
宫颈鳞癌中高表达己糖激酶2和丙酮酸激酶2与放射治疗抵抗及预后的相关性研究*
2015-12-14 08:00刘妙妙黄昕琼赵雅洁申良方
中国现代医学杂志 2015年32期
刘妙妙,黄昕琼,赵雅洁,申良方
(中南大学湘雅医院肿瘤科 湖南 长沙 410008)
·论著·
宫颈鳞癌中高表达己糖激酶2和丙酮酸激酶2与放射治疗抵抗及预后的相关性研究*
刘妙妙,黄昕琼,赵雅洁,申良方
(中南大学湘雅医院肿瘤科 湖南 长沙 410008)
目的宫颈鳞癌中高表达的己糖激酶2(HK2)和丙酮酸激酶2(PKM2)与放射治疗抵抗的相关性尚不清楚。该实验研究HK2和PKM2的表达对放射治疗抵抗及预后的意义。方法回顾性分析132例在中南大学湘雅医院肿瘤放疗科和湖南省肿瘤医院接受放射治疗的局部进展期宫颈鳞癌(LACSCC)患者的临床资料。其中放射治疗敏感患者85例,放射治疗抵抗患者47例。采用免疫组织化学法检测HK2及PKM2表达。结果放射治疗抵抗组和放射治疗敏感组的HK2高表达分别为80.9%和45.9%,差异有统计学意义(P=0.000);放射治疗抵抗组和放射治疗敏感组的PKM2高表达分别为87.2%和57.6%,差异有统计学意义(P=0.000)。根据患者的临床资料放射治疗抵抗组分为3个亚组:放射治疗未控制组、局部复发组及远处转移组。3个亚组的HK2和PKM2高表达与放射治疗敏感组比较,差异有统计学意义。HK2低表达组和高表达组的5年无进展生存率分别为80.8%和56.5%,差异有统计学意义(P=0.000)。PKM2低表达组和高表达组的5年无进展生存率分别为80.4%和60.5%,差异有统计学意义(P=0.008)。单因素分析表明,HK2、PKM2、国际妇产科协会(FIGO)分期及肿瘤大小为宫颈鳞癌无进展生存期(PFS)的独立预后指标[HRHK2=3.454,(95%CI1:1.764,6.765),PHK2=0.000;HRPKM2= 3.278,(95%CI2:1.535,7.000),PPKM2=0.002;HRFIGO分期=2.610,(95%CI3:1.453,4.689),PFIGO分期=0.001;HR肿瘤大小= 3.366,(95%CI4:1.885,6.012),P肿瘤大小=0.000]。结论高表达HK2和PKM2与放射治疗抵抗性密切相关,是LACSCC预后不良因素之一。HK2和PKM2有望成为预测放射治疗疗效和指导治疗的理想指标,值得深入研究。
局部进展期宫颈鳞癌;放射治疗抵抗;糖酵解;己糖激酶2;丙酮酸激酶2;免疫组织化学
宫颈癌占女性癌症的9%,是世界范围内女性第3大常见的恶性肿瘤。每年全球范围内有>52.9万例宫颈鳞状细胞癌患者接受放射治疗(以下简称放疗),27.5万例患者因宫颈癌死亡[1]。放疗为治疗局部晚期宫颈癌的主要手段,但放疗后复发和转移仍然是宫颈癌患者局部晚期治疗中的主要问题。因此,研究其潜在的分子机制是解决放疗抵抗的关键[2]。一些相关因素如缺氧、细胞周期、DNA损伤和修复、凋亡、生长因子、致癌基因、干细胞基因及表观遗传学等可以影响放射敏感性。有研究表明,恶性肿瘤的糖酵解代谢与放射敏感性相关[3]。最近研究发现,一些分子生物学改变在宫颈癌和宫颈癌癌前病变的进展中扮演重要角色。该异常分子可以作为生物学标志物,因其与宫颈鳞癌(squamous cell carcinoma,SCC)放射抵抗相关,也可作为预后指标[4]。早期检测放疗敏感性可以早期治疗从而改善预后。
