
基于气相色谱-质谱技术对用咪康唑处理的白念珠菌的代谢组学研究
2015-08-05 04:50曹颖瑛朱臻宇第二军医大学药学院药物分析学教研室新药研究中心上海200433
药学实践杂志 2015年3期
杨 宇,王 慧,曹颖瑛,朱臻宇(第二军医大学药学院,.药物分析学教研室;.新药研究中心,上海 200433)
随着免疫功能低下患者的增加、器官移植等医疗技术的发展以及免疫抑制剂的使用,真菌感染率居高不下,其中白念珠菌成为真菌感染的主要病原菌[1]。
咪康唑因生物利用度好和不良反应少成为防治真菌感染的一线药物。研究表明,咪康唑可通过影响过氧化物酶和线粒体而发挥抗真菌活性[2]。同时,咪康唑可增加活性氧使细胞死亡[3]。然而,目前咪康唑的作用机制尚不明确,因此研究其作用机制显得尤为重要。
在生命系统中,代谢组能将基因组、转录组和蛋白组的微小变化放大。因此,代谢组学能更直接地反应生物体的功能变化。
笔者基于气相色谱-质谱(GC-MS)代谢组学技术,研究咪康唑给药前后白念珠菌的代谢特征谱,结合药物作用机制,探讨代谢物产生差异的原因,为阐明药物作用机制提供参考。
1 仪器与试药
1.1 仪器 Thermo Trace Ultra/DSQⅡGC-MS(美国赛默飞公司);微型漩涡混合仪(美国赛默飞公司);-80℃低温冰箱(美国赛默飞公司);冻干机(美国Virtis公司);MJx型智能霉菌培养箱(宁波江南仪器厂)。
1.2 试剂 甲氧基胺盐酸盐、N-甲基-N-(三甲基硅烷)三氟乙酰胺(MSTFA)、吡啶、三甲基氯硅烷(TMCS)、咪康唑均购自Sigma公司;正庚烷(上海晶纯实业公司)。
1.3 菌株和培养基 白念珠菌SC5314;YPD培养液:酵母浸膏10g,蛋白胨20g,葡萄糖20g,溶解于1 000ml三蒸水,高压灭菌(121 ℃,15min),4 ℃保存。
2 方法
2.1 真菌培养
2.1.1 咪康唑半数抑菌浓度(IC50)的确定 挑取白念珠菌单克隆,接种至1ml YPD培养液,于30℃,200r/min培养至指数生长后期,于紫外分光光度计600nm[4]下测菌液光密度(D)值,用 YPD培养液调整菌液浓度至D值为0.1,取20ml加入各锥形瓶中培养,至D值为0.2时,给药组中加入咪康唑,使其终浓度分别为0.5、1、2、4、8、16、32、64μg/ml,继续培养(对照组中加入相同体积的二甲基亚砜,其浓度应小于0.05%,以确保不影响白念珠菌的生长),分别在3、4、5、6h测定各组的D值。与对照组的D值相比,选择继续培养4h,其中咪康唑的IC50为16μg/ml。
2.1.2 培养条件 按照IC50的测定方法和给药浓度,培养给药组和对照组各6份。
2.2 样品前处理
2.2.1 样品淬灭 向样品中加入等体积的60%甲醇(预冷至-20 ℃),迅速摇匀,静置5min(-40℃),6 000r/min离心。
