
沙眼衣原体主要外膜蛋白21-387的原核表达及其免疫原性
2015-12-27 06:12刘巧琼涂建欣林晓云熊一融朱珊丽陈韶张丽芳
温州医科大学学报 2015年2期
刘巧琼,涂建欣,林晓云,熊一融,朱珊丽,陈韶,张丽芳
(温州医科大学 分子病毒和免疫研究所、微生物学和免疫学教研室,浙江 温州 325035)
·论 著·
沙眼衣原体主要外膜蛋白21-387的原核表达及其免疫原性
刘巧琼,涂建欣,林晓云,熊一融,朱珊丽,陈韶,张丽芳
(温州医科大学 分子病毒和免疫研究所、微生物学和免疫学教研室,浙江 温州 325035)
目的:探讨沙眼衣原体(Ct)E血清型主要外膜蛋白(MOMP21-387)的原核表达及其免疫原性。方法:利用PCR方法扩增Ct E血清型MOMP第21至第387氨基酸的基因序列,克隆至pET21a(+)原核表达载体构建重组质粒pET21a(+)/MOMP21-387,并进行原核表达和纯化,经SDS-PAGE和Western blot法分析鉴定后,通过BALB/c小鼠免疫检测MOMP21-387蛋白的免疫原性,即通过ELISA法检测小鼠血清IgG和生殖道分泌物IgA抗体反应,乳酸脱氢酶(LDH)法检测其脾细胞的特异性杀伤作用。结果:在原核表达系统成功表达了MOMP21-387融合蛋白,经SDS-PAGE及Western blot法鉴定在相对分子质量(Mr)约44 000处出现特异性条带;并经Ni-NTA亲和层析的方法获得了纯化的MOMP21-387融合蛋白,免疫BALB/c小鼠可诱导产生特异性血清IgG抗体和生殖道分泌物IgA抗体,至第6周达到高峰,MOMP21-387组的IgG和IgA抗体较PBS组差异均有统计学意义(P<0.05);LDH检测结果显示,MOMP21-387组小鼠脾细胞对靶细胞的杀伤率,在10∶1、20∶1和40∶1和80∶1时均明显高于PBS组,差异有统计学意义(P<0.05)。结论:Ct E型MOMP21-387融合蛋白具有良好的免疫原性,为基于MOMP21-387的Ct的ELISA法检测方法的开发和疫苗研究奠定了基础。
沙眼衣原体;主要外膜蛋白;原核表达;免疫原性
沙眼衣原体(Chlamydia trachomatis,Ct)是全球最常见的性传播疾病(sexually transmitted diseases,STDs)病原体之一,通过性传播感染可致不孕和不育[1],而E型Ct是目前泌尿生殖道感染最常见的血清型之一[2-3]。尽管有抗生素可供治疗,但由于Ct耐药性的增加及其潜伏感染、慢性迁延等特点[4],全球每年生殖道Ct感染的新发病例超过9 000万[5]。因此,如何防治Ct感染是现在的研究热点之一。主要外膜蛋白(major outer membrane protein,MOMP)是各型Ct的主要结构蛋白,含有丰富的抗原表位,是目前Ct疫苗及基于此的亚单位疫苗和表位疫苗研究的最佳靶抗原之一[6-8]。我们通过原核表达系统制备了Ct E血清型的MOMP21-387融合蛋白,并对其免疫原性进行了研究。
1 材料和方法
