
加工方式对花生致敏性的影响及其致敏性评价研究进展
2015-10-29 02:47薛文通
食品科学 2015年23期
饶 欢,田 阳,陶 莎,薛文通*
(中国农业大学食品科学与营养工程学院,北京 100083)
加工方式对花生致敏性的影响及其致敏性评价研究进展
饶欢,田阳,陶 莎,薛文通*
(中国农业大学食品科学与营养工程学院,北京100083)
花生蛋白是一种理想的食品加工原料,同时也是一种致死率较高的食物过敏原,严重危害人类健康。本文主要综述食品加工方式对花生过敏蛋白致敏性的影响,以及体内、体外致敏性评价方法,为低致敏或无致敏花生制品的开发提供参考和指导。
加工;花生过敏源;致敏性评价
食物过敏是过敏性疾病的一种,在欧美等发达国家,食物过敏影响着约6%~8%的儿童和4%的成年人,中国也有约5%的儿童和2%的成年人深受其害,且其发病率在过去的几十年中呈明显上升趋势,成为全球关注的公共卫生问题[1-4]。常见致敏食品有牛奶、小麦、鸡蛋、黄豆、花生、坚果、鱼以及贝类。其中,花生过敏症是一种严重的免疫球蛋白E(immune globulin E,IgE)介导的Ⅰ型超敏反应,过敏个体暴露于微量花生过敏原中即可导致致命危害。花生过敏症通常是伴随终生,且目前没有治疗花生过敏的有效医疗手段,因此通过食品加工控制食物过敏原并为食物过敏患者提供安全的食品是食品工业的重要任务之一。
花生在不同加工过程中会发生各种复杂的物理化学变化,可不同程度地改变花生过敏原的含量和结构等,增加或降低花生致敏性。同时,花生过敏原的改变也会影响其消化时的分解方式,以及吸收时穿过肠黏膜屏障和呈递给免疫系统的形式。食物基质对引发过敏反应亦有很大影响。本文结合文献报道,在体内、体外以及分子水平综合揭示各种改性加工方法对花生致敏性的影响规律,为今后开发安全有效的花生脱敏方法奠定理论基础。
1 加工方式对花生致敏性的影响
1.1热加工
1.1.1加热方式对花生致敏蛋白的影响
热加工是食品加工中常见的方法,包括烘焙、蒸煮、烧烤、干燥等。热加工可导致某些食物过敏蛋白三维构象表位变化,使得其与人IgE结合活性降低,或者由于一些过敏蛋白非常不稳定,经过热处理后使其致敏性丧失;然而热加工亦可引起蛋白致敏性新表位的产生,或者之前隐藏的蛋白表位暴露,增强人体免疫系统对过敏原蛋白的识别和反应,而导致食物蛋白致敏效力增大。研究表明,花生中主要致敏蛋白Ara h1、Ara h2和Ara h6属于热稳定性蛋白,其结构稳定,加工过程对它们的影响也较小,热加工还可能增加其致敏性[5]。Kroghsbo等[6]分别喂饲棕色挪威(Brown Norway,BN)大鼠烘烤花生、花生酱和脱皮花生,观察大鼠过敏反应。结果发现烘烤花生并没有比脱皮花生具有更高的敏化能力。
为进一步研究热加工对花生致敏性的影响,许多学者对单一过敏原结构、致敏性及消化特性进行研究,因为致敏蛋白质在各种理化反应后,其可溶性和稳定性变差,不利于血清学和临床分析,从而加大了对食物中致敏蛋白结构及致敏性研究的难度。英国学者研究了不同热加工方式对花生过敏原Ara h1结构及过敏原性的影响,发现水煮后的Ara h1发生聚合和水解反应,形成复杂分支状结构,导致IgE结合能力和细胞因子释放能力下降。而商品化烘烤花生保留了大量β-折叠结构,保持了蛋白的过敏原性,从而使其致敏性高于水煮花生[7]。但是,Vissers等[8]研究发现Ara h2/6经145 ℃烘烤20 min后,其IgE结合能力和脱颗粒能力比未加工的花生有明显降低。因此,对加工方式降低花生致敏性的研究,还需综合考虑加工过程中多重因素与多种变应原的关系,以期达到最大程度降低致敏性,甚至消除致敏性的目的。