2.2.2 样品提取 样品用超纯水迅速清洗后,重悬于1ml沸水(含10μl 200mmol/L的α-氨基丁酸内标)[4],静置15min,放入 -80 ℃ 冰箱冰 冻15min,取出,60℃水浴溶解15min,再重复冻融操作2次,13 200r/min离心,取上清液,放置在-20℃冰箱预冻。后置于冻干机中冻干,获得冻干粉。
2.2.3 衍生化 向冻干粉中加入75μl甲氧基胺盐酸盐/吡啶溶液,40℃温孵90min,再加入75μl MSTFA 溶液 (1%TMCS),40 ℃ 温孵 50min,13 200r/min离心,取上清液,装入进样小瓶。
2.2.4 菌体干重的测定 将样品沉淀在室温下风干至恒重,称样品干重,至少称量3次以确保样品完全干燥。
2.3 GC-MS条件 毛细管柱:HP-5MS石英毛细管柱(30m×0.25mm,0.25μm);进样口温度:260℃;升温程序:起始温度为70℃,保持3min,4℃/min升至220℃,再8℃/min升至310℃,保持10min,图谱从第7.6分钟开始采集;载气:高纯氮气;流速:1.0ml/min。进样量为1μl。
电子轰击源(EI);离子源温度:200℃;接口温度:280℃;电子能量:70eV;调谐方式:标准调谐;质谱扫描方式:扫描范围15~800amu,扫描速度5s/dec。
2.4 数据处理 将原始数据(.RAW)经Xculibur数据处理系统转化成CDF格式,利用XCMS软件对数据峰校正和峰积分,并采用 MATLAB 7.0(The MathWorks,Inc.,USA)过滤离子峰,留下相同保留时间下峰度最大的离子峰。为校正质谱响应,每个样品的各个峰面积先除以各自的细胞干重,再除以内标衍生化后峰度最大的离子峰。利用SIMCA-P V 11.0(Umetrics,Sweden)软件将数据中心化和帕累托变换,标准化后进行主成分分析(PCA)和偏最小二乘法(PLS-DA)分析,根据variable influence in the projection(VIP)值来预测代谢物对模型的贡献程度。VIP>1的数据对模型有明显贡献。接着对数据进行独立样本t检验,保留P<0.01的数据。将数据与NIST数据库匹配,并用标准品定位,得到潜在的生物标志物。
3 结果
3.1 GC-MS色谱图 给药组和对照组白念珠菌的胞内代谢物,经提取,衍生化后进样分析,其GC-MS色谱图,如图1所示。
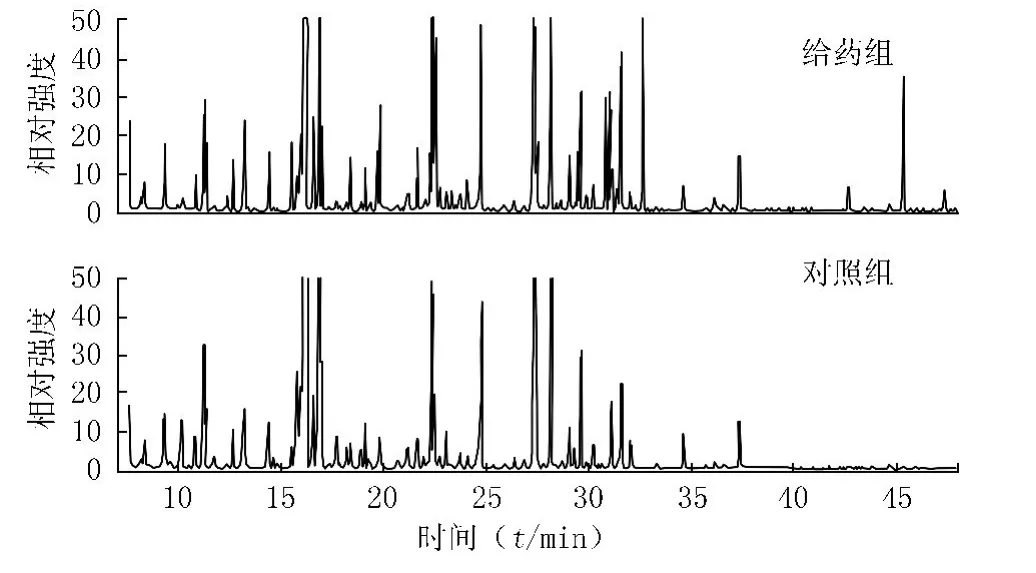
图1 给药组和对照组的GC-MS色谱图
3.2 PCA与PLS-DA结果 数据校正后,对给药组和对照组进行PCA和PLS-DA分析,如图2A和2B所示,结果表明两组明显分开。其载荷图和S-plot图,如图2C和2D所示。载荷图用于潜在生物标志物的筛选,根据载荷图所示,离原点越远的离子对分类贡献越大。得到23个潜在的生物标志物,给药组和对照组代谢物含量变化见图3。
4 讨论