1.1 材料 Ct E型株购自美国典型物保藏中心(ATCC:VR.348B),由本实验室保存传代。P815细胞由本实验室保存。6~8周龄雌性BABL/c小鼠18只,由上海实验动物中心提供(No.WYDW2012-0041)。E.coli BL21(DE3)、DH5α菌株和质粒pET21a(+)表达载体购自Novagen公司。限制性内切酶、T4 DNA连接酶购自MBI公司。Taq Mix购自天根公司。IPTG、DNA marker、小规模质粒提取试剂盒、琼脂糖胶DNA纯化回收试剂盒均购自上海捷瑞公司。预染蛋白质分子质量(Mr)标准购自Fermentas公司。辣根过氧化酶(HRP)标记山羊抗鼠二抗购自上海联科生物技术公司。小鼠抗His标签的单克隆抗体(mAb)购自上海联科生物技术有限公司,HRP-ECL显色试剂盒购于碧云天公司。MOMP的CTL表位肽(SYVNVNMGL)由上海生工生物公司合成。LDH释放试剂盒购于Promega公司。MOMP21-387基因引物设计与合成:以E 型Ct的MOMP基因为基础,设计合成了两条引物,正向引物为第21个氨基酸编码的核苷酸起始,同时根据pET21a(+)质粒引入酶切位点。正向引入的酶切位点为BamHI,反向引入的酶切位点为SacI。正向引物:引物由上海生工生物公司合成。
1.2 方法
1.2.1 MOMP21-387基因的扩增:以Ct E型标准株为模板,PCR反应体系为:2×Taq Mix 10 μL,模板2 μL,正向引物(10 μmol/L)0.1 μL,反向引物(10 μmol/L)0.1 μL,ddH2O补齐20 μL。PCR反应条件:95 ℃预变性5 min;95 ℃变性30 s,58 ℃退火30 s,72 ℃延伸2 min,循环35次;再72 ℃延伸10 min。10 g/L琼脂糖凝胶电泳鉴定PCR产物,并用DNA产物回收试剂盒纯化PCR产物。
1.2.2 pET21a(+)/MOMP21-387重组质粒的构建和鉴定:将纯化的PCR产物与pET21a(+)载体分别BamHI和SacI双酶切,回收后经T4 DNA连接酶16 ℃酶连过夜,并转化至DH5α感受态中,提取质粒,酶切和测序鉴定。
1.2.3 pET21a(+)/MOMP21-387重组质粒的原核表达:将鉴定好的pET21a(+)/MOMP21-387重组质粒转化入E.coli BL21(DE3)感受态细胞中,37 ℃,250 r/ min培养过夜,按1∶100的比例接种入Amp+LB培养基中,37 ℃,250 r/min振荡摇菌3 h,至吸光度(A)= 0.6~0.8。取出加入IPTG至终浓度为0.6 mmol/L,37 ℃,250 r/min诱导表达4 h,收集细菌。
1.2.4 MOMP21-387融合蛋白的纯化:将收集的细菌用PBS洗涤并重悬,100 W 3 s、工作3 s、间歇超声10 min;12 000 r/min 室温离心10 min,弃去上清,5 mL 2 mol/L尿素重悬,室温静置30 min;12 000 r/min室温离心10 min,5 mL 2 mol/L尿素洗涤1次,弃去上清,用包涵体溶解液(8 mol/L尿素)10 mL,4 ℃溶解包涵体沉淀3 h;12 000 r/min 4℃离心10 min,弃去沉淀,收集包涵体溶解液进一步用Ni-NTA亲和层析法纯化MOMP21-387融合蛋白,并用SDS-PAGE和Western blot法分析鉴定。鉴定成功的MOMP21-387融合蛋白加入蛋白复性液后,置于4 ℃PBS透析过夜。