1.1.2食品基质对花生过敏蛋白的影响
食物加工过程中另一个有待阐明的方面是食物基质对过敏原免疫原性的影响。食物基质包括外界溶液体系以及花生本身含有的脂肪、碳水化合物和其他非致敏蛋白质等营养成分,均有可能影响到蛋白质的致敏潜力[9]。例如在食品热加工过程中最重要的非酶促化学变化——美拉德反应。过敏原氨基酸与糖相互作用产生褐色物质及特殊香气使食品增强感官性状,同时它也可能影响食物变应原致敏性。研究发现美拉德反应终产物与IgE有更高的亲和性,是导致烘烤花生致敏性高于生花生的原因之一[10]。Blanc等[7]利用花生主要致敏蛋白Ara h1与葡萄糖反应,模拟在热加工过程中发生的美拉德反应,发现此过程产生大量的果糖胺及大分子聚合物,致使IgE结合能力和β-氨基己糖苷酶释放能力降低。但与烘烤花生发生的美拉德反应比对发现,实际反应中并未产生大量果糖胺,作者猜测可能是花生中其他成分如脂肪等参与了此复杂反应过程。
花生是高脂肪含量食品,脂肪含量能达到50%[11]。有研究表明,脂肪可在消化过程中保护蛋白质,还可影响免疫细胞功能,导致蛋白质致敏性的激活和增强[12]。脂肪还有可能影响美拉德反应产物的生成量及生成速率。总体来说脂肪对花生致敏性影响的研究较少,大部分实验采用脱脂花生进行致敏研究,排除了脂肪在其中可能发挥的作用,而少数实验提出了脂肪可能对致敏性产生影响,但其具体机制还有待进一步研究[7]。
1.2非热加工
非热加工技术是食品加工过程中另一大类技术,包括高压、辐射、脉冲磁场、冻融等物理技术[13]。与传统热加工技术相比,食品非热加工的成本更低,货架期也有可能更长,可以满足消费者对食品的新鲜、营养、安全及功能性的需求。其作用除了杀菌、钝酶外,还能对食品中大分子成分进行降解,在提高食品可消化性的同时改变过敏原的结构,降低其致敏性。
辐照及超高压微射流分别处理花生过敏原Ara h2和Ara h6后,发现其各类型二级结构及三级结构发生改变,使蛋白结构由折叠变为展开,疏水性增强,致敏蛋白含量减少,抗原性随处理强度的增加而降低。此外,辐照处理所引起的Ara h6结构及抗原性变化比超高压微射流处理所引起的变化更加显著,而超高压处理在降低致敏性的同时不影响其他成分功能性质,也同样显示出其控制食物过敏原致敏性的强大优势[14-15]。许舒婷等[16]研究了电子束辐照降低花生过敏原免疫原性的效果及辐照对花生生化性质的影响,发现电子束辐照改变了过敏原的生化性质,降低了其免疫原性,且辐照对液体状态下过敏原的影响更显著。Chung等[17]分别对花生提取蛋白及花生酱进行4 min脉冲紫外线处理,发现脉冲紫外线能有效地减少63 kD致敏蛋白含量及IgE结合能力,对18~20 kD致敏蛋白没有显著影响。但随着处理时间延长和光强度的增加,Ara h2的致敏性也大大降低。Yang等[18]的实验结果也同样证实了脉冲紫外线可以有效降低花生蛋白致敏性,且对蛋白无特异性影响,但无法达到完全消除的效果。虽然各种方法都可以对过敏原性产生影响,但每种方法都有其局限性,所以各加工方法之间的联用技术也成为今后一个新的研究发展方向。
