
岩生红豆苗木光合特性初步研究
2015-12-22 03:38李晓芳安明态张建利王加国杨焱冰贵州大学林学院贵州贵阳55005贵州科学院山地资源研究所贵州贵阳55000
安徽农业科学 2015年27期
李晓芳,杨 瑞,安明态*,张建利,王加国,杨焱冰(.贵州大学林学院,贵州贵阳55005;.贵州科学院山地资源研究所,贵州贵阳55000)
岩生红豆(Ormoia saxatilia)隶属豆科(Leguminosae)红豆树属(Ormosia),我国特有种,仅产于贵州、湖南。因其树干通直,纹理美观,结构细致,为珍贵用材,大树多被砍伐,已被世界自然保护联盟(IUCN)列为极危树种[1-2]。根据《贵州野生珍贵植物资源》记载以及笔者近年来的调查可知,截至2014年12月,该种仅在贵州省贵阳市黔灵山(模式产地)、修文、望谟、惠水、花溪、三都和独山等17地方的山地林、灌丛或林缘中有分布,各分布点皆十分零散,有的仅分布1、2株。调查同时发现,野生岩生红豆天然更新不良,主要更新方式为根蘖、伐桩萌蘖等。光合作用是绿色植物吸收光能,把CO2和H2O合成有机物释放O2的过程,是生物界获得能量、食物及氧气的根本途径,被称为“地球上最重要的化学反应”[3],它的强弱与植物生产力密切相关[4]。近年来,由于珍贵用材树种受人为乱砍滥伐和自然更新慢等因素影响,其自然资源日渐稀缺,引起越来越多的业内人士的重视和研究。针对被发现较晚、宣传甚少、人们知之甚少的珍贵红木类用材树种——岩生红豆,目前仅笔者所在课题组就其资源现状、濒危原因、天然林主要树种生态位[2]、天然群落特征及物种多样性[5]和种苗繁育[6]等方面作了简单的研究和分析,而对其光合生理方面的研究也尤为重要。该文拟通过对岩生红豆苗木的光合速率、蒸腾速率、水分利用效率、光强—光响应和CO2响应等光合特性的研究,以期为岩生红豆苗木的光照管理与驯化栽培提供理论参考,同时也为其野生种群天然更新的进一步保育提供一定的科学依据。
1 材料与方法
1.1 供试材料与试验地点 该研究在贵州大学林学院(贵阳花溪)实验苗圃内进行,所选材料为生长健康的3年生岩生红豆实生苗木。
1.2 研究方法 选择2014年7月下旬至8月上旬的晴朗天气进行测定,采用的仪器为Li-6400XT便携式光合仪,内容包括光合日变化测定、光合—光响应测定和光合—CO2响应测定。其中,光合日变化相关指标(净光合速率Pn、蒸腾速率Tr、水分利用效率WUE、胞间CO2浓度Ci等)的测定时间为8:00~18:00,每2 h测1次,共测定大小、体型特征和生长基本一致的5株岩生红豆苗木,从每株中选中上部健康、成熟的功能叶(东、南、西、北向各1片)作为测定样叶,每片样叶重复测定5次,计算结果为剔除异常值后的平均值。光合—光响应与光合—CO2响应的测定时间选择在早上9:00~11:00进行,每株测1片叶,重复3次,温度控制为28℃。其中,CO2浓度分别设置 50、100、200、300、400、500、600、800、1 000、1 300、1 600 和2 000 μmolCO2/mol共12 个梯度;光强设置 0、5、10、20、50、100、200、400、600、800、1 200、1 400、1 800 μmol/(m2·s)共13个梯度。因YE[7]的直角双曲线修正模型拟合结果最接近实测值,该研究利用其光合计算软件进行相关光合数据的处理和拟合分析。
