体浓度的增加对自身生长存在的抑制作用,并能较好地拟合分批发酵过程中的菌体生长规律。因此,本研究采用该模型描述菌体细胞的生长,即
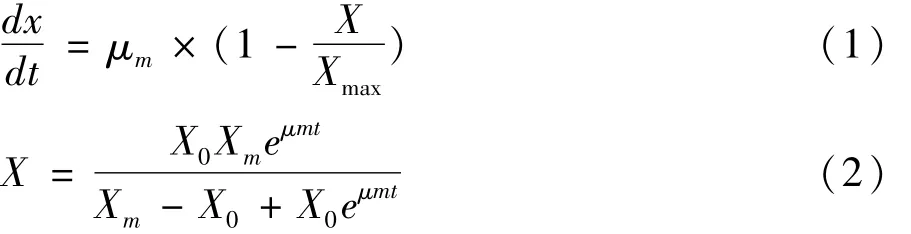
式中:X 为菌体数/109个/mL;μm为最大比生长速率/109个/h;Xmax为菌体数最大值/109个/mL。
以方程(2)为自定义函数,用SPSS软件进行非线性拟合,拟合结果如图2所示。
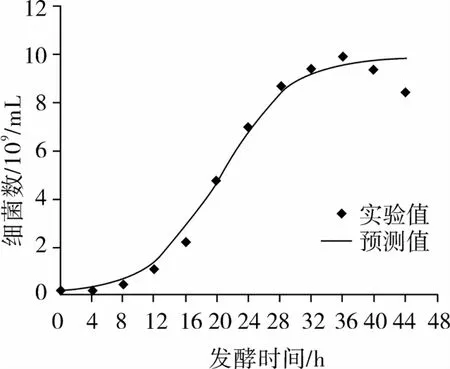
图2 枯草芽孢杆菌生长试验值与模型方程拟合曲线
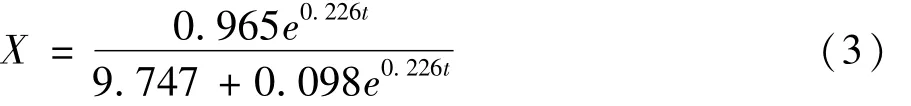
式中:X0=0.098;Xmax=9.845;μm=0.226;R2=0.984。从图2可发现,方程(3)的计算值与试验数据能较好地吻合。因此该方程较好的描述了枯草芽孢杆菌正常液态发酵过程中菌体生长规律,为优化控制提供了参考。
2.3 多肽生成动力学
产物主要指的是细胞培养过程中代谢生成的细胞量以外的产物。按照产物生成与菌体生长的关系,可以分为3类。产物形成与菌体生长偶联型:只是在菌体生长阶段才有产物产生;产物形成与菌体生长部分偶联型:在菌体生长阶段有部分产物形成,而部分或大部分产物是在菌体处于生长稳定期形成的;产物形成与菌体生长非偶联型:只要细胞存在产物就会生成。Luedeking-Piret方程是由Luedeking和Piret提出的用来描述产物形成与细胞生长关系的模型[17]:

当α≠0,β=0时,产物形成为生长偶联型;当α=0,β≠0时,产物形成与细胞生长无关;α≠0,β≠0,产物形成与细胞生长部分偶联。从图1可知,多肽的生成属于生长部分偶联型。通过数学求解得到积分式:
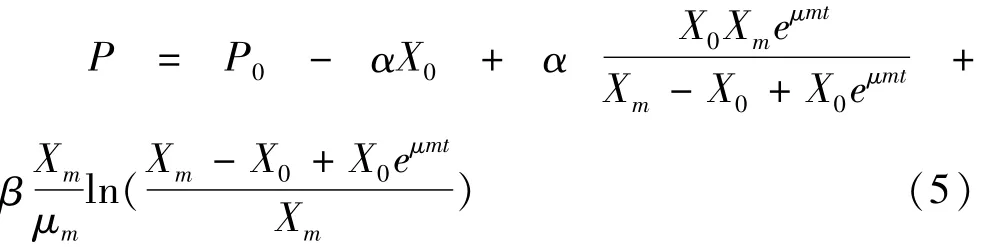
式中:P 为多肽浓度/mg/mL;P0为初始多肽浓度/mg/mL;α,β 为动力学模型参数;α为与菌体生长率相关的产物形成率;βX为非伴随菌体生长的产物形成率。
以方程(5)为自定义函数,用SPSS软件进行非线性拟合,拟合结果如图3所示。
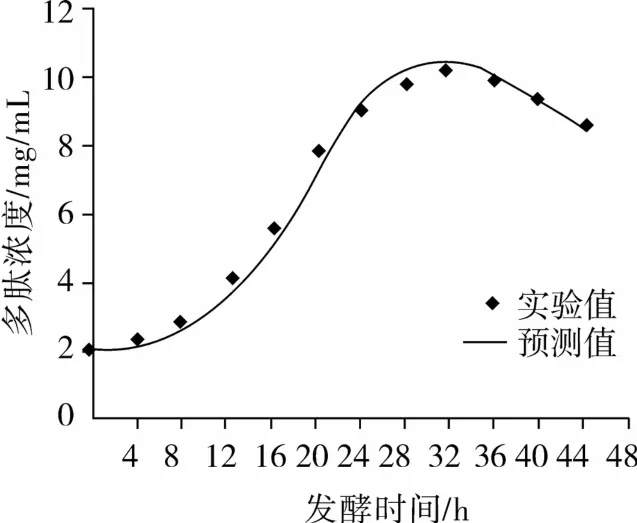
图3 多肽浓度试验值与模型方程拟合曲线
多肽浓度(P)随时间变化的函数为
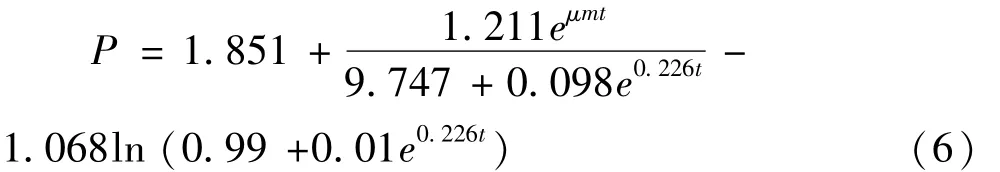
式中:P0=1.974;α =1.255;β = -0.023;R2=0.974。β为负值,说明多肽随着菌株生长而减少,分析其原因为菌株分泌的蛋白酶将菜籽多肽进一步水解成游离氨基酸,导致多肽含量的降低,从图1中可知游离氨基酸浓度从32 h后急剧升高,因此模型准确。从图3中可知方程(6)与试验所得数据能很好地拟合,较好地描述了枯草芽孢杆菌正常液态发酵过程多肽生成情况。
2.4 底物蛋白消耗动力学
在枯草芽孢杆菌液态发酵菜籽粕过程中,菜籽粕中的蛋白主要用于菌株生长、多肽生成、游离氨基酸生成和细胞的维持,多肽和氨基酸的生成是由于菌株生长过程中释放的酶作用于蛋白而生成,可看成菌株生长的一部分,故蛋白消耗可用于Luedeking-Piret方程相似的方程式表示见式(7)。
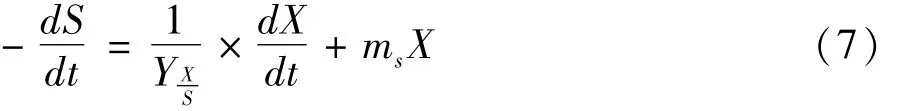
通过数学求解得到积分式:

式中:S为底物蛋白剩余量/×100 mg;S0为初始底物蛋白剩余量/×100 mg;Yx/s为底物用于菌体生长的得率常数;ms为微生物维持常数。
以方程(8)为自定义函数,用spss软件进行非线性拟合,拟合结果如图4所示。