1.3酶法处理
与物理化学改性方法比较,生物改性方法具有毒副作用小、专一性强、效率高等优点。王烁等[19]通过免疫印迹法检测花生致敏蛋白含量,研究物理改性、化学改性及酶法处理对花生蛋白致敏性的影响,结果表明物理方法及化学方法虽然能在一定程度上降低致敏性,但效率不高,而经转谷氨酰胺酶处理后的致敏蛋白含量及致敏性随加酶量的增加而降低。目前,关于酶对过敏原的研究主要集中在两个方面:一是加工过程中添加单一或复合酶制剂,通过对过敏蛋白交联或降解以降低致敏性;二是研究人体消化吸收过程中酶体系对过敏原致敏性的影响机制。
花生加工过程中添加酶制剂而减低致敏性的实质主要是蛋白质酶法交联原理。利用一种或多种酶催化蛋白质内部或蛋白质分子间形成共价键而发生交联反应,改变了蛋白质的空间结构,减少致敏原含量,从而使花生致敏性有所变化。目前,谷氨酰胺转胺酶(transglutaminase,TG)、过氧化物酶(peroxidase,POD)、多酚氧化酶(polyp henol oxidase,PPO)都可用于蛋白质酶法交联。Chung等[20]用POD处理焙烤花生60 min后,其过敏原性显著降低,甚至低于生花生的过敏性。这是由于烘烤花生暴露在外的酪氨酸残基在酶催化作用下发生交联反应,从而减少了花生中过敏原的含量;作者又用PPO催化交联花生中两种主要过敏原Ara h1和Ara h2,同样取得良好抗敏效果[21]。
为研究过敏原在体内消化吸收过程中的变化机制,通常采用体外酶系模拟消化系统的研究手段,其实质主要是酶法降解[22]。花生致敏蛋白在酶系作用下分解成小肽段,破坏原有蛋白表位或生成新表位,从而影响蛋白致敏原性。Burks等[23]模拟胃液和十二指肠液(胰蛋白酶、胰凝乳蛋白酶和肠黏膜肽酶)对烘烤的花生蛋白进行连续的酶解反应,探究其经消化后的致敏性。Hong等[24]发现花生过敏蛋白经胃蛋白酶和胰蛋白酶水解后,破坏了IgE结合表位,但仍然存在可激活T细胞活性的表位。Koppelman等[25]模拟胃液对花生过敏原的研究中发现,Ara h1、Ara h3能迅速被胃蛋白酶消化,与此相反,Ara h2、Ara h6中存在的二硫键则使其可以耐受高浓度的胃蛋白酶,保持相对稳定的状态。何伟逸等[26]也证实胃蛋白酶与胰蛋白酶对花生蛋白大分子的消化能力更强。此外,通过重组表达经点突变的花生过敏原,进行花生过敏的低致敏性研究取得了很大进展。
2 花生致敏性评价
致敏性评价是衡量脱敏效果好坏的评判标准,只有标准准确、统一才能使结果更具科学性、严谨性,而目前国内对花生过敏动物模型和细胞模型以及其评价体系的研究比较缺乏,国外文献中涉及的模型纷繁复杂,无从选择,因此通过对评价方法的总结和分析可以方便找到合适的评价模型。
2.1特异性抗体评价
过敏原与IgE结合是过敏原发挥生物活性的中心环节,因此测定过敏原特异性IgE抗体在过敏症诊断及过敏原评价中具有重要地位[27]。其中酶联免疫吸附实验和免疫印迹实验是最常用的方法[28]。此外,研究显示人和小鼠因超敏反应引起的死亡还与IgG类抗体有关,因此检测食物过敏而产生的特异性IgG抗体可辅助诊断食物过敏[29]。用花生过敏患者血清进行过敏原性评价的结果比较准确,但我国花生过敏患者血清的来源有限,从而限制了该方法的推广。
2.2细胞学评价