根据给药组和对照组的差异代谢物实验结果,分别讨论如下:
4.1 葡萄糖 给药组与对照组相比,葡萄糖含量显著升高。研究表明咪康唑通过影响线粒体ATP酶和钾离子泵而发挥抗真菌活性[5],其中钾离子泵是主动运输,消耗能量,而葡萄糖可产生能量。因此,葡萄糖含量的升高可能与咪康唑作用钾离子通道有关。
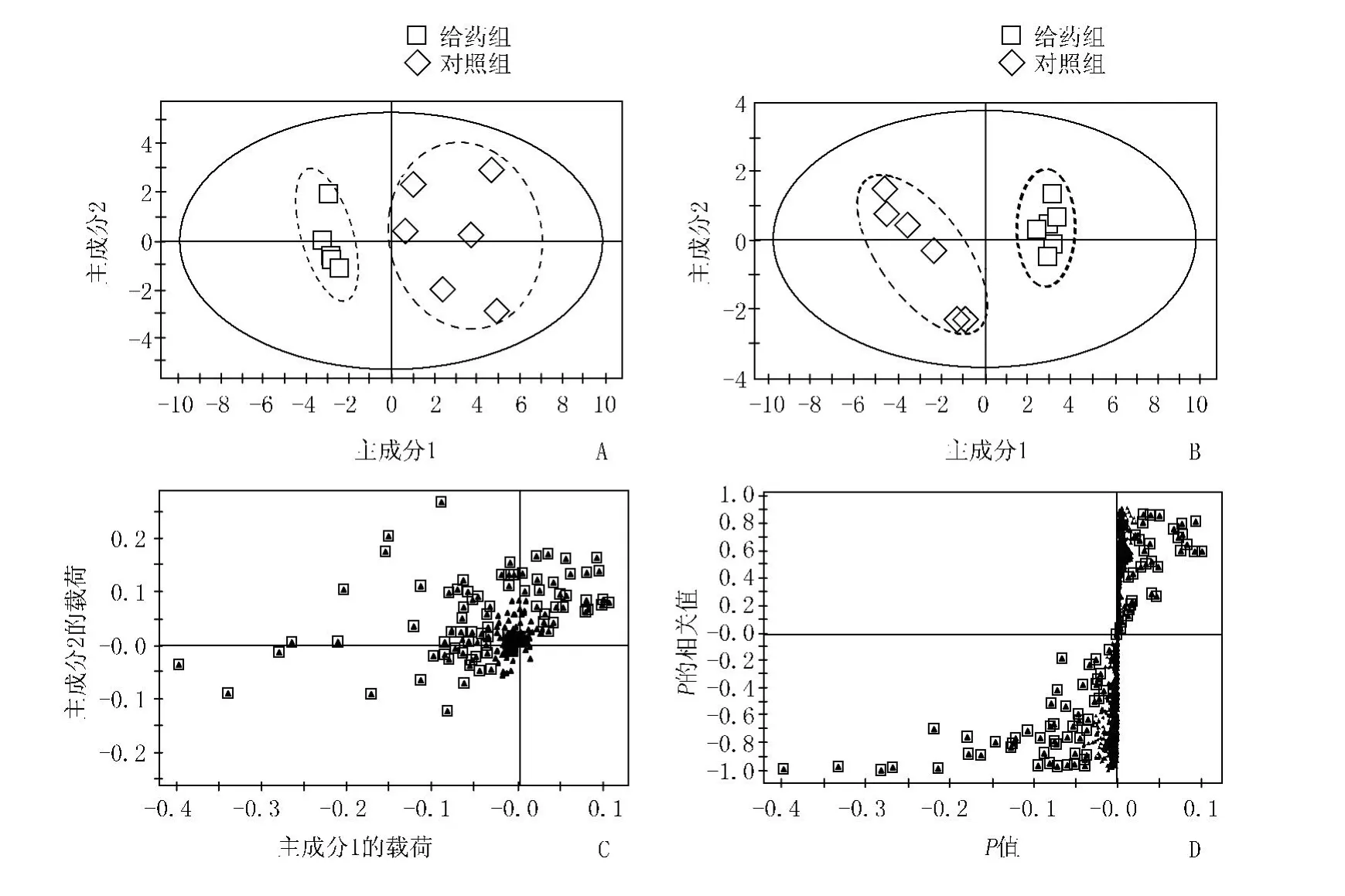
图2 给药组和对照组的主成分分析图(A)、偏最小二乘法得分图(B)、载荷图(C)和S-plot图(D)
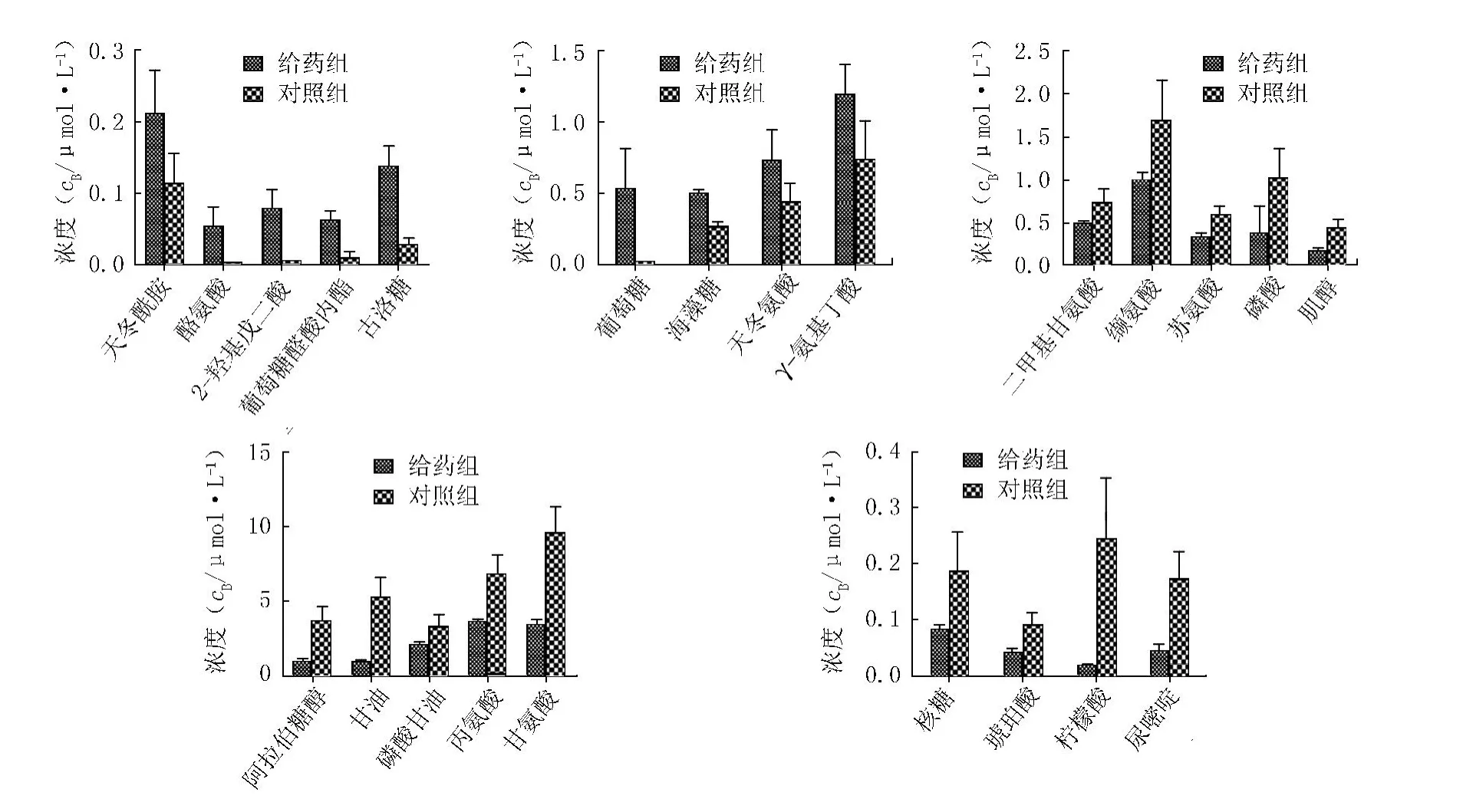
图3 给药组和对照组的差异代谢物柱状图
4.2 海藻糖 白念珠菌受到外界刺激时会采取保护性防御机制,其中氧化应激刺激细胞内非还原性二糖海藻糖明显增加[6]。在本实验中,咪康唑可能是一种氧化应激,给药后使细胞内海藻糖大量积累,从而起到抗氧化的作用。
4.3 三羧酸循环相关代谢物 三羧酸循环不仅是需氧生物体重要的代谢途径,还是糖类、脂质和氨基酸代谢联系的枢纽。酸刺激下,胞外谷氨酸通过反向转运体(GadT)转移至胞内,在谷氨酸脱羧酶的作用下生成γ-氨基丁酸(GABA),该反应消耗氢离子,使胞内pH值升高。进而GABA通过GABA支路在GABA/α-酮戊二酸氨基转移酶的作用下脱氨基形成琥珀酸半醛(SSA),SSA在琥珀酸半醛脱羧酶的作用下生成琥珀酸,进而影响三羧酸循环[7],同时GABA支路可降低细胞内活性氧[8]。这与前期的研究结果相符[9]。由此可知,三羧酸循环中琥珀酸和柠檬酸含量的降低可能与咪康唑影响GABA支路有关。
4.4 2-羟基戊二酸(2-HG) 研究表明,2-HG脱氢酶活性降低导致2-HG大量积累,而2-HG脱氢酶可降低DNA和组蛋白的甲基化,从而抑制正常细胞向癌细胞转化[10]。因此,咪康唑给药组2-HG的大量积累可能与2-HG脱氢酶活性降低有关,从而促进正常细胞向癌细胞转化。
4.5 氨基酸类 天冬酰胺、天冬氨酸、酪氨酸在给药组中含量升高。其中,酪氨酸磷酸化参与细胞信息传递,因此酪氨酸的增加可能与细胞信息传递有关。天冬氨酸和天冬酰胺可合成多种氨基酸。有研究表明,苏氨酸生物合成中间体β-天冬氨酸半醛(ASA)的积累对细胞有一定的损伤作用,同时可加速Gcn4的降解,Gcn4可调控多种氨基酸和维生素的生物合成[11]。因此丙氨酸、二甲基甘氨酸、缬氨酸、苏氨酸和甘氨酸含量的降低与咪康唑促使ASA积累有关。
4.6 磷酸、磷酸甘油、甘油类 磷脂是细胞膜的重要组成部分。Pasrija等[12]报道药物外转运蛋白倾向定位于细胞膜。因此,咪康唑可能通过影响细胞膜而导致药物外转运蛋白的活性降低,使细胞内的药物积累而发挥治疗作用。
本实验基于GC-MS的代谢组学方法对咪康唑给药前后白念珠菌的代谢物进行研究,经比较分析,得到23个潜在的生物标志物,它们主要参与了氨基酸代谢、三羧酸循环、氧化应激、糖酵解和磷脂代谢等相关通路,对咪康唑抗真菌作用机制的阐明具有重要意义。
