
微小RNA-146a-5p在新生隐球菌、脂多糖诱导THP-1细胞中表达的对比分析
2015-12-02 01:59:00廖宁馨陈江汉
中国感染与化疗杂志 2015年6期
陈 欢,陈 宏,廖宁馨,陈江汉
中枢神经系统真菌感染中以隐球菌性脑膜炎(隐脑)最为常见,其致病菌为新生隐球菌和格特隐球菌,我国以新生隐球菌感染为主,格特隐球菌感染少见。流行病学显示我国的大部分隐球菌病患者发生在“免疫功能正常”的人群,患有人类免疫缺陷综合征(AIDS)/人类免疫缺陷病毒感染的患者仅占六分之一[1],这类“免疫功能正常”隐脑患者临床治疗困难,治疗不良反应大,耗时长,病死率高,预后较差[2]。随着高效抗反转录病毒治疗的应用,艾滋病相关隐脑的发病率已显著下降,而非艾滋病相关隐脑的发病率却呈逐年上升趋势[3]。探索“免疫功能正常”隐脑患者内在的发病机制,对这类患者的临床治疗显得尤为重要。我们通过观察并分析微小RNA-146a-5p(miR-146a-5p)在新生隐球菌、脂多糖(LPS)刺激THP-1细胞中表达的差异,为探讨miR-146a-5p在新生隐球菌所致隐脑中的调控作用及机制奠定基础。
1 材料与方法
1.1 材料
人单核细胞系THP-1购于中国科学院细胞库,新生隐球菌(标准株WM148)来源于上海长征医院皮肤真菌保藏实验室,LPS 购自美国Sigma 公司;RPMI-1640培养基、胎牛血清购自美国Gibco 公司,人肿瘤坏死因子(TNF)-α、白细胞介素(IL)-6购于美国ebioscience;TRIzol购于美国Invitrogen;TaqMan MicroRNA Reverse Transcription Kit、TaqMan Universal PCR Master Mix、Taq-Man MicroRNA Assays购于大连TaKaRa公司);氯仿、异丙醇及无水乙醇等均为国产分析纯。
1.2 方法
1.2.1 细胞培养 人单核细胞系THP-1培养于含10%的胎牛血清、100 u/mL 青霉素、100 u/mL 链霉素,细胞置于37℃、5% CO2、饱和湿度的培养箱中培养;每2~3天更换培养液1次,细胞密度适度时常规传代培养。
1.2.2 细胞处理及分组 将THP-1细胞按1×106个/mL接种于六孔板,每孔2 mL。实验分为2组:①新生隐球菌诱导组,按照数量1∶1比例将灭活新生隐球菌与THP-1细胞同培养;②LPS刺激组:培养基中加入LPS(终质量浓度1 mg/L)。分别培养0(空白对照)、3、6、9、12 h后,离心收集各时点培养上清液和沉淀细胞,于-80℃保存,用于酶联免疫吸附测定(ELISA)检测和TRIzol法提取总RNA。
1.2.3 实时荧 光定量 聚合酶 链反应(Real-time PCR)测定miR-146a-5p的表达 ①总RNA 抽提。采用TRIzol法提取沉淀细胞中的总RNA。通过琼脂糖凝胶电泳检测总RNA 质量,紫外分光光度计测定总RNA 的水平;②反转录。采用TaKaRa公司Prime ScriptTMRT reagent Kit(Perfect Real Time)特异miRNA 的茎环引物。所有操作均在冰上完成。10μL 的反应体系包括:2μL 5×Prime Script Buffer,0.5μL Prime Script RT Enzyme mixⅠ,0.5μL Gene specific primer,free water组成的master mix。反应条件:42℃15 min,85℃5 s;4℃;③Real-time PCR。采用SYBR®Premix Ex TaqTM II(Tli RNaseH Plus)检测试剂盒。20μL的反应体系包括:10μL SYBR®Premix Ex Taq II(Tli RNaseH Plus)(2×),0.8μL的PCR Forward Primer(10 μmol/L),0.8 μL 的PCR Reverse Primer(10μmol/L),2μL DNA 模板(<100 ng),2μL dH2O(灭菌蒸馏水)。在ABI7300定量PCR仪上扩增和检测,反应条件:95℃30 s,40个扩增循环(95℃5 s,60℃30 s)。选取使用U6作为内参照,同时进行PCR,获得Ct值后,应用比较Ct法进行相对定量,miR-146a-5p 的相对 表达量采用2-ΔΔCt计 算。PCR引物由上海生工生物有限公司合成,见表1。