细胞学评价常应用于花生致敏程度监测及反应机制研究。该方法从细胞生物学的角度在来监测花生致敏蛋白与免疫细胞结合引起效应细胞免疫应答、细胞脱颗粒能力、组胺释放能力以及细胞因子分泌情况等。参与花生致敏过程的免疫细胞主要有树突细胞、T细胞、肥大细胞、嗜碱性粒细胞等。血液或动物来源的原代细胞,虽然更接近人体状况,但无论从获取还是培养角度都难以适应实验过程。因此,从人或鼠身上分离、克隆的人系或鼠系传代细胞株,如RBL细胞系被广泛应用于花生致敏性评价[30]。近些年,使用外周血单个核细胞模型进行T细胞极化分析成为研究热点,此方法可在不同时间阶段测定特定细胞的应答反应。表1总结了几种典型评价花生致敏性的细胞模型。
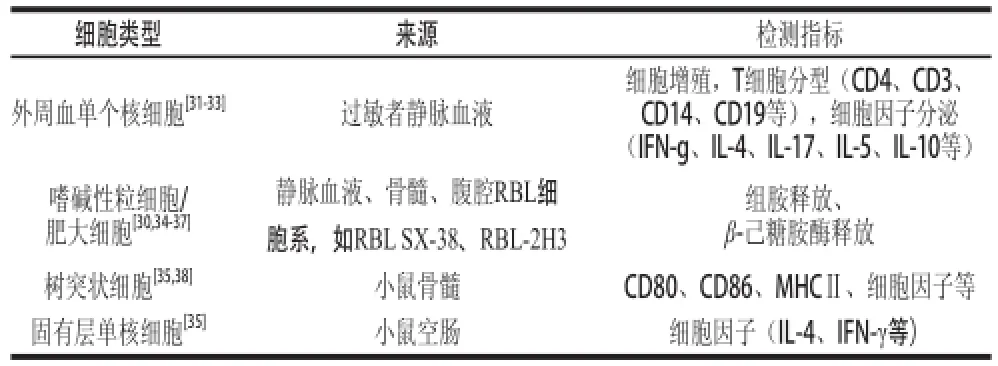
表1 几种典型评价花生致敏性的细胞模型Table 1 List of cell models for the evaluation of peanut sensitization
2.3致敏动物评价
对人体内过敏反应的研究,由于要考虑伦理及安全性,不适合大规模开展,特别是对花生此类高致死性过敏原的研究。体外实验大量依赖患者血清,并且血清IgE也不完全与过敏反应相关,无法真实反映机体的过敏状态。因此,越来越多的研究以动物模型作为辅助评估食品潜在致敏性的工具。根据实验目的选择适合的实验动物和致敏方式,文献报道的花生过敏动物模型有:C3H/HeJ小鼠、BALB/c小鼠、C57/BL6小鼠、BN大鼠、豚鼠及幼猪、狗等[39]。近交系BALB/c小鼠是Th1型细胞应答为主导的动物,对过敏原更易感,受过敏原刺激后可产生高效价的IgE,常用于食物致敏性研究,且相对其他品系动物,其具有经济优势。但由于BALB/c小鼠易产生免疫耐受,其作为食物过敏动物模型的可行性也一直存在争议。C3H/HeJ小鼠在过敏原激发后产生明显的与人类相似的过敏临床症状,它是高IgE应答和Toll样受体4基因突变型品系,也是近几年国内外研究中最常用的品系。BN大鼠与其他种属动物相比,更适合用于研究食物中蛋白质在人体中的致敏性,其大小适中,易连续采血观察血清特异性IgE和IgG动态变化;还可研究致敏大鼠经口服激发后肠道渗透性、呼吸功能和血压等变化。幼猪和狗更多是用于过敏机制研究而非致敏性评价[40]。常用的致敏方式有灌胃、腹腔 注射、皮下注射、静脉注射等。灌胃给予药物是最接近食物摄取的方式,但对于样品量少、需直观检测应激反应的实验则需采用注射方式进行。几种花生致敏动物模型总结见表2。

表2 几种花生致敏动物模型列表Table 2 List of several animal models for peanut sensitization
3 结 语
综上所述,目前为止并没有一种行之有效的花生完全脱敏方法,通过研究各种加工方式降低花生致敏性的机制,开发综合加工方法脱除花生致敏性是今后重点研究方向。要从过敏反应过程入手,结合过敏原构象变化,在免疫血清学检测、体外细胞实验水平和体内动物实验水平,构建多级评价结构模式,深入探讨致敏过程中炎性介质及细胞因子的变化情况与花生致敏蛋白构象间的相关性,从而深层次揭示不同改性加工方法对花生蛋白致敏性的影响规律,为有效降低花生过敏性提供理论依据。
[1]吕相征, 刘秀梅, 杨晓光. 健康人群食物过敏状况的初步调查[J]. 中国食品卫生杂志, 2005, 17(2): 119-121.