己糖激酶2(hexokinase 2,HK2)是催化糖代谢途径的第一步,阻止葡萄糖进入细胞,对三磷酸腺苷(adenosine triphosphate,ATP)的产生起至关重要的作用[5]。笔者推测,阻断肿瘤细胞的糖酵解途径,可减少肿瘤细胞中ATP的生成,可能会阻滞肿瘤细胞的生长增殖。丙酮酸激酶2(pyruvate kinase isozyme type M2,PKM2)作为糖酵解反应最后阶段的关键酶,催化磷酸烯醇式丙酮酸(Phosphoenolpyruvate,PEP)转变为丙酮酸。PKM2通过调节糖代谢,促进肿瘤细胞的增生及能量的供给,在宫颈癌的发生、发展中发挥重要作用。为探讨糖酵解与宫颈鳞状细胞癌的相关性,本实验选取糖酵解途径中的关键限速酶HK2和PKM2,通过免疫组织化学法检测两种关键酶的表达水平,研究其与放射抵抗性的相关性和影响预后的相关因素。
1 资料与方法
1.1病例资料
本实验采用回顾性队列研究,选取2005年1月-2012年3月在中南大学湘雅医院肿瘤放疗科和湖南省肿瘤医院接受放射治疗的局部进展期宫颈鳞癌(locally advanced cervical squamous cell carcinoma,LACSCC)患者132例。纳入标准:①病理学证实为SCC;②诊断时无证据表明存在远处转移,国际妇产科协会(Federation International of Gynecology and Obstetrics,FIGO)分期为ⅠB~ⅣA);③组织切块可用于本实验;④放射治疗前未接受其他抗肿瘤治疗且放疗后未实施手术。本研究经本院伦理研究委员会批准。随访时间至2012年5月,中位随访时间为45个月(2.0~85.5个月)。中位无进展生存期(progression free survival,PFS)为43.5个月(0.0~85.5个月)。中位年龄为51岁(28~80岁)。将132例患者分为放疗敏感组(85例)和放疗抵抗组(47例)。放疗敏感组指初始治疗后3年内无局部复发和远处转移,即PFS≥36个月。放疗抵抗组指初始治疗后3年内出现放疗未控制,局部复发或远处转移,即PFS<36个月[2]。将放疗抵抗组分为3个亚组:放疗未控制组、局部复发组及远处转移组。放疗未控制组指肿瘤从未消失直至患者死亡。局部复发组指初始治疗后3年内出现局部复发;远处转移组指初始治疗后3年内出现远处转移。PFS定义为治疗结束至首次证实有局部复发或远处转移的时期。如在初始放疗后诊断为放疗未控制、局部复发或远处转移必需有临床检查、病理活检或影像学检查等证据。每个宫颈肿瘤大小的评估由临床检查直接测量,而不采用影像学手段间接测量。本实验中,放疗未控制组9例,局部复发组17例,远处转移组22例。其中1例在放疗期间出现远处转移,直至患者死亡肿块从未消失,笔者认为这1例患者既属于放疗未控制亚组也属于远处转移亚组。另有3例患者在放疗后出现远处转移,但该3例患者PFS>36个月,所以将这3例患者归为放疗敏感组。
所有患者接受放射肿瘤医师制定的外放射治疗和高剂量率的腔内放疗,外照射治疗3、4周后开始腔内放疗。A点中位总剂量为90 Gy(66~102 Gy),外照射在A点的中位剂量为46 Gy(30~52 Gy),腔内放疗在A点的中位剂量为42 Gy(20~54 Gy)。其中部分患者同时接受以铂类为基础的化疗,但化疗药物并不统一,甚至铂类药物也不统一,如顺铂、卡铂、奥沙利铂等。这些差异增加统计分析难度。
1.2免疫组织化学法检测