1.2.5 MOMP21-387融合蛋白的SDS-PAGE电泳和Western blot法分析:将经诱导的细菌离心破碎后用120 g/L SDS-PAGE分离蛋白,考马斯亮蓝染色,分析蛋白表达情况。同时将另一块SDS-PAGE胶转至PVDF膜。用含50 g/L脱脂奶粉4 ℃封闭过夜,TBST洗膜5次,TBST中加入小鼠抗His标签的mAb(1∶10 000),37℃摇床,孵育3 h;TBST洗膜5次,TBST中加入HRP标记的山羊抗小鼠IgG(1∶8 000)作为二抗,37 ℃孵育2 h;TBST洗膜5次,用ECL试剂盒中显色液显色,条带出现后胶片压片。
1.2.6 小鼠免疫程序及标本的收集:6~8周的雌性BABL/c小鼠18只随机分为3组,即MOMP21-387组、Ct灭活组和PBS对照组,每组6只。MOMP21-387组用含弗氏佐剂的MOMP21-387蛋白以每只100 μg的剂量于实验的第1、第3、第5周,分3次,多点皮下注射免疫小鼠。Ct灭活组以IFU(inclusion-forming unit)106/mL的剂量,于65 ℃灭活10 min后,和PBS对照组均取100 μL分别与弗氏佐剂等体积混合后免疫小鼠。实验的第0、第2、第4、第6周小鼠分别采集阴道分泌物和断尾采血。用无菌的PBS冲洗小鼠阴道,每次50 μL,冲洗4次,将冲洗液收集至灭菌的1.5 mL EP管中,加入蛋白酶抑制剂,-80 ℃冰箱保存。断尾取血装入1.5 mL EP管,37 ℃水浴静置1 h,4 ℃,3 000 r/min,离心10 min,小心吸取上层血清于1.5 mL EP管中,-80 ℃冰箱保存。
1.2.7 ELISA法检测体液免疫效应:Ct全菌体于65℃灭活10 min后,超声破菌获得Ct全菌体蛋白(IFU 104/mL)包被96孔酶标板,每孔100 μL,置于4 ℃过夜;洗涤,3 g/L BSA 37 ℃封闭1 h;洗去封闭液后,再分别加入1∶5稀释的待检测的小鼠阴道分泌物100 μL和1∶100的稀释待检测小鼠血清100 μL,37 ℃孵育1 h;PBST反复洗涤后,分别对应加入辣根过氧化物酶标记的山羊抗小鼠IgA(1∶1 000)和辣根过氧化物酶标记的山羊抗小鼠IgG(1∶2 000),37 ℃孵育1 h,酶标仪下于450 nm处读取A值,用以检测小鼠局部黏膜分泌型IgA和血清IgG的表达水平。所有的血清及分泌物标本均重复3孔检测,并设计标准的阳性对照和阴性对照以及本底对照。
1.2.8 LDH释放法检测细胞CTL活性:于动物免疫的第8周,小鼠断头处死,无菌取脾细胞,加入10 μg/mL的MOMP21-387的CTL表位肽(SYVNVNMGL)体外刺激脾细胞4 d得效应细胞;取P815细胞与MOMP21-387的CTL表位肽20 μg/mL置于37 ℃培养箱,50 mL/L CO2共同孵育3 h得靶细胞;将效应细胞与靶细胞按效靶比10∶1、20∶1、40∶1和80∶1加入96孔板中,同时设定效应细胞自发释放孔、靶细胞自发释放孔、靶细胞最大释放孔、培养基背景对照孔和体积校正孔,每种对照设3个平行孔。37 ℃,50 mL/L培养箱共同孵育5 h后,按照LDH释放试剂盒说明书检测CTL活性,以490 nm测得的A值,按以下方法计算。特异性细胞杀伤率=(A实验孔-A靶细胞自发释放-A效应细胞自发释放+A培养基背景对照)/(A靶细胞最大释放-A体积校正-A靶细胞自发释放+A培养基背景校正)×100%。
1.3 统计学处理方法 采用SPSS13.0统计学软件进行统计学处理。所有计量数据资料以±s表示,3组之间的抗体水平差异及特异性细胞杀伤率差异进行两因素的方差分析(Two-way ANOVA),2组之间行两独立样本的t检验分析。P<0.05为差异有统计学意义。
2 结果