1.2.4 ELISA 检测培养上清液中的TNF-α、IL-6含量 分别按照美国Ebioscience公司的TNF-α、IL-6 ELISA 试剂盒使用说明书步骤进行操作,于450 nm 处测量吸光值。根据标准品光密度值,对应相应稀释度,以线性回归方程计算相应培养液细胞因子TNF-α、IL-6的水平。
1.2.5 统计方法 所有的统计处理均采用SPSS 18.0统计软件进行分析,多样本均数的比较采用单因素方差分析,组间两两比较采用Dunnett-t检验,P<0.05为差异有统计学意义。
2 结果
2.1 miR-146a-5p表达
新生隐球菌诱导THP-1细胞,3、6、9、12 h后,细胞中miR-146a-5p较0h组明显升高,3 h达到最高值,随后逐渐下降(P<0.05)。LPS诱导THP-1细胞3、6、9、12 h后,细胞中表达miR-146a-5p逐渐升高,12 h达到最高值;与0h组相比,差异有统计学意义(P<0.05),见图1。
2.2 TNF-a、IL-6表达
新生隐球菌诱导THP-1 细胞,3、6、9、12 h 后上清液中TNF-α 的表达逐渐升高,12 h 达到最高值,与0h比较差异有统计学意义(P<0.05);LPS诱导THP-1细胞3、6、9、12 h上清液中TNF-α 的表达3 h后一直处于峰值,相对0h差异有统计学意义(P<0.05),见图2-1。
新生隐球菌诱 导THP-1 细胞以 后,IL-6 的 表达各时间点变化不明显,差异无统计学意义(P>0.05);在LPS诱导后,3、6、9、12 h上清液中IL-6的表达逐渐增加,12 h达到最高值,与0h组相比差异有统计学意义(P<0.05),见图2-2。
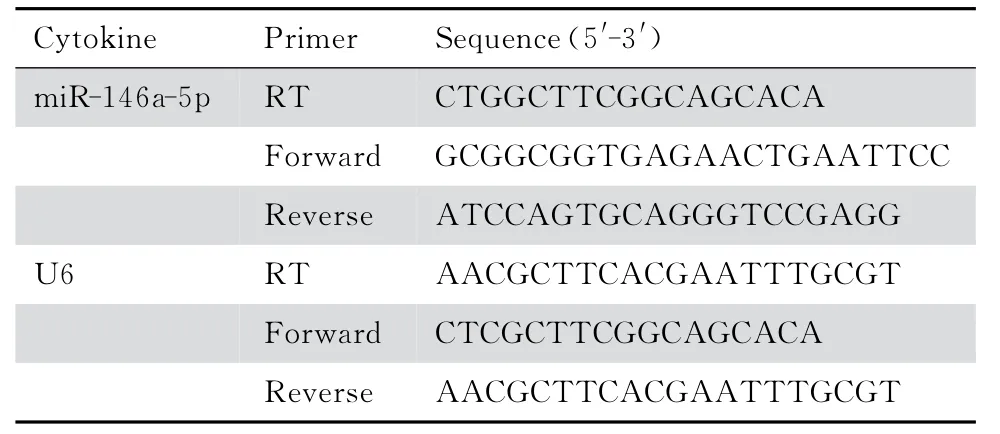
表1 Real-time PCR引物信息表Table 1 Primers used in real-time polymerase chain reaction
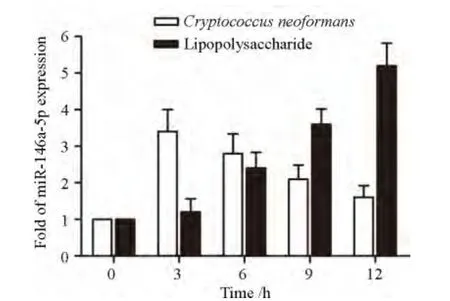
图1 miR-146a-5p在新生隐球菌、LPS不同诱导时间THP-1细胞中表达水平Figure 1 Expression of miR-146a-5p in THP-1 cells at different time points following stimulation with Cryptococcus neoformans or lipopolysaccharide
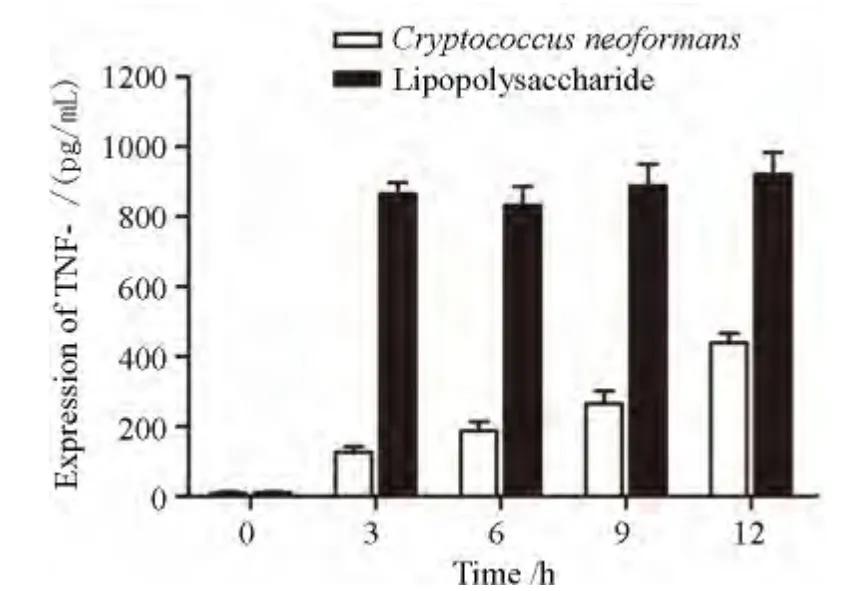
图2-1 TNF-a在新生隐球菌、LPS不同诱导时间THP-1细胞中表达水平Figure 2 -1 Expression of TNF-αin THP-1 cells at different time points following stimulation with Cryptococcus neoformans or lipopolysaccharide
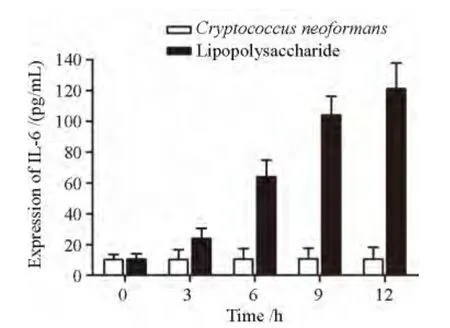
图2-2 IL-6在新生隐球菌、LPS不同诱导时间THP-1细胞中表达水平Figure 2 -2 Expression of IL-6 in THP-1 cells at different time points following stimulation with Cryptococcus neoformans or lipopolysaccharide