采用免疫组织化学法检测HK2和PKM2,将4μm的组织切片经二甲苯脱蜡处理,0.01 mol柠檬酸盐缓冲剂(pH=6.0)中微波处理(中火10 min)。冷却30 min后于磷酸盐缓冲液中浸泡,用3%过氧化氢孵育30 min以消除内源性过氧物酶活性,10%常规山羊血清(磷酸盐缓冲液稀释)中孵育30 min,滴加1∶600比例稀释的抗HK2抗体(美国Abcam公司)和1∶600比例稀释的抗PKM2抗体(美国Abcam公司),4℃过夜。按照Chem Mate试剂盒(丹麦Dako Denmark公司)和3’3-二氨基联苯胺显色试剂盒(上海碧云天生物工程有限公司)说明书进行染色。阴性对照组使用非免疫性同型抗体替代原始抗体。
1.3染色评价标准
由两位不知晓临床病理结果的病理学家对所有样本进行双盲法阅片,观察组织染色。首先通过低放大倍数(×40)观察全片HK2和PKM2的表达,再通过高放大倍数(×400)进一步确认。采用细胞计数法评价结果。阳性细胞缺乏诊断为阴性(-),观察到的阳性细胞<25%为弱阳性(±),阳性细胞数比例占25%~50%为阳性(+),观察到的阳性细胞数比例>50%为强阳性(++)[4]。根据该评估方法,染色强度(-)或(±)为低表达,染色级别(+)或(++)为高表达。
1.4统计学方法
HK2和PKM2的表达与宫颈鳞癌临床病理因素相关性分析采用χ2检验和Fisher确切概率法,放疗剂量的差异性分析用t检验。生存分析中截尾值包括随访截止及因其他原因而非宫颈鳞癌死亡的患者。根据Kaplan-Meier计算生存曲线,组间差异性用log-rank检验。采用Cox比例风险模型进行单因素和多因素生存分析。回归模型包括HK2表达(高表达与低表达)、PKM2表达(高表达与低表达)、年龄(≥50岁与<50岁)、FIGO分期(Ⅲ+Ⅳa与Ⅰb+Ⅱ)、组织病理学分级(中低级与高级别)和肿瘤直径(>4 cm与≤4 cm),P<0.05为差异有统计学意义。
2 结果
2.1临床病理学特征
132例LACSCC患者(放疗抵抗组47例,放疗敏感组85例)。其中<50岁49例,≥50岁83例;FIGO分期Ⅰb+Ⅱ和Ⅲ+Ⅳa分别为70(6+64)和62(56+6)例;按照WHO病理分级,高、中、低分化分别为10、114和8例;肿块最大径≤4 cm 79例,>4 cm 53例。放疗抵抗组与放疗敏感组的肿块大小、FIGO分期、组织病理学分级比较,差异有统计学意义(P= 0.000、0.004和0.014)。两组患者年龄、联合化疗(以铂类为基础)、A点总剂量、外照射A点剂量、腔内照射A点剂量比较,差异无统计学意义。见表1。
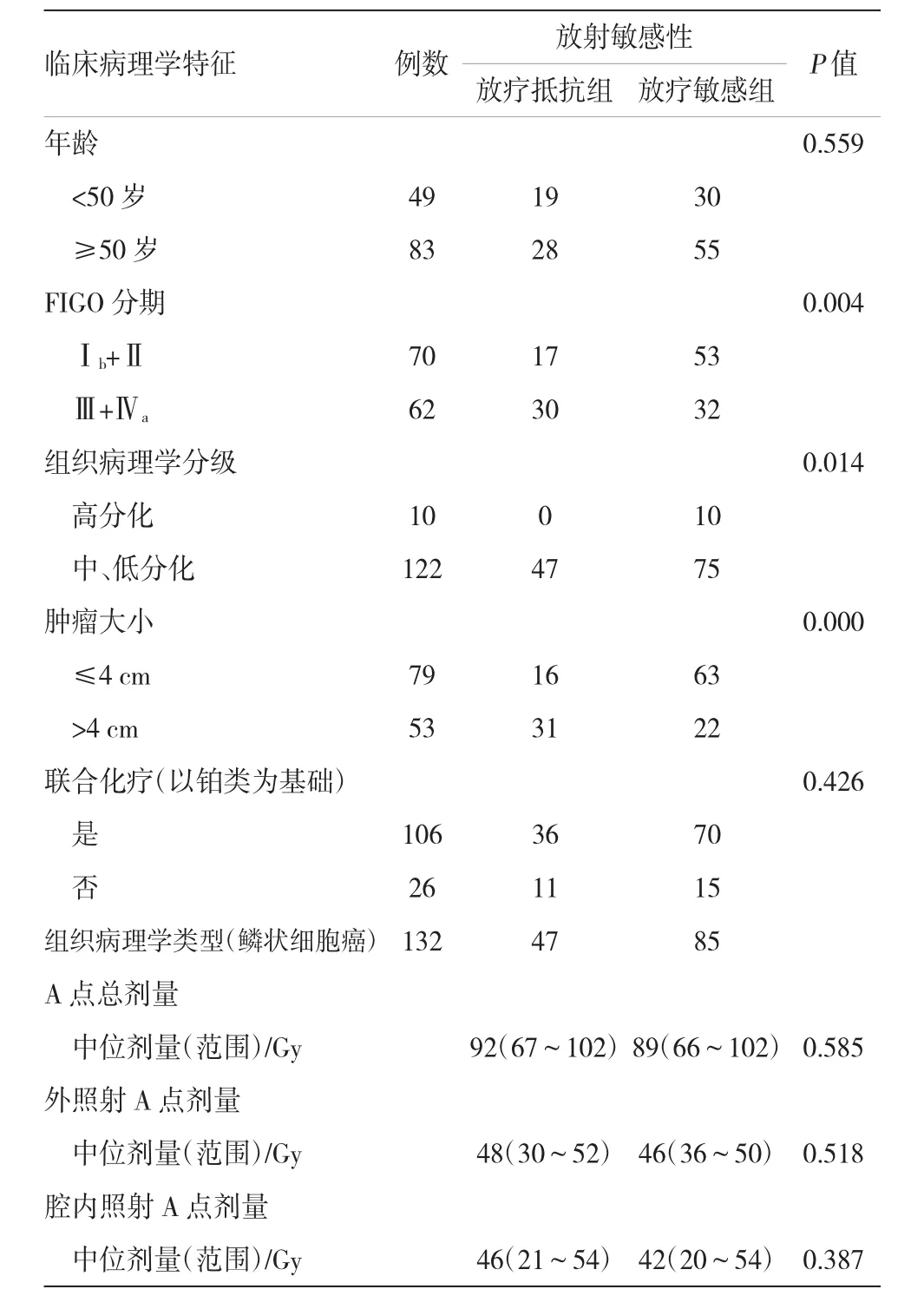
表1 临床病理学特征与放射敏感性的相关性例
2.2 HK2、PKM2表达与临床病理学特征的相关性
HK2位于宫颈癌细胞质中,放疗抵抗组较放疗敏感组染色明显(见图1)。132例宫颈癌患者中,HK2低表达55例(41.7%),高表达77例(58.3%)。结果表明HK2表达与患者年龄、FIGO分期、病理分级及肿瘤大小无关。见表2。
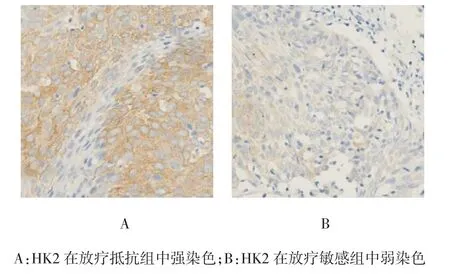
图1 HK2在放疗抵抗组和放疗敏感组中的典型染色(×400)
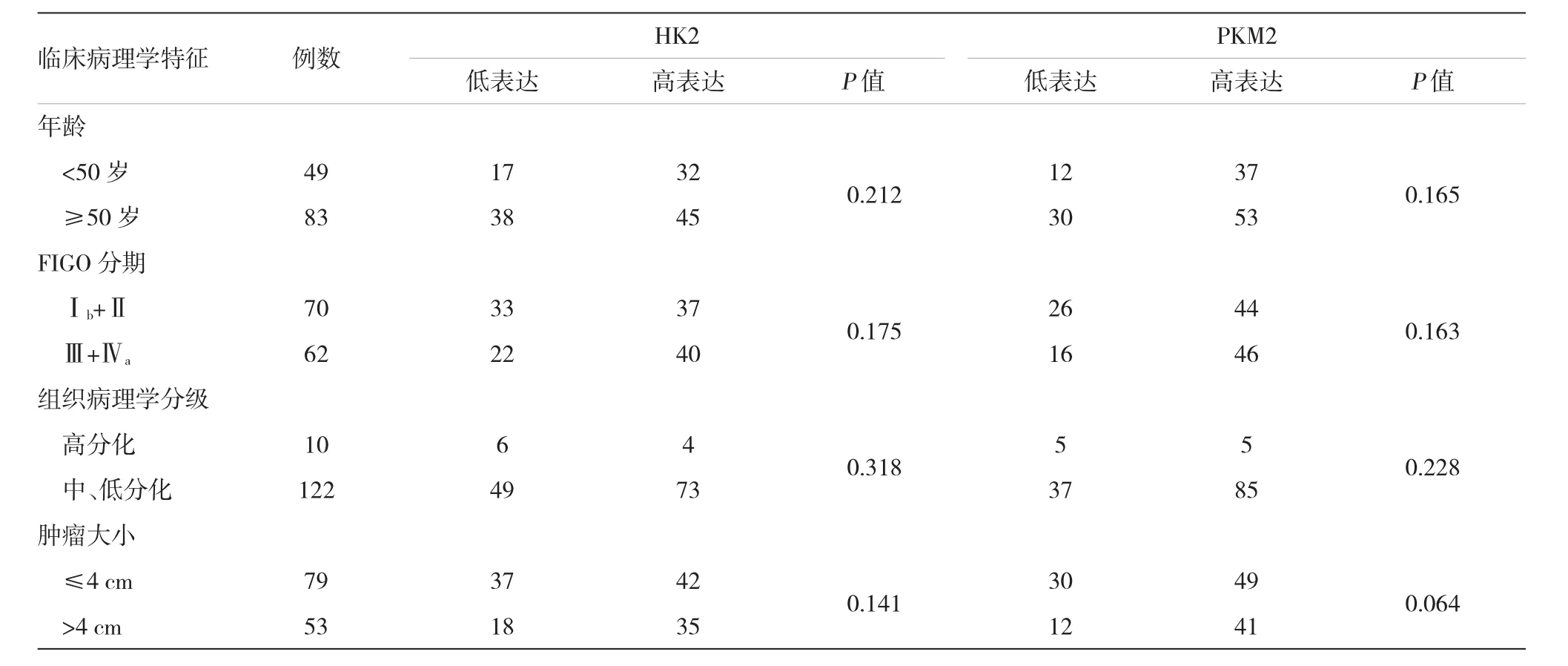
表2 宫颈癌临床病理学特征与HK2、PKM2表达的相关性例
PKM2位于宫颈癌细胞质中,放疗抵抗组较放疗敏感组染色明显(见图2)。132例宫颈癌患者中,PKM2低表达42例(31.8%),高表达90例(68.2%)。结果表明PKM2表达与患者年龄、FIGO分期、病理分级及肿瘤无关。见表2。