2.1 pET21a(+)/MOMP21-387重组质粒鉴定 构建的pET21a(+)/MOMP21-387重组质粒经Bam HI和Sac I双酶切,得到MOMP21-387重组基因片段为1 107 bp,与特异性引物扩增出的PCR产物大小一致,与预期一致(见图1);将酶切鉴定正确的重组质粒进行测序分析,结果正确。
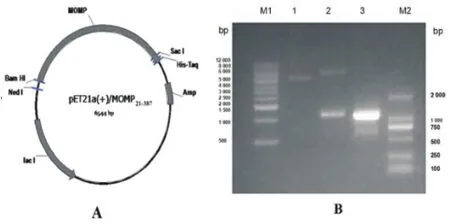
图1 pET21a(+)/MOMP21-387重组质粒图谱及双酶切鉴定结果
2.2 MOMP21-387蛋白的表达、纯化和鉴定 pET21a(+)/ MOMP重组质粒经原核表达系统诱导表达MOMP21-387融合蛋白,通过NI-NTA亲和层析法进行纯化,经SDSPAGE电泳及Western blot法分析验证,结果显示,在SDS-PAGE电泳图上Mr大小约为44 000处出现浓染条带,分子量大小与预期结果一致(见图2A);以小鼠抗His标签的单克隆抗体为一抗的Western blot法检测结果显示在目的位置有单一阳性条带(见图2B)。
2.3 小鼠血清中IgG抗体的ELISA法检测 以灭活的Ct全菌体为ELISA包被抗原,检测各组小鼠血清中特异性IgG抗体结果见图3。3次免疫后,MOMP21-387组及Ct灭活组均诱导小鼠在血清中产生了特异性的IgG抗体,且3组之间差异均有统计学意义(P<0.05)。其中,MOMP21-387组小鼠血清中的IgG抗体表达水平高于PBS组,差异有统计学意义(P<0.05),而MOMP21-387组与Ct灭活组相比,其血清中IgG抗体表达水平差异也有统计学意义(P<0.05)。
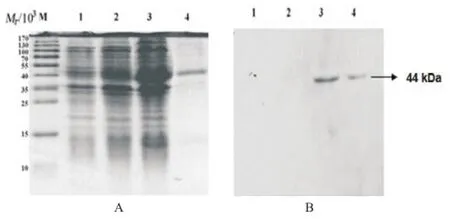
图2 pET21a(+)/MOMP21-387融合蛋白的SDS-PAGE和Western blot法检测结果
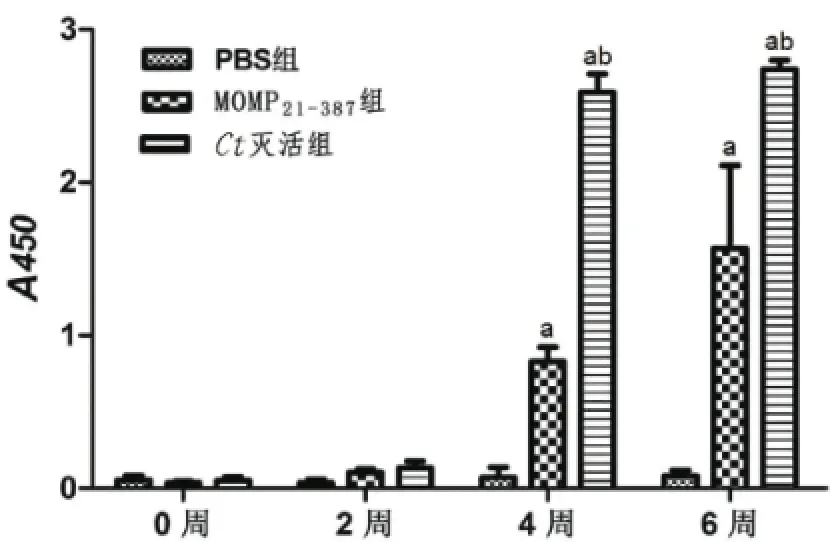
图3 各组小鼠血清特异性IgG抗体的ELISA法检测结果
2.4 小鼠分泌物中IgA抗体的ELISA法检测 以灭活的Ct全菌体为ELISA包被抗原,检测各组小鼠生殖道分泌物中特异性IgA抗体结果见图4。3次免疫后,MOMP21-387组和Ct灭活组均诱导小鼠在分泌物中产生特异性的IgA抗体,且3组之间差异有统计学意义(P<0.05)。其中,MOMP21-387组小鼠生殖道分泌物中IgA抗体表达水平高于PBS组,差异有统计学意义(P<0.05),而Ct灭活组分泌物中IgA抗体表达水平高于MOMP21-387组,差异也有统计学意义(P<0.05)。
2.5 特异性的CTL活性检测 利用LDH释放试剂盒测定效应细胞(脾细胞)对靶细胞(P815)的杀伤结果,以MOMP21-387CTL肽孵育靶细胞及脾细胞,在效/靶比为10∶1、20∶1、40∶1的条件下,Ct灭活组靶细胞杀伤率分别达到:13.4%±3.4%,32.7%±3.9%,42.2%±2.7%,MOMP21-387组的杀伤率分别达到:14.5% ±2.6%,37.3%±1.9%,45.7%±3.5%(见图5)。经两因素方差分析后,效靶比(E/T)及组别均影响杀伤率,两者具有交互作用(P<0.05),且在效/靶比10∶1、20∶1、40∶1及80∶1的条件下,Ct灭活组和MOMP21-387组杀伤效率均明显高于PBS组(P<0.05)。具体来说,MOMP21-387组小鼠脾细胞杀伤靶细胞的效率明显高于PBS,差异有统计学意义(P<0.05),而MOMP21-387组与Ct灭活组比较,杀伤效率差异无统计学意义(P>0.05)。