[1]Kriengkauykiat J,Ito JI,Dadwal SS.Epidemiology and treatment approaches in management of invasive fungal infections[J].Clin Epidemiol,2011,3:175-191.
[2]Vandenbosch D,De Canck E,Dhondt I,et al.Genomewide screening for genes involved in biofilm formation and micon-azole susceptibility in Saccharomyces cerevisiae[J].FEMS Yeast Res,2013,13(8):720-730.
[3]Zhu CX,Yan L,Wang XJ,et al.Transposition of the Zorro2retrotransposon is activated by miconazole in Candida albicans[J].Biol Pharm Bull,2014,37(1):37-43.
[4]吴海棠,李 祥,王添琦,等.黄芩素作用白假丝酵母菌的GCMS代谢组学研究[J].第二军医大学学报,2013,34(2):184-189.
[5]Calahorra M,Lozano C,Sanchez NS,et al.Ketoconazole and miconazole alter potassium homeostasis in Saccharomyces cerevisiae[J].Biochim Biophys Acta,2011,1808(1):433-445.
[6]Sanchez-Fresneda R,Guirao-Abad JP,Arguelles A,et al.Specific stress-induced storage of trehalose,glycerol and D-arabitol in response to oxidative and osmotic stress in Candida albicans[J].Biochem Biophys Res Commun,2013,430(4):1334-1339.
[7]Feehily C,O'Byrne CP,Karatzas KA.Functionalγ-Aminobutyrate Shunt in Listeria monocytogenes:role in acid tolerance and succinate biosynthesis[J].Appl Environ Microbiol,2013,79(1):74-80.