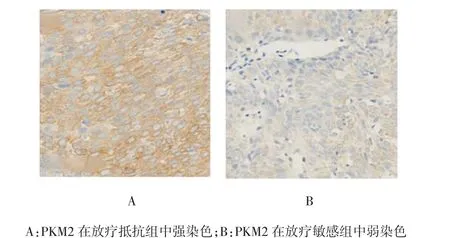
图2 PKM2在放疗抵抗组和放疗敏感组中的典型染色(×400)
2.3HK2、PKM2表达与放射敏感性的相关性
47例放疗抵抗组中,9例为HK2低表达(19.1%),38例为HK2高表达(80.9%)。85例放疗敏感组中,46例为HK2低表达(54.1%),39例为HK2高表达(45.9%)。两组HK2高表达率比较,差异有统计学意义(P=0.000)。根据患者的临床资料放疗抵抗组分为3个亚组:放疗未控制组、局部复发组及远处转移组。这3个亚组的HK2高表达率分别为:88.9%(8/9)、82.4%(14/17)和77.3%(17/22)。放疗未控制组、局部复发组及远处转移组的HK2高表达与放疗敏感组比较,差异有统计学意义(P=0.030、0.006和0.009)。见表3。
47例放疗抵抗组中,6例为PKM2低表达(14.6%),41例为PKM2高表达(87.2%)。85例放疗敏感组中,36例为PKM2低表达(42.4%),49例为PKM2高表达(57.6%)。两组PKM2高表达率比较,差异有统计学意义(P=0.000)。放疗未控制组、局部复发组及远处转移组的PKM2高表达率分别为:100.0%(9/9)、88.2%(15/17)和81.8%(18/22)。放疗未控制组、局部复发组及远处转移组的PKM2高表达与放疗敏感组比较,差异有统计学意义(P=0.012、0.026和0.048)。见表3。
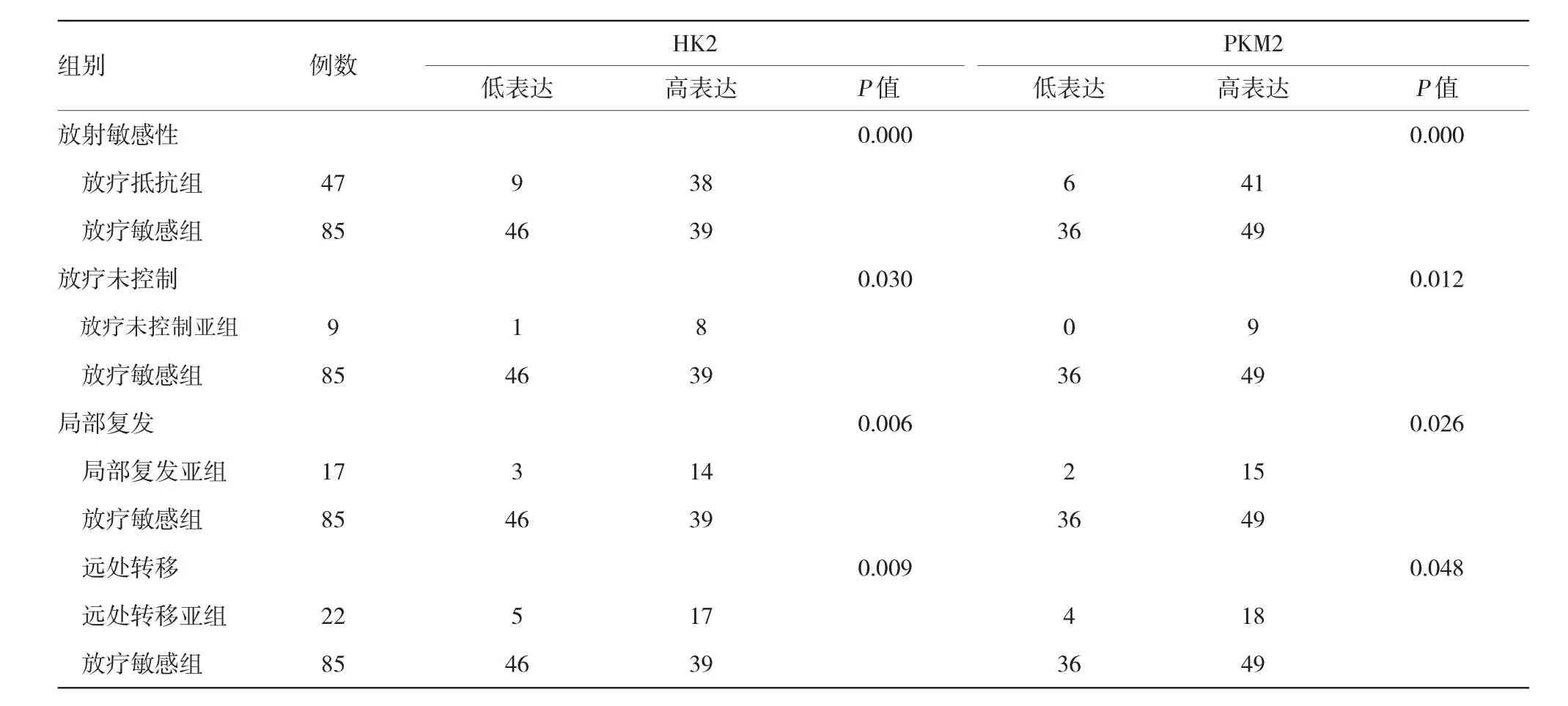
表3 宫颈癌组织放射敏感性与HK2、PKM2表达的相关性例
2.4HK2、PKM2表达与生存率的相关性
根据肿瘤的HK2表达进行分层,HK2低表达组(55例)和高表达组(77例)的5年无进展生存率分别为80.8%和56.5%。Kaplan-Meier生存分析(logrank检验)表明两组的生存率比较,差异有统计学意义(P=0.000)(见图3A)。
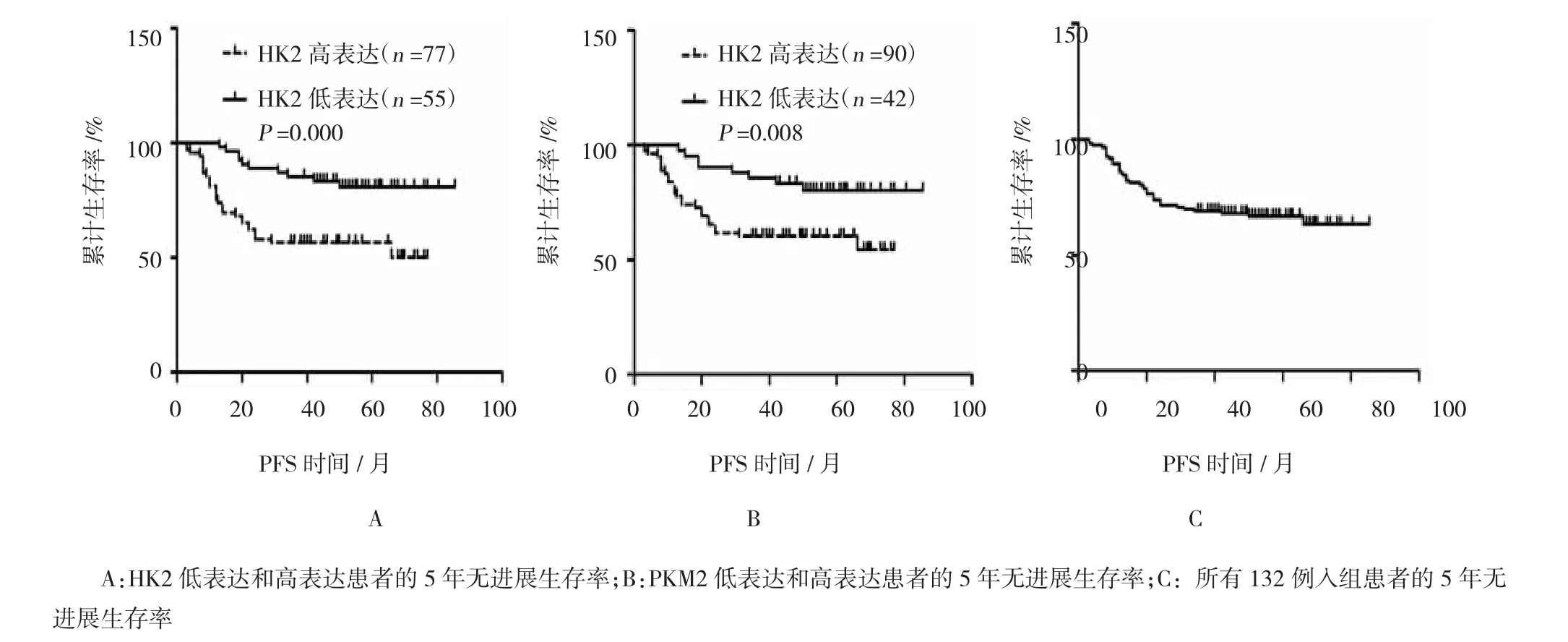
图3 Kaplan-Meier生存曲线
根据肿瘤的PKM2表达分层,PKM2低表达组(42例)和高表达组(90例)的5年无进展生存率分别为80.4%和60.5%。Kaplan-Meier生存分析(logrank检验)表明两组的生存率比较,差异有统计学意义(P=0.008)(见图3B)。
所有132例入组患者的5年无进展生存率为66.8%(见图3C)。
单因素分析表明,LACSCC患者PFS的预测因素包括HK2、PKM2、FIGO分期及肿瘤大小[HRHK2= 3.454,(95%CI1:1.764,6.765),PHK2=0.000;HRPKM2= 3.278,(95%CI2:1.535,7.000),PPKM2=0.002;HRFIGO分期= 2.610,(95%CI3:1.453,4.689),PFIGO分期=0.001;HR肿瘤大小=3.366,(95%CI4:1.885,6.012),P肿瘤大小= 0.000]。见表4。
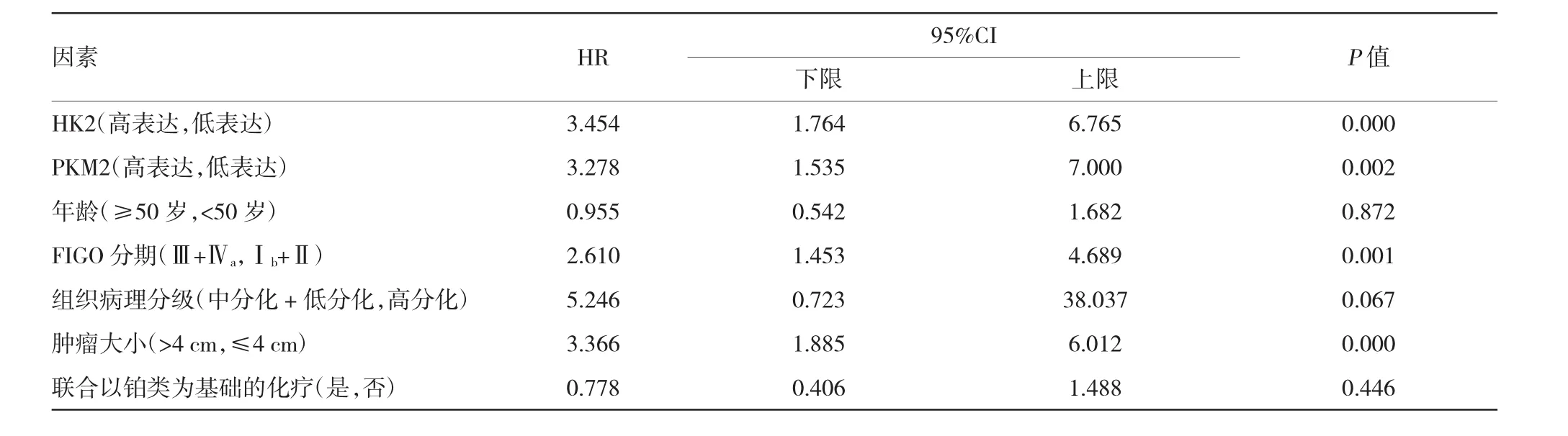
表4 PFS的单因素回归分析
3 讨论