底物蛋白消耗量(S)随时间变化的函数为
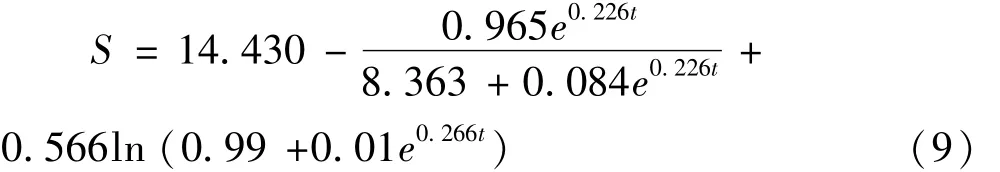
式中:S0=14.316;Yx/s=0.858;ms=0.013;R2=0.968。
从图4中可知方程(9)与试验所得数据能很好的拟合,较好地描述了枯草芽孢杆菌正常液态发酵过程底物蛋白消耗情况。
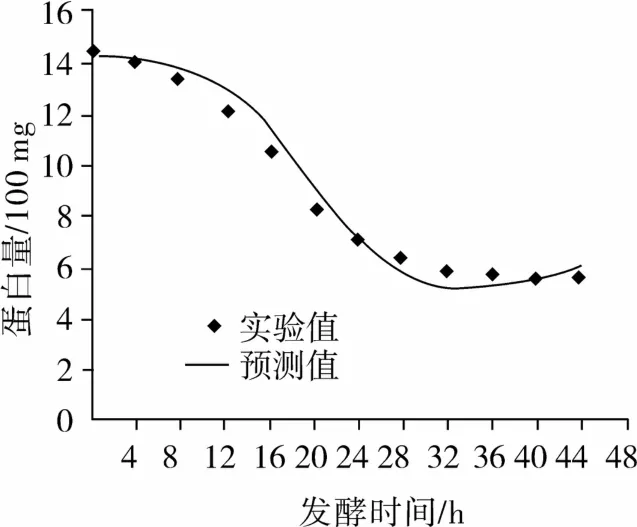
图4 底物蛋白消耗试验值与模型方程拟合曲线
2.5 模型验证
根据试验数据,通过SPSS软件模拟所得枯草芽孢杆菌液态发酵菜籽粕动力学模型参数值见表1。

表1 动力学模型参数
为了所得模型能很好的描述正常发酵过程中菌体生长、多肽生成和基质蛋白消耗,在本试验条件下进行其他发酵试验组,实测值与模型预测值比较结果见表2。从表2可以看出,模型计算值与实测值的相对误差绝大部分低于10%,拟合情况良好。虽然发酵前期(6~14 h)多肽生成模型方程曲线值与实测值误差达到10%以上,但此时多肽浓度及变化不大,对以后菌体的生长及发酵结果影响不大,因此该发酵动力学模型方程能较好地反应枯草芽孢杆菌正常液态发酵菜籽粕的发酵过程。

表2 试验值与模型计算值的比较
3 结论
3.1 通过对枯草芽孢杆菌正常液态发酵菜籽粕发酵特征的分析,以及构建Logistic和Luedeking-Piret方程,建立了描述发酵过程中菌体生长、多肽生成和基质蛋白消耗的动力学模型,经模型验证试验情况良好。本研究创新性地提出了复杂体系发酵底物中的某种成分消耗动力学模型研究方法,并通过发酵中与该成分相关的代谢产物来推出该成分在发酵中的消耗情况,对以后复杂发酵底物的某种成分消耗动力学的研究具有重要的参考价值。
3.2 本研究是在枯草芽孢杆菌正常发酵条件下,但微生物发酵过程的复杂性和影响因素的多变性,使所得发酵动力学模型不能完全反映实际生产中的微生物发酵过程。因此,在今后实际应用中需要对实际发酵过程进行多次优化以及对所建模型进行修正和补充,使模型更好更准确地对实际发酵过程进行预测和控制。
3.3 就本试验而言,8~28 h是菌体生长对数期,是提高菌体浓度的最佳时期;从32 h以后多肽浓度下降,游离氨基酸大度上升,基质蛋白几乎耗完,菌体不再生长。结合动力学模型中微生物维持常数ms较低,用于菌体代谢(包括多肽的生成)的底物较少,因此如果要实现较高的菌体浓度和产肽水平可考虑采取中期(32 h左右)及后期补料。通过发酵动力学的研究,对枯草芽孢杆菌液态发酵菜籽粕有了进一步的了解,对以后发酵过程的工艺控制、改进和放大提供了理论指导。
[1]金晶,徐志宏,魏振承,等.超声微波辅助法提纯菜籽蛋白的研究[J]. 现代食品科技,2009,25(3):275-278
Jin J,Xu Z H,Wei Z C,et al.Ultrasonic and microwave assisted extraction of rapeseed protein [J].Modern Food Science& Technology,2009,25(3):275-278
[2]严奉伟,吴谋成,江洪,等.菜籽粕综合提取工艺研究[J]. 农业工程学报,2004,20(2):209-212
Yan F W,Wu M C,Jiang H,et al.Study on comprehensive extraction technology of rapeseed meal[J].Transactions of the Chinese Society of Agricultural Engineering,2004,20(2):209-212