图4 各组小鼠生殖道分泌物特异性IgA抗体的ELISA法检测结果

图5 各组小鼠脾细胞CTL特异性杀伤结果
3 讨论
基于pET载体的原核表达系统是目前原核蛋白表达引用最多的系统之一,它具有筛选便捷,蛋白高产及纯化简便等优势[9]。但Ct的MOMP全长基因较难在pET载体的原核表达系统中表达,主要原因是组成Ct MOMP蛋白的起始22个氨基酸为信号肽(sign peptide),该信号肽虽不参与MOMP的主要生物学功能,但对大肠杆菌有毒性作用[10-12]。此外,根据影响蛋白稳定性的“N端原则”[13-15],当与N端Met相邻的氨基酸为Arg、Lys、Phe、Leu、Trp、Tyr时,其蛋白容易降解[16]。而Ct E型MOMP全长蛋白,去除信号肽后的第1个氨基酸为Leu(即MOMP的第23个氨基酸),表达后易降解。因此,本研究直接从MOMP氨基酸肽链的第21个氨基酸开始设计,并在pET原核表达系统中成功表达了MOMP21-387融合蛋白。
为避免载体蛋白对目的蛋白的影响,本研究选择PET21a载体,在设计上利用其多克隆位点的Bam HI(GGATCC),利用原核表达系统表达的目的蛋白MOMP21-387尽可能减少载体蛋白片段对实验结果的干扰,经小鼠免疫可产生高效价的抗体,并经Western blot法分析,该蛋白可被特异性抗体识别,表明本研究制备的MOMP21-387融合蛋白具有较强的免疫原性和抗原性。
机体感染Ct可产生体液免疫和细胞的保护性反应,体液免疫表现为产生全身的血清IgG抗体和黏膜局部的IgA抗体[17-19]。本研究利用原核表达的MOMP21-387融合蛋白免疫小鼠,并检测Ct特异性的血清IgG和分泌物IgA抗体水平以评价该蛋白的免疫原性。结果表明,MOMP21-387融合蛋白免疫组的特异性血清IgG及分泌物IgA抗体水平均高于PBS组,且差异具有统计学意义(P<0.05)。该结果与吕慧等[20]利用真核表达系统表达的MOMP蛋白结果相似,均能有效地刺激小鼠产生血清及局部黏膜特异性抗体。此外,本研究也对MOMP21-387融合蛋白所诱导的特异性CTL反应进行了检测,结果表明免疫了融合蛋白的小鼠脾细胞在效靶比为10∶1、20∶1、40∶1和80∶1时,均对靶细胞有很强的杀伤作用,且与Ct灭活组相比,杀伤率差异无统计学意义(P>0.05),表明MOMP21-387融合蛋白有较强的CTL杀伤活性。提示本研究通过原核表达的MOMP融合蛋白有较强的诱导机体产生体液免疫和细胞免疫的免疫原性[18-19]。且MOMP全长蛋白去除起始端信号肽的20个氨基酸残基,并不影响融合蛋白的免疫原性,可刺激小鼠产生良好的特异性体液免疫应答和细胞免疫应答。
综上,本研究在原核表达系统成功表达并纯化了E型Ct MOMP21-387融合蛋白,具有较强的免疫原性,此为后续的基于Ct MOMP的实验室检测方法的开发和疫苗研究等奠定了基础。
[1]Hu VH, Holland MJ, Burton MJ, et al.Trachoma: protective and pathogenic ocular immune responses to Chlamydia trachomatis[J].PLoS Negl Trop Dis, 2013, 7(2): 1-13.