本研究表明HK2、PKM2表达与临床病理学因素(包括年龄、FIGO分期、肿瘤大小)无关。放疗抵抗组与放疗敏感组的HK2、PKM2高表达率比较,差异有统计学意义;3个放疗抵抗亚组的HK2、PKM2高表达率与放疗敏感组比较,差异有统计学意义。笔者发现HK2和PKM2高表达患者的5年无进展生存率明显低于低表达患者,HK2和PKM2高表达提示放疗抵抗,可能导致肿瘤未控制、局部复发及远处转移。
HK2作为糖酵解途径中第一个反应限速酶,能阻止葡萄糖进入细胞,对ATP的产生起至关重要的作用[5]。笔者推测,阻断糖酵解途径可减少肿瘤细胞中ATP的生成,可能阻滞肿瘤细胞的生长和增殖。线粒体结合型HK比游离型能更有效地利用高浓度ATP[6]。在有氧环境中,肿瘤细胞中>1/2的ATP来源于糖酵解;在缺氧环境下,糖酵解比线粒体氧化磷酸化生成更多ATP,保证肿瘤细胞生长和恶性增殖[6]。有研究表明,HK2不仅保证高度恶性肿瘤细胞在不同的代谢环境中存活,同时其糖酵解产物可以影响线粒体介导的细胞凋亡[6-7]。HK2高表达同时也保护细胞免受氧化诱导的细胞死亡[8-9]。有研究证实,HK2在肺上皮细胞中表达,且HK2的表达能防止氧化诱导的细胞死亡[9]。因此笔者认为HK2在糖酵解中发挥重要作用。
PKM2是丙酮酸激酶的一个重要的同工酶,参与糖酵解反应的最后阶段,催化PEP脱磷成丙酮酸,在缺氧条件下生成ATP[10],改变糖酵解通量以满足增殖的肿瘤细胞需要[11]。与正常组织比较,PKM2在各种恶性肿瘤中表达明显增加,如肺癌、乳腺癌、前列腺癌、血液、宫颈癌、肾癌、膀胱癌及结肠癌[12]。在正常增殖细胞中,PKM2以有活性的四聚体形式存在。与此相反,与底物PEP亲和力低的无活性二聚体常存在于肿瘤细胞中,这种形式的存在导致肿瘤细胞高水平的糖代谢,与生物大分子如核酸、磷脂和氨基酸的合成密切相关,为肿瘤的增殖提供一条新的途径[13]。同时,细胞内的葡萄糖合成磷脂、核酸及氨基酸,大量的谷氨酰胺转变为乳酸并提供能量。与线粒体有氧呼吸不同的是,丙酮酸激酶能量代谢不依赖于氧气,所以无论是否缺氧,肿瘤细胞中PKM2都从四聚体转化为二聚体,这有利于肿瘤细胞的增殖[14]。CHRISTOFK等[15]发现,在糖酵解环境中,PKM1可以转化为PKM2,且体内、外实验证实PKM2的高表达可增强Warburg效应,促进肿瘤生长。STETAK等[16]发现,PKM2从细胞质进入细胞核后,会引起肿瘤细胞的凋亡。因此,笔者认为PKM2在糖酵解反应中有重要意义。
本实验结果表明,HK2、PKM2表达与LACSCC放疗抵抗密切相关。但是没有类似基于临床水平的研究证实。有研究显示,恶性肿瘤中上调的糖酵解活动与肿瘤细胞恶性程度相关,提示放疗或者化疗抵抗,预后不良[17-18]。有研究表明,放疗期间抑制糖酵解活动可增强放疗敏感性[3]。HK2和PKM2在糖酵解反应中起重要作用。因此笔者推测HK2和PKM2的高表达与放疗抵抗相关。
放疗抵抗的机制非常复杂。首先,缺氧会导致肿瘤细胞放疗抵抗[3]。当放射线直接损伤组织时产生自由基,自由基和活性氧破坏双链DNA导致细胞凋亡[19]。缺氧可以使受损的DNA恢复双链结构,因此在缺氧环境中DNA损伤程度降低[20]。BROWN等[20]认为,细胞在缺氧条件下必须增加放射剂量才能获得和需氧条件下相同的存活分数,并证明当氧分压<10 mmHg时放疗抵抗尤为明显。其次,实体肿瘤的乳酸含量与放疗抵抗相关,50%肿瘤控制剂量值与肿瘤乳酸水平呈正相关。治疗期间瞬态抑制糖酵解可能会导致肿瘤细胞放疗敏感性的增高[21],有研究发现,糖酵解产物如乳酸、丙酮酸是一种有效的游离基清除剂,可以抵消放疗放射代谢产物如活性氧产生的大量自由基,进而导致放射抵抗[22]。