[3]顾斌,马海乐,刘斌,等.混菌发酵菜籽粕制备多肽技术的研究[J]. 食品工业科技,2011,32(5):190-192
Gu B,Ma H L,Liu B,et al.Study on mixed fermentation of rapeseed meal to prepare peptide [J].Science and Technology of Food Industry,2011,32(5):190-192
[4]顾斌,马海乐,刘斌.菜籽粕混菌固态发酵制备多肽饲料的研究[J]. 中国粮油学报,2011,26(1):83-87
Gu B,Ma H L,Liu B.Preparation of bio-feedstuff with rich peptides from rapeseed meal by mixed solid fermentation[J].Journal of the Chinese Cereals And Oils Association,2011,26(1):83-87
[5]何荣海,蒋边,朱培培,等.枯草芽孢杆菌固态发酵菜籽粕生产多肽及降解硫苷的研究[J].食品工业科技,2014,10(35):228-234
He R H,Jiang B,Zhu PP,et al.The study of Bacillus subtilis fermented rapeseed meal production and degradation of glucosinolate polypeptide [J].Science and Technology of Food Industry,2014,10(35):228-234
[6]Murray D.Rapeseed,a potential global source of high quality plant protein[J].Asia Pacific Food Industry,2001,4(6):30-34
[7]何荣海,刘磊,蒋边,等.枯草芽孢杆菌液态发酵菜籽粕制备抗氧化肽[J].中国食品学报,2013,13(12):12-19
He R H,Liu L,Jiang B,et al.Bacillus subtilis fermentation of rapeseed meal preparation of antioxidant peptides [J].Journal of Chinese Institute of Food Science and Technlolgy,2013,13(12):12-19
[8]李明,双宝,李海涛,等.枯草芽孢杆菌的研究与应用[J]. 东北农业大学学报,2009,40(9):111-114
Li M,Shuang B,Li H T,et al.Study and application of Bacillus subtilis[J].Journal of Northeast Agricultural University,2009,40(9):111-114
[9]谭显东,胡伟,段娅宁,等.三七渣发酵生产蛋白饲料的菌体生长动力学[J]. 中国粮油学报,2014,29(4):72-76
Tan X D,Hu W,Duan Y N,et al.Microbial growth kinetics of protein feed production from notoginseng residues by fermentation [J].Journal of the Chinese Cereals and Oils Association,2014,29(4):72-76