2 结果与分析
2.1 光合特征日变化分析
2.1.1 净光合速率与胞间CO2浓度日变化分析。光合速率常指单位时间单位叶面积的CO2吸收量、O2释放量或干物质积累量。胞间CO2浓度(Ci)是光合生理生态研究中常用的一个重要参数,它的变化方向是确定光合速率变化的主要原因是否为气孔因素的必不可少的判断依据[8]。由图1可知,净光合速率与胞间CO2浓度日变化均呈单峰曲线。经过夜间的富集,胞间CO2浓度在清晨较高,随后开始缓慢降低。10:00以前,净光合速率随胞间CO2浓度的降低而逐渐升高,两者呈负相关,说明净光合速率随光强的增加而增加主要是叶肉细胞的光合活性增大的结果。上午10:00净光合速率达到最大,其值7.87 μmol CO2/(m2·s)。10:00 以后,净光合速率随胞间CO2浓度的降低而降低,二者呈正相关,此阶段净光合速率逐渐降低是气孔导度降低引起胞间CO2浓度降低所致。岩生红豆净光合速率日均值为6.43 μmol CO2/(m2·s),高于女贞(Ligustrum lucidum)、茅栗(Castanea seguinii)等阳性树种[9],说明岩生红豆也是阳性树种,具有较强的固定大气CO2的能力和光合能力。
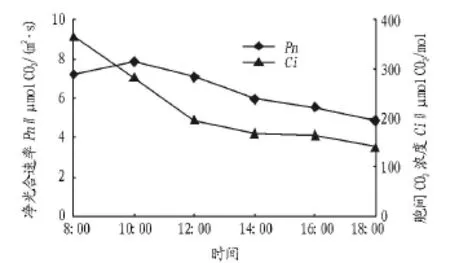
图1 净光合速率与胞间CO2浓度日变化
2.1.2 蒸腾速率及水分利用效率日变化分析。蒸腾速率(Tr)是揭示植物水分状况最重要的生理指标,可表明植物蒸腾作用的强弱,Tr越低说明植物越能适应干旱的环境[10]。水分利用效率(WUE)表示每损失单位数量的H2O固定CO2的数量[8],是植物生存的关键因子[11],客观反映植物对水分利用状况[12]。由图2可知,蒸腾速率日变化呈双峰型曲线,上午和下午各有1个峰值,分别出现在上午10:00[3.89 mmolH2O/(m2·s)]和下午 14:00[3.36 mmolH2O/(m2·s)],存在一定的“午休”现象,这可能是因为中午前后的高光强、高温导致水分亏缺,引起气孔部分关闭,导致对光合作用产生抑制,总体上与前人研究结果[13-17]一致。岩生红豆苗木蒸腾速率日均值为2.62 mmolH2O/(m2·s),略低于鸡冠刺桐(Erythrina crista -galli)[18]、香椿(Toona sinensis)[9]等较为速生的树种,即岩生红豆蒸腾耗水能力低于鸡冠刺桐、香椿等。与蒸腾速率相反,岩生红豆苗木水分利用效率日变化呈“W”型曲线,最大值出现在上午8:00,之后随光强和叶温的增加而迅速下降,10:00开始回升,12:00达到第2个峰值后开始下降,14:00达到第2个谷值后又开始回升,18:00时达到第3个峰值。岩生红豆苗木水分利用效率日均值为2.30 mgCO2/gH2O,低于女贞(Ligustrum lucidum)、构树(Broussonetia papyrifera)等[9],即岩生红豆不能有效地利用土壤水分。因此,栽培岩生红豆苗木时应保证充足的水分供应。
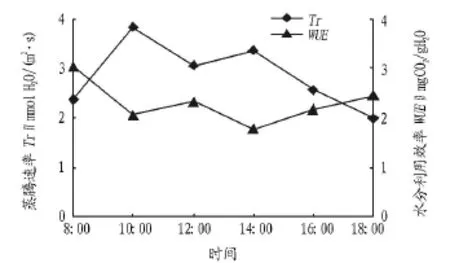
图2 蒸腾速率与水分利用效率日变化