还有研究表明,原发肿瘤病灶的乳酸含量可以预示高转移率和低生存率[23]。在外膜上HK2与线粒体相结合,阻断促凋亡蛋白Bax和VDAC的交互作用[24],这在Hela细胞和人胚肾细胞的抗细胞凋亡机制中得到证实[24]。通过抑制VDAC与Bax、Bak的交互作用,防止线粒体通透性增加,阻止膜间隙细胞色素C的释放,从而抑制细胞凋亡[25]。近期研究表明,上诉机制可能与依赖于丝氨酸/苏氨酸激酶(Akt/PKB)的生长因子诱导的信号转导途径相关,Akt/PKB是生长因子介导细胞存活的主要下游效应器[26]。因此,HK2高表达与放疗抵抗密切相关。目前HK2作为肿瘤不良预后因子已在早期宫颈癌、胶质母细胞瘤、肝癌等多种肿瘤中得到证实[4,27-28]。
本研究显示,PKM2在多种肿瘤细胞中高表达,参与多条细胞信号转导通路,与肿瘤的发生、发展密切相关。SCHAFER等[29]在FTO2B鼠肝癌细胞的研究中发现,葡萄糖的增加引起脱磷酸转录因子SP1的数量增加,导致转录因子DNA结合活性增高,最终引起PKM2表达增加。体内、外实验证实,PKM2在肝细胞癌中发挥致癌基因作用[30]。最近研究发现,通过Akt/PI3K/哺乳动物雷帕霉素靶蛋白(mammalian target of rapamycin,mTOR)途径,PKM2通过缺氧诱导因子-1α(hypoxia inducible factor-1α,HIF-1α)/c-Myc表达上调,促进Warburg效应和肿瘤生长[31]。mTOR通过HIF-1α介导的PKM基因转录和c-Myc介导的选择性剪切,共同上调PKM2的表达,引发Warburg效应,从而促进肿瘤的发生、发展。联合抑制糖酵解和mTOR功能可协同阻碍肿瘤的发展[32]。人乳头瘤病毒16型(human papilloma virus-16,HPV-16)的E7蛋白可直接与PKM2结合,提高PKM2二聚体形式的稳定性,E7的表达可保护葡萄糖碳合成途径,减少肿瘤细胞对氧气的需要量,在细胞转化过程中发挥关键性作用[33]。其次,缺氧条件下可以通过稳定的转录因子HIF-1α上调PKM基因的表达[34],增加乏氧细胞对化疗与放疗的抵抗[35]。此外,PMK2表达影响肿瘤细胞的分化、增殖及总生存率。在宫颈浸润性癌中,PKM2表达强于宫颈上皮内瘤变[36],因此认为PKM2与宫颈癌的形成和浸润相关。越来越多的研究发现,PKM2基因敲除不仅降低肺癌细胞的糖酵解作用,而且限制肺癌细胞的增殖[15],并通过抑制Akt和磷酸肌醇依赖激酶-1(phosphoinositide-dependent kinase 1,PDK1)增强细胞外调节蛋白激酶1/2(extracellular regulated protein kinases,ERK1/2)和糖原合成激酶3β(glycogen synthesis kinase 3β,GSK3β)磷酸化,加速肿瘤细胞的自噬和凋亡,增强非小细胞肺癌(non small cell carcinoma,NSCLC)的放射敏感性[37]。所以高表达PKM2预示放疗抵抗及预后不良。
综上所述,局部晚期宫颈鳞癌患者中高表达的HK2、PKM2与放疗抵抗相关,是LACSCC预后不良因素之一。这将成为放疗抵抗分子机制上的理想治疗靶点,具有良好的临床应用前景。
[1]JEMAL A,BRAY F,CENTER MM,et al.Global cancer statistics[J].Cancer J Clin,2011,61(2):69-90.