3 讨论
来源于单核细胞的巨噬细胞,是机体抵御病原体入侵的第一道防线,同时其介导的天然免疫反应还调控后续的适应性免疫的程度和类型,在抗感染免疫反应中起重要作用。研究表明,巨噬细胞等吞噬细胞在抗真菌感染的过程中具有重要作用,在隐球菌免疫逃逸的过程中也发挥重要影响[4-5]。经研究,白血病患者的单核细胞(THP-1)具有单核细胞和巨噬细胞的功能和生理学特性,被广泛用于毒理学、免疫学和动脉粥样硬化等研究领域[6],因此,本研究将其作为研究对象。
miRNA 是在转录后水平上调节基因表达的一组非编码小RNA 分子,广泛存在于病毒、线虫、植物及动物体内。miRNA 表达具有时空特异性,在生物体生长、发育和功能表达过程中发挥十分重要的作用,且能调控许多疾病相关基因的表达[7]。miR-146a作为miRNA 的一种,在调节固有免疫、炎性反应、病毒感染以及一些人类疾病中都发挥着重要的作用。即这些疾病的发病机制可能与miR-146a表达的变化相关[8-10]。
LPS为革兰阴性细菌外壁层中的主要成分,也是全身炎性反应的重要触发剂,可以与体内多种蛋白结合,引起各种炎性效应。它可以通过单核巨噬细胞表面的Toll样受体4(TLR4)来活化核因子κB(NFκB),从而诱导大量包括TNF-α在内的前炎性介质和细胞因子的表达[11]。Taganov 等[12]用LPS刺激人单核细胞系THP-1 后,用芯片技术检测了200 个成熟miRNA 的表达,发 现3 个miRNA(miR-132,miR-146a,miR-155)表达水平升高。进一步研究miR-146a对其他TLR 配体和一些细胞因子刺激后的反应,结果显示,TLR2、TLR4 和TLR5的配体可以明显刺激miR-146a表达上调。
我们通过研究miR-146a-5p 在新生隐球菌及LPS分别诱导THP-1 细胞炎性反应中的表达差异,同时对比分析炎性因子的变化情况,初步探讨miR-146a-5p在新生隐球菌所致隐脑中的调控作用及机制。实验结果发现,新生隐球菌刺激THP-1细胞后,miR-146a-5p的表达3 h达到峰值,LPS刺激后miR-146a-5p表达逐步升高,在12 h测量时间点达到高峰。我们推测可能与灭活的新生隐球菌含有多种免疫细胞识别的主要靶分子病原相关分子模式(PAMPs),被与之相对应的受体——模式识别受体(RRPs)识别,迅速启动免疫应答,同时也启动miRNA 调控基因的表达有关。
不同研究还发现,刺激THP-1细胞之后,新生隐球菌诱导组TNF-α表达逐步升高,IL-6无明显表达(P>0.05);LPS诱导组TNF-α在3 h的测量时间点达到峰值,与Wong 等[11]的研究结果一致,IL-6的表达逐步升高,在12 h 测量时间点达到峰值,与Ren等[13]的研究结果相符。结果表明,新生隐球菌刺激THP-1细胞之后,炎性因子的表达量和表达规律与LPS诱导组有明显差别。从miR-146a-5p的表达来看,新生隐球菌诱导后,miR-146a-5p表达迅速增高,相应炎性因子TNF-α表达缓慢、IL-6的表达不明显;LPS诱导后,miR-146a-5p表达缓慢增高,对相应的炎性因子TNF-α、IL-6 的影响逐步表现出来。
综上所述,miR-146a在新生隐球菌和LPS 诱导单核细胞炎性反应中都发挥重要的作用,miR-146a的各种病原菌诱导的免疫反应中的负调控作用得到进一步证实。因此,本实验表明:新生隐球菌诱导单核细胞的炎性反应机制与LPS 不同,miR-146a在调控过程中具有重要作用,具体作用机制有待进一步深入研究。
[1]Yuchong C,Fubin C,Jianghan C,et al.Cryptococcosis in China(1985-2010):review of cases from Chinese database[J].Mycopathologia,2012,173(5-6):329-335.