[2]Han Y, Yin YP, Shi MQ, et al.Difference in distribution of Chlamydia trachomatis genotypes among different provinces: a pilot study from four provinces in China[J].Jpn J Infect Dis, 2013, 66(1): 69-71.
[3]Donati M, Di Francesco A, D’Antuono A, et al.Chlamydia trachomatis serovar distribution and other concurrent sexually transmitted infections in heterosexual men with urethritis in Italy[J].Eur J Clin Microbiol Infect Dis, 2009, 28(5): 523-526.
[4]Choroszy-Krol IC, Frej-Madrzak M, Jama-Kmiecik A, et al.Characteristics of the Chlamydia trachomatis speciesimmunopathology and infections[J].Adv Clin Exp Med, 2012, 21(6): 799-808.
[5]幺山山, 曾明.沙眼衣原体疫苗的研发[J].国际生物制品学杂志, 2013, 36(1): 17-21.
[6]Ercolini AM, Miller SD.The role of infections in autoimmune disease[J].Clin Exp Immunol, 2009, 155(1): 1-15.
[7]Findlay HE, McClafferty H, Ashley RH, et al.Surface expression, single-channel analysis and membrane topology of recombinant Chlamydia trachomatis Major Outer Membrane Protein[J].BMC Microbiology, 2005, 5(1): 1-15.
[8]Zhu S, Chen J, Zheng M, et al.Identification of immunodominant linear B-cell epitopes within the major outer membrane protein of Chlamydia trachomatis[J].Acta Biochim Biophys Sin, 2010, 42(11): 771-778.
[9]Wallner M, Gruber P, Radauer C, et al.Lab scale and medium scale production of recombinant allergens in Escherichia coli[J].METHODS, 2004, 32(3): 219-226.
[10] Cheng C, Pal S, Tifrea D, et al.A vaccine formulated with a combination of TLR-2 and TLR-9 adjuvants and the recombinant major outer membrane protein elicits a robust immune response and significant protection against a Chlamydia muridarum challenge[J].Microbes Infect, 2014, 16(3): 244-252.
[11] Pal S, Theodor I, Peterson EM, et al.Immunization with the Chlamydia trachomatis mouse pneumonitis major outer membrane protein can elicit a protective immune response against a genital challenge[J].Infect Immunity, 2001, 69 (10): 6240-6247.
[12] Stephens RS, Mullenbach G, Sanchez-Pescador R, et al.Sequence analysis of the major outer membrane protein gene from Chlamydia trachomatis serovar L2[J].JB, 1986, 168 (3): 1277-1282.
[13] Jones HM, Kubo A, Stephens RS.Design, expression and functional characterization of a synthetic gene encoding the Chlamydia trachomatis major outer membrane protein[J].Gene, 2000, 258(1): 173-181.
[14] Tobias JW, Shrader TE, Rocap G, et al.The N-end rule inbacteria[J].Science, 1991, 254(5036): 1374-1377.
[15] Ucisik MN, Chakravorty DK, Merz KM Jr.Structure and dynamics of the N-terminal domain of the Cu(I) binding protein CusB[J].Biochemistry, 2013, 52(39): 6911-6923.
[16] Hirel PH, Schmitter MJ, Dessen P, et al.Extent of N-terminal methionine excision from Escherichia coli proteins isgoverned by the side-chain length of the penultimate amino acid[J].Proc Natl Acad Sci USA, 1989, 86(21): 8247-8251.
[17] Arsovic A, Nikolov A, Sazdanovic P, et al.Prevalence and diagnostic signifcance of specifc IgA and anti-heat shock protein 60 Chlamydia trachomatis antibodies in subfertile women[J].Eur J Clin Microbiol Infect Dis, 2014, 33(5): 761-766.