[2]KIM MK,KIM TJ,SUNG CO,et al.Clinical significance of HIF-2 alpha immunostaining area in radioresistant cervical cancer[J].J Gynecol Oncol,2011,22(1):44-48.
[3]SATTLER UG,MEYER SS,QUENNET V,et al.Glycolytic metabolism and tumour response to fractionated irradiation[J].Radiother Oncol,2010,94(1):102-109.
[4]PENG GQ,YANG YA,ZHONG CG,et al.A study of association between expression of hOGG1,VDAC1,HK-2 and cervical carcinoma[J].J Exp Clin Cancer Res,2010,29:129.
[5]PRINTZ RL,OSAWA H,ARDEHALI H,et al.HexokinaseⅡgene:structure,regulation and promoter organization[J].Biochem Soc Trans,1997,25(1):107-112.
[6]PEDERSEN PL,MATHUPALA S,REMPEL A,et al.Mitochondrial bound typeⅡhexokinase:a key player in the growth and survival of many cancers and an ideal prospect for therapeutic intervention[J].Biochim Biophys Acta,2002,1555(1/2/3):14-20.
[7]PASTORINO JG,HOEK JB.HexokinaseⅡ:the integration of energy metabolism and control of apoptosis[J].Curr Med Chem, 2003,10(16):1535-1551.
[8]MATHUPALA SP,KO YH,PEDERSEN PL.HexokinaseⅡ: cancer's double-edged sword acting as both facilitator and gatekeeper of malignancy when bound to mitochondria[J].Oncogene, 2006,25(34):4777-4786.
[9]AHMAD A,AHMAD S,SCHNEIDER BK,et al.Elevated expression of hexokinaseⅡprotects human lung epithelial-like A549 cells against oxidative injury[J].Am J Physiol Lung Cell Mol Physiol,2002,283(3):573-584.
[10]VANDER HMG,CANTLEY LC,THOMPSON CB.Understand-ing the warburg effect:the metabolic requirements of cell proliferation[J].Science,2009,324(5930):1029-1033.
[11]IQBAL MA,GUPTA V,GOPINATH P,et al.Pyruvate kinase M2 and cancer:an updated assessment[J].FEBS Lett,2014, 588(16):2685-2692.
[12]BLUEMLEIN K,GRUNING NM,FEICHTINGER RG,et al.No evidence for a shift in pyruvate kinase PKM1 to PKM2 expression during tumorigenesis[J].Oncotarget,2011,2(5):393-400.
[13]MAZUREK S,EIGENBRODT E.The tumor metabolome[J].Anticancer Res,2003,23(2):1149-1154.
[14]MAZUREK S,ZWERSCHKE W,JANSEN-DURR P,et al. Metabolic cooperation between different oncogenes during cell transformation:interaction between activated ras and HPV-16 E7[J].Oncogene,2001,20(47):6891-6898.
[15]CHRISTOFK HR,VANDER HMG,HARRIS MH,et al.The M2 splice isoform of pyruvate kinase is important for cancer metabolism and tumour growth[J].Nature,2008,452(7184): 230-233.
[16]STETAK A,VERESS R,OVADI J,et al.Nuclear translocation of the tumor marker pyruvate kinase M2 induces programmed cell death[J].Cancer Res,2007,67(4):1602-1608.
[17]SUH DH,KIM MA,KIM H,et al.Association of overexpression of hexokinaseⅡwith chemoresistance in epithelial ovarian cancer[J].Clin Exp Med,2014,14(3):345-353.
[18]HUANG XQ,CHEN X,XIE XX,etal.Co-expressionof CD147 and GLUT-1 indicates radiation resistance and poor prognosis in cervical squamous cell carcinoma[J].Int J Clin Exp Pathol,2014,7(4):1651-1666.
[19]HOOGSTEEN IJ,MARRES HA,VAN DER KOGEL AJ,et al. The hypoxic tumour microenvironment,patient selection and hypoxia-modifying treatments[J].Clin Oncol,2007,19(6):385-396.
[20]BROWN JM,WILSON WR.Exploiting tumour hypoxia in cancer treatment[J].Nat Rev Cancer,2004,4(6):437-447.
[21]QUENNET V,YAROMINA A,ZIPS D,et al.Tumor lactate content predicts for response to fractionated irradiation of human squamous cell carcinomas in nude mice[J].Radiother Oncol,2006, 81(2):130-135.
[22]MEIJER TW,KAANDERS JH,SPAN PN,et al.Targeting hypoxia,HIF-1,and tumor glucose metabolism to improve radiotherapy efficacy[J].Clin Cancer Res,2012,18(20):5585-5594.
[23]WALENTA S,MUELLER-KLIESER WF.Lactate:mirror and motor of tumor malignancy[J].Semin Radiat Oncol,2004,14(3): 267-274.