[2]BRANUM A M, LUKACS S L. Food allergy among children in the United States[J]. Pediatrics, 2009, 124(6): 1549-1555.
[3]GRUNDY J, MATTHEWS S, BATEMANB, et al. Rising prevalence of allergy to peanut in children: data from 2 sequential cohorts[J]. Journa l of Allergy and Clinical Immunology, 2002, 110(5): 784-789.
[4]HOURIHANE J O B, AIKEN R, BRIGGS R, et al. The impactof government advice to pregnant mothers regarding peanut avoidance on the prevalence of peanut allergy in United Kingdom children at school entry[J]. Journal of Allergy and Clinical Immunology, 200 7, 1 19(5):1197-1202.
[5]CHUNG S Y, BUTTS C L, MALEKI S J, et al. Linking peanut allergenicity to the processes of maturation, curing, and roasting[J]. Journal of Agricultural and Food Chemistry, 2003, 51(15): 4273-4277.
[6]KROGHSBO S, RIGBY N M, JOHNSON P L F, et al. Assessment of the sensitizing potential of processed peanut proteins in Brown Norway rats: roasting does not enhance allergenicity[J]. PLoS ONE,2014, 9(5): e96475. doi: 10.1371/journal.pone.0096475.
[7]BLA NC F, VISSERS Y M, ADEL P K, et al. Boiling peanut Ara h 1 results in the formation of aggregates with reduced allergenicity[J]. Molecular Nutrition & Food Research, 2011, 55(12): 1887-1894.
[8]VISSERS Y M, BLANC F, SKOV P S, et al. Effect of heat ing and glycation on the allergenicity of 2S albumins (Ara h 2/6) from peanut[J]. PLoS ONE, 2011, 6(8): e23998. doi: 10.1371/journal. pone.0023998.
[9]TEUBER S S. Hypothesis: the protein body effectand other aspects of food matrix effects[J]. Annals of the New York Academy of Sciences,2002, 964(1): 111-116.
[10] MALEKI S J, CHUNG S Y, CHAMPAGNE E T, et al. The effects of roasting on the allergenicpro perties of peanut proteins[J]. Journal of Allergy and Clinical Immunology, 2000, 106(4): 763-768.
[11] 王丽, 王强, 刘红芝, 等. 花生加工特性与品质评价研究进展[J]. 中国粮油学报, 2011, 26(10): 122-128.
[12] LEHRER S B, AYUSO R, REESE G. Current understanding of food allergens[J]. Annals of the New York Academy of Sciences, 2002,964(1): 69-85.
[13] 王允圃, 刘玉环, 阮榕生, 等. 食品热加工与非热加工技术对食品安全性的影响[J]. 食品工业科技, 2011, 32(7): 463-467.
[14] HU C, CHEN H, GAO J, et al. High-pressure microfluidisa tioninduced changes in the antigenicity and conformation of allergen Ara h 2 purified from Chinese peanut[J]. Journal of the Science of Food and Agriculture, 2011, 91(7): 1304-1309.
[15] 罗春萍. 热加工, 辐照及超高压微射流对花生过敏原Arah6结构与抗原性的影响[D]. 南昌: 南昌大学, 2011: 42-52.
[16] 许 舒婷, 高美须, 支玉香, 等. 电子束辐照对花生过敏原免疫原性及生化性质影响的研究[J]. 核农学报, 2012, 26(7): 1006-1011.
[17] CHUNG S Y, YANG W, KRISHNAMURTHY K. Effects of pulsed UV-light on peanut allergens in extracts and liquid peanut butter[J]. Journal of Food Science, 2008, 73(5):C400-C404.
[18] YANG W W, MWAKATAGE N R, GOODRICH S R, et al. Mitigation of major peanut allergens by puls ed ultraviolet light[J]. Food and Bioprocess Technology, 2012, 5(7): 2728-2738.
[19] 王烁, 吴海文, 邱志龙, 等. 改性处理对花生蛋白致敏性的影响[J].食品研究与开发, 2011, 32(4): 1-4.
[20] CHUNG S Y, MALEKI S J, CHAMPAGNE E T. Allergenic properties of roasted peanut allergens may be reduced by peroxidase[J]. Journal of Agricultural a nd Food Chemistry, 2004, 52(14): 4541-4545.