2.2 光强—光响应曲线分析 光强—光响应曲线反映植物光合速率随光照强度增减的变化规律[16],光强过高或过低都不利于植物生长[19]。由图3可知,弱光下,净光合速率随光强的增大呈线性上升,在此光强范围内,光强是光合作用的主要限制因子;当光强继续增加超过一定强度时,净光合速率上升趋势变缓、幅度变小,最后稳定在一个较高的水平,呈现出光饱和现象。通过模拟得出,相关系数R2达0.98,说明该模型能较好模拟真实值,拟合效果较好。光补偿点为21.33 μmol/(m2·s),光饱和点为 1 343.90 μmol/(m2·s),光补偿点低于而光饱和点又高于茅栗(Castanea seguinii)、栾树(Koelreuteria paniculata)及女贞(Ligustrum lucidum)等阳性树种[9],说明岩生红豆属偏喜光树种,对强光和弱光的适应性较强,栽培时应保证足够的光环境。
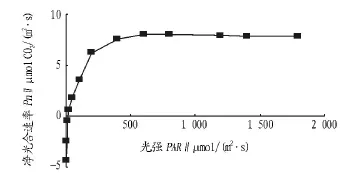
图3 光强—光响应曲线
2.3 CO2响应曲线分析 CO2响应曲线反映植物净光合速率与CO2浓度梯度之间的关系,以及植物对CO2的适应能力[20]。由图4可知,低CO2浓度下,净光合速率随CO2浓度的增加呈比例增加;当CO2浓度增加到一定范围时,净光合速率上升幅度变小,逐渐稳定并趋于饱和。通过模型对CO2响应曲线的拟合,相关系数R2达0.99,说明该模型同样可以较准确地模拟岩生红豆叶片光合速率的CO2响应规律。CO2补偿点为 228.55 μmolCO2/mol,CO2饱和点为 1 480.82 μmolCO2/mol,与香樟、红椿(Toona ciliate)[20]和紫楠(Phoebe sheareri)[21]等珍贵用材树种相比,岩生红豆CO2补偿点较高而CO2饱和点略低,其对CO2和光能的利用能力较弱,其正常生长对大气CO2浓度要求较高,栽培时应保证有充足的CO2供应。
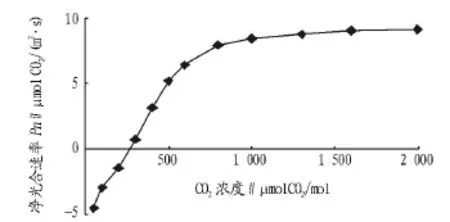
图4 CO2响应曲线
3 结论与讨论
(1)岩生红豆净光合速率日变化呈单峰曲线,上午10:00达到峰值后逐渐下降,未发现光合“午休”现象,1 d中光合效率最佳时段在上午;净光合速率日均值高,属阳性树种,固定大气CO2的能力和强光环境下的光合能力强。蒸腾速率日变化呈双峰曲线,上午和下午各有1个峰值,存在一定的“午休”现象;水分利用效率日变化与蒸腾速率相反,呈“W”型曲线;蒸腾速率与水分利用效率日均值均不高,不能有效地利用土壤水分。
(2)光响应曲线相关系数R2达0.98,说明该模型拟合效果较好;光饱和点高而光补偿点低,对光环境的适应范围广,岩生红豆属于偏喜光树种。CO2响应曲线相关系数R2=0.99,表明模型能够较为准确地模拟其CO2响应规律;岩生红豆CO2补偿点较高而CO2饱和点略低于香樟、红椿和紫楠等珍贵用材树种,说明其正常生长对CO2浓度的要求较高。
(3)野外调查研究亦表明,岩生红豆在疏林下及灌木灌丛中皆能生长良好,但在过荫蔽或光照过强环境下则生长不良,其中以过分荫蔽的环境生长最差(望谟笔架山和贵阳黔灵山皆有此现象)。因此,栽培时给予充足的水分、足够的CO2和光环境可提高成活率,有利于苗木生长发育。
[1]赵莉娜,李金亚,刘慧圆,等.有关IUCN红色名录中“地点”参数合理应用的讨论[J].植物资源与环境学报,2014,23(3):102 -107.