[24]BRYSON JM,COY PE,GOTTLOB K,et al.Increased hexokinase activity,of either ectopic or endogenous origin,protects renal epithelial cells against acute oxidant-induced cell death[J]. J Biol Chem,2002,277(13):11392-11400.
[25]ARZOINE L,ZILBERBERG N,BEN-ROMANO R,et al.Voltage-dependent anion channel 1-based peptides interact with hexokinase to prevent its anti-apoptotic activity[J].J Biol Chem, 2009,284(6):3946-3955.
[26]VANDER HMG,PLAS DR,RATHMELL JC,et al.Growth factors can influence cell growth and survival through effects on glucose metabolism[J].Mol Cell Biol,2001,21(17):5899-5912.
[27]WOLF A,AGNIHOTRI S,MICALLEF J,et al.Hexokinase 2 is a key mediator of aerobic glycolysis and promotes tumor growth in human glioblastoma multiforme[J].J Exp Med,2011,208(2): 313-326.
[28]MILANE L,DUAN Z,AMIJI M.Role of hypoxia and glycolysis in the development of multi-drug resistance in human tumor cells and the establishment of an orthotopic multi-drug resistant tumor model in nude mice using hypoxic pre-conditioning[J]. Cancer Cell Int,2011,11:3.
[29]SCHAFER D,HAMM-KUNZELMANN B,BRAND K.Glucose regulates the promoter activity of aldolase A and pyruvate kinase M2 via dephosphorylation of Sp1[J].FEBS Lett,1997,417 (3):325-328.
[30]LIU WR,TIAN MX,YANG LX,et al.PKM2 promotes metastasis by recruiting myeloid-derived suppressor cells and indicates poor prognosis for hepatocellular carcinoma[J].Oncotarget, 2015,6(2):846-861.
[31]SUN Q,CHEN X,MA J,et al.Mammalian target of rapamycin up-regulation of pyruvate kinase isoenzyme type M2 is critical for aerobic glycolysis and tumor growth[J].Proc Natl Acad Sci USA,2011,108(10):4129-4134.
[32]孙倩.M2型丙酮酸激酶(PKM2)在肿瘤代谢和发生中的作用[J].基础医学与临床,2012(5):462-467.
[33]ZWERSCHKE W,MAZUREK S,MASSIMI P,et al.Modulation of type M2 pyruvate kinase activity by the human papillomavirus type 16 E7 oncoprotein[J].Proc Natl Acad Sci USA, 1999,96(4):1291-1296.
[34]KRESS S,STEIN A,MAURER P,et al.Expression of hypoxia-inducible genes in tumor cells[J].J Cancer Res Clin Oncol, 1998,124(6):315-320.
[35]LARA PC,LLORET M,CLAVO B,et al.Severe hypoxia induces chemo-resistance in clinical cervical tumors through MVP over-expression[J].Radiat Oncol,2009,4:29.
[36]YANG Y,PENG GQ,TIAN Y,et al.A study of PKM2, PFK-1,and ANT1 expressions in cervical biopsy tissues in China[J].Med Oncol,2012,29(4):2904-2910.
[37]MENG MB,WANG HH,GUO WH,et al.Targeting pyruvate kinase M2 contributes to radiosensitivity of non-small cell lung cancer cells in vitro and in vivo[J].Cancer Lett,2015,356(2): 985-993.
(申海菊 编辑)
Association of high expressions of hexokinase 2 and pyruvate kinase isozyme type M2 with radiation resistance and prognosis in cervical cancer*
Miao-miao LIU,Xin-qiong HUANG,Ya-jie ZHAO,Liang-fang SHEN
(Department of Oncology,Xiangya Hospital,Central South University, Changsha,Hunan 410008,P.R.China)
【Objective】To investigate the association of high expressions of hexokinase 2(HK2)and pyruvate kinase isozyme type M2(PKM2)with radiation resistance in locally advanced cervical squamous cell carcinoma(LACSCC)and its significance.【Methods】In this study 132 female patients who received radiotherapy for LACSCC were retrospectively analyzed,including 85 radiation-sensitive cases and 47 radiation-resistant cases.The expressions of HK2 and PKM2 were examined by immunohistochemistry.【Results】High expression rate of HK2 in the radiation-resistant and radiation-sensitive groups was 80.9%and 45.9%,respectively,and the difference was statistically significant(P=0.000).The high expression rate of PKM2 was87.2%and 57.6%in the radiation-resistant and radiation-sensitive groups respectively,and the difference was statistically significant(P=0.000).The 5-year progress-free survival(PFS)rate in the patients with low and high expression of HK2 was 80.8%and 56.5%respectively with statistical difference(P=0.000).The 5-year progress-free survival rate in the patients with low and high expression of PKM2 was 80.4%and 60.5%respectively,and the difference was statistically significant(P=0.008).Univariate analyses showed that HK2[HR= 3.454,(95%CI:1.764,6.765),P=0.000],PKM2[HR=3.278,(95%CI:1.535,7.000),P=0.002],FIGO stage [HR=2.610,(95%CI:1.453,4.689),P=0.001]and tumor diameter[HR=3.366,(95%CI:1.885,6.012),P= 0.000]were the prognostic predictors of PFS in the patients with cervical SCC.【Conclusions】Over-expressions of HK2 and PKM2 indicate LACSCC with radioresistance.Either of them can be considered as an independent poor prognostic biomarker for cervical squamous carcinoma,may become the ideal molecular therapeutic target with good clinical application prospects and is worthy of exploring.
locally advanced cervical squamous cell carcinoma;radiation resistance;glycolysis;hexokinase 2;pyruvate kinase isozyme type M2;immunohistochemistry
R737.33
A
1005-8982(2015)32-0001-08
2015-07-23
国家自然科学基金(No:81372792);湖南省科技厅重点项目(No:2013sk2019);湖南省自然科学基金(No:2015JJ4055)
申良方,E-mail:slf1688@sina.com;Tel:13975805137
猜你喜欢
核安全(2022年3期)2022-06-29
天津医科大学学报(2021年3期)2021-07-21
世界最新医学信息文摘(2021年25期)2021-05-13
世界科学技术-中医药现代化(2021年12期)2021-04-19
健康必读·下旬刊(2020年2期)2020-03-19
中国循证心血管医学杂志(2020年11期)2020-01-08
中成药(2019年12期)2020-01-04
中国组织化学与细胞化学杂志(2017年1期)2017-06-15
中华老年口腔医学杂志(2016年4期)2017-01-15
中国医药生物技术(2015年4期)2015-12-26