[21] CHUNG S Y, KATO Y, CHAMPAGNE E T. Polyphenol oxidase/ caffeic acid may reduce the allergenic properties of peanut allergens[J].Journal of the Science of Food and Agriculture, 2005,85(15): 2631-2637.
[22] MORENO F J. Gastroint estinal digestion of food allergens: effect on their allergenicity[J]. Biomedicine & Pharmacotherapy, 2007, 61(1):50-60.
[23] BURKS A, WILLIAMS L W, THRESHER W, et al. Allergenicity of peanut an d soybean extracts altered by chemical or thermal denaturation in patients with atopic dermatitis and positive food challenges[J]. Journal of Allergy and Clinical Immunology, 1992,90(6): 889-897.
[24] HON G S J, MICHAEL J G, FEHRINGER A, et al. Pepsin-digested peanut contains T-cell epitopes but no IgE epitopes[J]. Journal of Allergy and Clinical Immun ology, 1999, 104(2): 473-477.
[25] KOPPELMAN S J, HEFLE S L, TAYLOR S L, et al. Digestion of peanut allergens Ara h 1, Ara h 2, Ara h 3, and Ara h 6: a comparative in vitro study and partial characterization of digestion-resistant peptides[J]. Molecular Nutrition & FoodResearch, 2010, 54(12):1711-1721.
[26] 何伟逸, 吴序栎, 刘志刚, 等. 消化酶对花生蛋白抗原性影响的研究[J].食品科技, 2013, 38(8): 90-94.
[27] NAKAGAWA T. IgE antibody determination[J]. Japanese Journal of Clinical Med icine, 2001, 59(10): 1950-1954.
[28] 石良, 王锡昌, 刘源, 等. 食物过敏原免疫学检测技术研究进展[J].分析测试学报, 2010, 29(9): 981-986.
[29] 张谦. 血清特异性IgG用于诊断食物过敏的分析[J]. 北京医学,2010(1): 31-34.
[30] DIBBERN J D A, PALMER G W, WILLIAMS P B, et al. RBL cells expressing human FcεRI are a sensitive tool for explo ring functional IgE-allergen interactions: studies with sera from peanut-sensitive patients[J]. Journ al of Immunological Methods, 2003, 274(1): 37-45.
[31] PRICKETT S R, VOSKAMP A L, DACUMOS H A, et al. Ara h 2 p eptides containing dominant CD4+T-cell epitopes: candidates for a peanut allergy therapeutic[J]. Journ al of Allergy and Clinical Immunology, 2011, 127(3): 608-615.
[32] THOTTINGALT B,STEFURA B P, SIMONS F, et al. Human subjects without peanut allergy demonstrate T cell-dependent,TH 2-biased, peanut-specific cytokine and chemokine responses independent of TH1 expression[J]. Journal of Allergy and Clinical Immunology, 2006, 118(4): 905-914.
[33] DELONG J H, SIMPSON K H, WAMBRE E, et al. Ara h 1: reactive T cells in individuals with peanut allergy[J]. Journal of Allergy and Clinical Immunology, 2011, 127(5): 1211-1218.
[34] WANG M, TAKEBA K, SHITAISHI Y, et al. Pean ut-induced intestinal allergy is mediated through a mast cell-IgE-FcεRI: IL-13 pathway[J]. Journal of Allergy and Clinical Immunology, 2010,126(2): 306-316.
[35] FENG B S, CHEN X, HE S H, et al. Disruption of T-cell immunoglobulin and mucin domain molecule (TIM)-1/TIM4 interactionas a therapeutic strategy in a dendritic cell: induced peanut allergy model[J]. Journal of Allergy and Clinical Immunology, 2008,122(1): 55-61.
[36] PALMER G W, DIBBERN J D A, BU RKS A, et al. Comparative potency of Ara h 1 and Ara h 2 in immunochemical and functional assays of allergenicity[J]. Clinical Immunology, 2005, 115(3): 302-312.
[37] SAVAGE J H, COURNEYA J P, STERBA P M, e t al. Kinetics of mast cell, basophil, and oral food challenge responses in omalizumabtreated adults with peanut allergy[J]. Journal of Allergy and Clinical Immunology, 2012, 130(5): 1123-1129.