[8]Cao J,Barbosa JM,Singh NK,et al.GABA shunt mediates thermotolerance in Saccharomyces cerevisiae by reducing reactive oxygen production[J].Yeast,2013,30(4):129-144.
[9]Kobayashi D,Kondo K,Uehara N,et al.Endogenous reactive oxygen species is an important mediator of miconazole antifungal effect[J].Antimicrob Agents Chemother,2002,46(10):3113-3117.
[10]Shim EH,Livi CB,Rakheja D,et al.L-2-hydroxyglutarate:an epigenetic modifier and putative oncometabolite in renal cancer[J].Cancer Discov,2014,4(11):1290-1298.
[11]Rawal Y,Qiu H,Hinnebusch AG.Accumulation of a threonine biosynthetic intermediate attenuates general amino acid control by accelerating degradation of Gcn4via Pho85and Cdk8[J].PLoS Genet,2014,10(7):e1004534.
[12]Pasrija R,Panwar SL,Prasad R.Multidrug transporters CaCdr1p and CaMdr1p of Candida albicans display different lipid specificities:both ergosterol and sphingolipids are essential for targeting of CaCdr1p to membrane rafts[J].Antimicrob Agents Chemother,2008,52(2):694-704.
猜你喜欢
医学理论与实践(2022年21期)2022-11-10
现代临床医学(2022年4期)2022-09-29
中国合理用药探索(2022年8期)2022-09-09
中外医学研究(2022年22期)2022-09-09
中山大学学报(自然科学版)(中英文)(2022年4期)2022-08-05
首都食品与医药(2022年11期)2022-06-06
国际医药卫生导报(2022年7期)2022-04-12
甘肃医药(2021年5期)2021-05-23
医学食疗与健康(2019年6期)2019-09-10
分忧(2015年3期)2015-06-08