[2]袁丛军,喻理飞,安明态,等.岩生红豆天然林主要树种生态位研究[J].生态科学,2013,32(4):487 -493.
[3]王忠.植物生理学[M].北京:中国农业出版社,2005:121.
[4]李慧,丁莉.不同生境条件下续断的光合特性研究[J].湖北民族学院学报(自然科学版),2011,29(2):227 -231.
[5]袁丛军,安明态,严令斌,等.两种干扰强度下岩生红豆天然群落特征及物种多样性分析[J].南方农业学报,2013(2):280-284.
[6]方小平,张廷忠,姚淑君,等.贵州省林科院树木园珍稀树种引种研究报告[J].贵州林业科技,1996,24(4):7 -13.
[7]YE Z P.A new model for relationship between irradiance and the rate of photosynthesis in Oryza sativa[J].Photosynthetica,2007,45(4):637 -640.
[8]许大全.光合作用学[M].北京:科学出版社,2013.
[9]姜霞,张喜,丁海兵.黔中10个树种苗期水分利用效率及光合特性的差异性研究[J].西部林业科学,2013,42(5):75 -81.
[10]柴仲平,王雪梅,孙霞,等.红枣光合特性与水分利用效率日变化研究[J].西南农业学报,2010,23(1):168 -172.
[11]曹生奎,冯起,司建华,等.植物叶片水分利用效率研究综述[J].生态学报,2009,29(7):3882 -3892.
[12]金华,玉米提·哈力克,阿丽亚·拜都热拉,等.阿克苏8种常见树种叶片水分利用效率特征[J].西北林学院学报,2015,30(2):44 -50.
[13]姬亚琴,杨鹏年.不同土壤含水量条件棉花光合作用日变化特性研究[J].节水灌溉,2015(2):21 -23.
[14]杨雨华,宗建伟,杨风岭.不同生长势马尾松光合日变化研究[J].中南林业科技大学学报,2014,34(8):25 -29.
[15]于文颖,纪瑞鹏,冯锐,等.不同生育期玉米叶片光合特性及水分利用效率对水分胁迫的响应[J].生态学报,2015,35(9):1 -11.
[16]尤扬,周建,贾文庆,等.鹅掌柴叶片秋季光合特性[J].东北林业大学学报,2009,37(9):25 -26.
[17]王娈,韦小丽,张怡.不同地理种源猴樟苗期的光合特性与蒸腾特性[J].贵州农业科学,2011,39(6):39 -42.
[18]李国泰.8种园林树种光合作用特征与水分利用效率比较[J].林业科学研究,2002,15(3):291 -296.
[19]黄林敏,袁丛军,严令斌,等.不同遮荫处理对清香木苗木生长与光合速率的影响[J].浙江农业科学,2014(2):217-219.
[20]殷国兰,谭斌,杨金亮,等.3种珍贵用材树种1年生苗木光合特性研究[J].西部林业科学,2014,43(3):81 -87.
[21]陈模芳,韦小丽,张怡.紫楠实生幼苗的光合生理特性[J].贵州农业科学,2013,41(10):55-58.
猜你喜欢
节水灌溉(2021年5期)2021-05-26
中国农业科技导报(2020年12期)2020-03-15
新课程·下旬(2017年7期)2017-08-14
传奇故事(破茧成蝶)(2015年7期)2015-02-28
火花(2015年3期)2015-02-27
西北林学院学报(2015年5期)2015-01-03
音乐生活(2014年2期)2014-04-25
赤峰学院学报·自然科学版(2012年19期)2012-10-14
植物营养与肥料学报(2011年2期)2011-10-26
植物营养与肥料学报(2011年2期)2011-10-26