[38] CHU D K, LLOP G A, WALKER T D, et al. IL-33, but not thymic stromal lym phopoietin or IL-25, is central to mite and peanut allergic sensitization[J]. Journal of Allergy and Clinical Immunology, 2013,131(1): 187-200.
[39] LEHRER S B, MCC LAIN S. Utility of animal models for predicting human allergenicity[J]. Regulatory Toxicology and Pharmacology,2009, 54(Suppl 3): 46-51.
[40] DEARMAN R J, KIMBER I. Animal models of protein allergenicity:potential benef its, pitfalls and challenges[J]. Clinical & Experimental Allergy, 2009, 39(4): 458-468.
[41] MEIJERINK M, WELLS J M, TAVERNE N, et al. Immunomodulatory effects of potential probiotics in a mouse peanut sensitization model[J]. FEMS Immunology & Medical Microbiology,2012,65(3): 488-496.
[42] KULIS M, CHEN X, LEW J, et al. The 2S albumin allergens of Arachis hypogaea, Ara h 2 and Ara h 6, are the major elicitors of anaphylaxisand can effectively desensitize peanut-allergic mice[J]. Clinical & Experimental Allergy, 2012, 42(2): 326-336.
[43] 刘志刚, 杨成彬, 闫浩, 等. Balb/c小鼠花生过敏模型的建立及发病机理[J]. 深圳大学学报: 理 工版, 2012, 29(2): 165-170.
[44] ARIAS K, CHU D K, FLADER K, et al. Distinct immune effector pathways contribute to the full expression of peanut-induced anaphylactic reactions in mice[J]. Journal of Al lergy and Clinical Immunology, 2011, 127(6): 1552-1561.
[45] SCHULZ V J, SMIT J J, HUIJGEN V, et al. Non-dioxin-like AhR ligands in a mouse peanut allergy model[J]. Toxicological Sciences,2012. doi: 10.1093/toxsci/kfs131.
[46] B.GH K L, KROGHSBO S, DAHLL, et al. Digested Ara h 1 has sensitizing capacity in Brown Norway rats[J]. Clinical & Experimental Allergy, 2009, 39(10): 1611-1621.
[47] KIMBER I, DEARMAN R J, PENNINKS A H, et al. Assessment of protein allergenicity on the basis of immune reactivity: animal models[J]. Environmental Health Perspectives, 2003, 111(8): 1125-1130.
Effects of Processing Methods on Peanut Allergenicity and Evaluation Methods for Peanut Allergenicity: A Review
RAO Huan, TIAN Yang, TAO Sha, XUE Wentong*
(College of Food Science and Nutritional Engineering, China Agricultural University, Beijing100083, China)
Peanut protein is an ideal raw material for food processing, but it is also a food allergen causing high mortality and serious harm to human health. This article reviews the effects of processing methods on peanut allergenicity. Meanwhile,methods for evaluating peanut allergenicity both in vitro and in vivo are summarized. This study will hopefully provide guidance for the development of hypoallergenic or non-allergenic peanut products.
processing; peanut allergens; allergenicity evaluation
TS201.1
A
1002-6630(2015)23-0283-05
10.7506/spkx1002-6630-201523052
2015-01-29
国家自然科学基金面上项目(31471614)
饶欢(1988—),女,博士研究生,研究方向为蛋白质工程。E-mail:raohuan881210@163.com
薛文通(1962—),男,教授,博士,研究方向为蛋白质工程。E-mail:xwt@cau.edu.cn
猜你喜欢
金属加工(热加工)(2023年2期)2023-02-27
金属加工(热加工)(2023年1期)2023-02-02
食品工业科技(2022年20期)2022-10-12
锻压装备与制造技术(2021年3期)2021-08-13
家庭科学·新健康(2021年2期)2021-02-22
农业科技与信息(2020年8期)2020-12-18
金属加工(热加工)(2020年12期)2020-02-06
分析化学(2018年8期)2018-11-01
食品与机械(2017年5期)2017-07-05
中外葡萄与葡萄酒(2016年1期)2016-03-28