[10]聂国兴,明红,宋东蓥,等.枯草芽孢杆菌E79分批发酵动力学[J]. 化学工程,2011,39(11):6-9
Nie G X,Ming H,Song D Y,et al.Kinetics of batch fermentation of Bacillus subtilis E79 [J].Chemical Engineering,2011,39(11):6-9
[11]郑毅,张志国,关雄.苏云金芽孢杆菌蛋白酶发酵动力学模型的构建[J]. 生物数学学报,2008,23(4):727-734
Zheng Y,Zhang Z G,Guan X.Construction of Bacillus thuringiensis Bacillus protease fermentation kinetics model[J].Journal of Biomathematics,2008,23(4):727-734
[12]张玲.微生物实验指导[M].北京:北京交通大学出版社,2007:54-56 Zhang L.Microbiology experimental instruction [M].Beijing:Beijing Jiaotong University Press,2007:54-56
[13]鲁伟,任国谱,宋俊梅.蛋白水解液中多肽含量的测定方法[J]. 食品科学,2005,26(7):169-171
Lu W,Ren G P,Song J M.Determination of content of peptides in protein hydrolysates [J].Food Science,2005,26(7):169-171
[14]张龙翔,张庭芳,李令媛.生化实验与技术[M].北京:教育出版社,1997
Zhang L X,Zhang T F,Li L Y.Biochemistry experiment and technology [M].Beijing:Educational Press,1997
[15]Huang J X,Yang J S,Wei Y.Dynamic models for polyhydroxybutyrate fermentation process[J].Chemical engineering,2005,33(1):44-47
[16]Elibol M,Mavituna F.A kinetic model for acti-norhodin production by Streptomyces coelicolor A3(2)[J].Process Biochemstry,1999,34(6/7):625-631
[17]陈坚,堵国成,李寅,等.发酵工程实验技术[M].北京:化学工业出版社,2003
Chen J,Du G C,Li Y,et al.Experimental technology of fermentation engineering [M].Beijing:Chemical Industry Press,2003.
Study on Kinetics Model of Peptides Production by Liquid-State Fermentation from Rapeseed Meal
He Ronghai Xing Huan Liu Lei Wang Zhiping Ma Haile