[2]Chen YY,Lai CH.Nationwide population-basedepidemiologic study of cryptococcal meningitis in Taiwan[J].Neuroepidemiology,2011,36(2):79-84.
[3]Lui G,Lee N,Ip M,et al.Cryptococcosis in apparently immunocompetent patients[J].QJM,2006,99(3):143-151.
[4]Charlier C,Nielsen K,Daou S,et al.Evidence of a role for monocytes in dissemination and brain invasion by Cryptococcus neoformans[J].Infect Immun,2009,77(1):120-127.
[5]Balloy V,Chignard M.The innate immune response to Aspergillus fumigatus[J].Microbes Infect,2009,11(12):919-927.
[6]Auwerx J.The human leukemia cell line,THP-1:a multifacetted model for the study of monocyte-macrophage differentiation[J].Experientia,1991,47(1):22-31.
[7]Lee HM,Nguyen DT,Lu LF.Progress and challenge of microRNA research in immunity[J].Front Genet,2014,5:178.
[8]Silvestri P,Di Russo C,Rigattieri S,et al.MicroRNAs and ischemic heart disease:towards a better comprehension of pathogenesis,new diagnostic tools and new therapeutic targets[J].Recent Pat Cardiovasc Drug Discov,2009,4(2):109-118.
[9]Dai R,Phillips RA,Zhang Y,et al.Suppression of LPSinduced interferon-gamma and nitric oxide in splenic lymphocytes by select estrogen-regulated microRNAs:a novel mechanism of immune modulation[J].Blood,2008,112(12):4591-4597.
[10]El Gazzar M,Church A,Liu T,et al.MicroRNA-146a regulates both transcription silencing and translation disruption of TNF-α during TLR4-induced gene reprogramming[J].J Leukoc Biology,2011,90(3):509-519.
[11]Wong LY,Cheung BM,Li YY,et al.Adrenomedullin is both proinflammatory and antiinflammatory:its effects on gene expression and secretion of cytokines and macrophage migration inhibitory factor in NR8383 macrophage cell line[J].Endocrinology,2005,146(3):1321-1327.
[12]Taganov KD,Boldin MP,Chang KJ,et al.NF-kappaBdependent induction of microRNA miR-146,an inhibitor targeted to signaling proteins of innate immune responses[J].Proc Natl Acad Sci U S A,2006,103(33):12481-12486.
[13]Ren W,Hu L,Hua F,et al.Myeloid differentiation protein 2 silencing decreases LPS-induced cytokine production and TLR4/MyD88 pathway activity in alveolar macrophages[J].Immunol Lett,2011,141(1):94-101.
猜你喜欢
天津农学院学报(2024年1期)2024-04-07 11:20:56
天津医科大学学报(2021年3期)2021-07-21 09:03:48
国际呼吸杂志(2019年1期)2019-01-28 09:37:06
现代检验医学杂志(2016年2期)2016-11-14 02:38:02
环境科技(2016年3期)2016-11-08 12:14:10
医学研究杂志(2015年9期)2015-07-01 17:28:19
温州医科大学学报(2014年7期)2014-07-18 02:43:14
现代检验医学杂志(2014年5期)2014-02-02 02:51:34
食品科学(2013年15期)2013-03-11 18:25:40
中国病理生理杂志(2012年5期)2012-01-26 04:39:37