[18] Brunham RC, Rey-Ladino J.Immunology of Chlamydia infection: implications for a Chlamydia trachomatis vaccine [J].Nat Rev Immunol, 2005, 5(2): 149-161.
[19] Joyee AG, Yang X.Plasmacytoid dendritic cells mediate the regulation of infammatory type T cell response for optimal immunity against respiratory Chlamydia pneumoniae infection[J].PLoS ONE, 2013, 8(12): 1-13.
[20] 吕慧, 赵蔚明, 于修平, 等.E型沙眼衣原体MOMP基因重组腺病毒的构建及免疫原性研究[J].山东大学学报(医学版), 2006, 44(12): 1189-1193.
(本文编辑:胡苗苗)
Prokaryotic expression and immunogenicity of major outer membrane protein 21-387 from Chlamydia tra-chomatis
LIU Qiaoqiong, TU Jianxin, LIN Xiaoyun, XIONG Yirong, ZHU Shanli, CHEN Shao, ZHANG Lifang.Institute of Molecular Virology and Immunology, Department of Microbiology and Immunology, Wenzhou Medical University, Wenzhou, 325035
Objective:To explore prokaryotic expression and immunogenicity of major outer membrane protein (MOMP21-387) of Chlamydia trachomatis serotype E.Methods:The gene encoding MOMP21-387was amplifed from genome DNA of C.trachomatis E by PCR analysis, and then cloned into pET21a (+) vector to construct recombinant plasmid pET21a (+)/MOMP21-387, and was expressed in the prokaryotic expression system.The MOMP21-387fusion protein was purifed and identifed by SDS-PAGE and western blot analysis.The immunogenicity was further assessed by immunizing BABL/c mice, the reactivity of specifc serum IgG in serum and genital tract mucosal IgA were tested by ELISA, and the specifc cytotoxicity of spleen cells was detected with lactate dehydrogenase (LDH) method.Results: The MOMP21-387fusion protein was successfully expressed in a prokaryotic expression system, and the specifc positive bands at the relative molecular mass (Mr) of about 44 000 was confrmed by SDS-PAGE and western blot analysis; And the purifed MOMP21-387fusion protein was obtained with the Ni-NTA affnity chromatography method.The mice can be induced to produce the specifc serum IgG and reproductive tract mucosal IgA by immunizing with the MOMP21-387fusion protein, and the value of the specifc serum IgG and mucosal IgAin immunized groups were signifcantly higher than that of the PBS control group (P<0.05), the antibodys detected by ELISA reached peak at the 6thweek post-immunization.The LDH analysis showed that, the killing rate of spleen cells to the target cell in MOMP21-387protein immunized groups was signifcantly higher than those in PBS control group, when the effector cells to target ratio reached to 10:1, 20:1, 40:1 and 80:1 (P<0.05).Conclusion:The MOMP21-387fusion protein of Chlamydia trachomatis serovar Eshows good immunogenicity, It may lay the foundation for the Chlamydia trachomatis ELISA detection and vaccine development based on MOMP21-387.
Chlamydia trachomatis; major outer memberane protein; prokaryotic expression; immunogenicity
·消 息·
R374.1,R392-33,Q344.13
A
10.3969/j.issn.2095-9400.2015.02.004
2014-07-21
国家自然科学基金资助项目(30972669);浙江省自然科学基金资助项目(Y2100611)。
刘巧琼(1988-),女,湖南衡阳人,硕士生。
张丽芳,教授,博士生导师,Email:wenzhouzlf@126.com。
猜你喜欢
中国动物保健(2022年2期)2022-05-05
江西农业学报(2021年4期)2021-04-20
上海农业学报(2017年4期)2017-04-10
特产研究(2016年3期)2016-04-12
中国学术期刊文摘(2016年1期)2016-02-13
中南民族大学学报(自然科学版)(2015年2期)2015-12-16
安徽医科大学学报(2015年9期)2015-12-16
西南医科大学学报(2015年1期)2015-08-22
中国当代医药(2015年9期)2015-03-01
现代检验医学杂志(2015年5期)2015-02-06