(School of Food and Bioengineering,Jiangsu University/Jiangsu Key Laboratory of Agricultural Product Physical Processing/Jiangsu Province Research Center of Bio-process and Separation of Agri-products,Zhenjiang 212013)
The metabolic characters of Bacillus subtilis in liquid fermentation of rapeseed meal were studied.In the process of batch fermentation,the cell growth curve was S-shaped,protein consumption curve showed anti-S-shaped,and the polypeptide concentration reached the highest 10.2 mg/mL in the stable phase of 32 h.The growth law of the normal liquid fermentation process of Bacillus subtilis was described by the spss matching Logistic equationThe regularity of the formation of polypeptide was described by the Luedeking-Piret equation1.068ln(0.99 +0.01e0.226t),and the law of substrate protein consumption was described by the Luedeking-Piret equation+ 0.566ln(0.99 +0.01e0.266t).The relative error between the calculated and measured values was less than 10% ,which reflected the dynamic characteristics of the batch fermentation process of Bacillus subtilis.
bacillus subtilis,kinetic model,liquid-state fermentation,rapeseed meal
TS229
A
1003-0174(2016)09-0069-06
国家十二五科技支撑计划(2013BAD10B03),国家星火计划(2014GA690262),江苏省自然科学基金(BK2012708),江苏省农产品物理加工重点实验室开放基金(JAPP2010-9)
2015-01-14
何荣海,男,1971年出生,教授,农产品深加工
猜你喜欢
CHINA TODAY(2022年4期)2022-11-22
当代水产(2022年1期)2022-04-26
湖南饲料(2021年4期)2021-10-13
花卉(2021年17期)2021-09-15
食品与发酵工业(2021年7期)2021-04-27
扬子江(2019年3期)2019-05-24
中国调味品(2017年2期)2017-03-20
北方牧业(2016年6期)2016-12-17
猪业科学(2015年11期)2015-01-25
创新科技(2014年12期)2014-07-27
