
福氏志贺菌2型外膜蛋白A的原核表达及多克隆抗体的制备
2015-06-18 11:27:52刘祥
动物医学进展 2015年5期
刘 祥
(陕西理工学院生物科学与工程学院,陕西汉中 723001)
福氏志贺菌(Shigellaflexneri)为革兰阴性致病菌,属于高度传染性和危害严重的急性肠道传染病,临床上可引起细菌性痢疾,病死率高[1-2];世界各地都有流行,尤其是发展中国家,患病群体主要为儿童[3]。随着抗生素的滥用,福氏志贺菌产生不同程度的耐药性[4],有待开发一种新型的安全、无耐药性的亚单位疫苗。外膜蛋白 A(outer membrane protein A,OmpA)为革兰阴性菌主要外膜蛋白之一[5],可促进福氏志贺菌与宿主菌的黏附与侵袭[6],是福氏志贺菌主要毒力蛋白。此外,研究发现将OmpA免疫小鼠可激活机体的细胞免疫和体液免疫水平[7],并产生很小的炎症反应,对福氏志贺菌的感染有很强的免疫保护作用[7-8]。有望作为一种新型的亚单位蛋白疫苗[9]。
本研究利用分子克隆方法获得福氏志贺菌OmpA蛋白表达菌株;纯化OmpA蛋白,免疫小鼠,制备小鼠OmpA抗血清;为OmpA蛋白介导福氏志贺菌侵染宿主的靶向蛋白研究,以及OmpA免疫保护功能及疫苗的开发研究奠定基础。
1 材料与方法
1.1 材料
1.1.1 菌株、质粒与实验动物Shigellaflexneri2a,E.coliDH5α,E.coliBL21菌株,pET-32a质粒本实验室保存;昆明鼠购于西安交通大学实验动物中心。
1.1.2 主要试剂 限制性内切酶,Taq酶,T4-DNA连接酶,核酸标准,蛋白标准均购自宝生物工程(大连)技术服务公司;质粒提取试剂盒、胶回收试剂盒购自上海生物工程公司;引物由西安沃尔森生物技术有限公司合成;基因测序为北京奥科鼎盛生物科技有限公司;IPTG、二抗为Sigma公司产品。
1.2 方法
1.2.1 OmpA重组质粒的构建 依据NCBI公布的福氏志贺菌2a菌株OmpA基因序列。设计引物:正向引物:5′-ACAGGATCCATGAAAAAGACAGCTAT-3′;逆向引物:5′-GCTAAGCTTTTAAGCCTGCGGCTGAGT-3′,下划线为BamHⅠ和HindⅢ限制性内切酶位点。PCR采用50μL反应体系:缓冲液5μL,基因组模板3μL,10mmol/L dNTP 2μL,25μmol/L引物各1μL,Taq酶0.5 μL,补水至50μL。PCR反应条件为:94℃3min;94℃30s,55℃ 45s,72℃ 90s,30个循环;最后72℃延伸10min,16℃终止反应。采用8g/L琼脂糖凝胶电泳检测PCR片段大小。将PCR产物与pET-32a质粒载体双酶切后,利用T4-连接酶连接,转化E.coliDH5α,提取重组质粒双酶切检测与测序检测确认后,转入表达菌株E.coliBL21,构建重组蛋白的表达菌株。
1.2.2 OmpA重组蛋白的表达检测与纯化 将重组表达菌株于LB液体培养基中培养至OD600约0.5时,加入终浓度0.5mmol/L IPTG,37℃诱导培养6h,收集菌体,SDS-PAGE蛋白电泳检测OmpA表达情况;并利用SDS-PAGE电泳切胶的方法,对OmpA蛋白进行纯化。
1.2.3 OmpA重组蛋白的复性 采用尿素浓度梯度的方法复性蛋白,主要步骤为:将纯化的OmpA蛋白溶解于8mol/L尿素溶液并置透析袋中,然后放入含6mol/L 尿素的50mmol/L Tris-HCl(pH 7.4)中,4℃放置4h;接着依次为4、2、1、0.5mol/L尿素,最后置于50mmol/L Tris-HCl(pH7.4)中,4℃放置4h;接着离心取上清溶液,吸取2μL用于蛋白电泳检测。
1.2.4 OmpA重组蛋白小鼠多克隆抗体制备 选取约5周龄的昆明鼠6只,50μg/只免疫OmpA蛋白,首次免疫采用弗氏完全佐剂。第1次免疫14d后进行第2次免疫,7d后,进行第3次加强免疫,再7d后小鼠眼部取血并置于4℃冰箱中析出,最后离心取上清,置-80℃冰箱中保存待用。
1.2.5 OmpA重组蛋白抗体效价检测 通过酶联免疫吸附试验(ELISA)检测抗血清效价,主要步骤为:将OmpA蛋白溶解至0.05μg/μL,在相应的96孔板中加入100μL溶液,37℃ 作用1h,然后加入300μL封闭液,37℃ 孵育2h,接着每孔加100μL不同滴度的小鼠一抗血清,37℃孵育30min;充分洗涤后加入100μL二抗,37℃作用30min;充分洗涤加入底物A与底物B的混合液100μL,37℃避光显色10min,迅速加入终止液,最后置于酶标仪450 nm处读OD值。
1.2.6 OmpA重组蛋白抗体特异性检测 利用Western blot方法检测小鼠 OmpA抗血清特异性[10]。为避免OmpA重组蛋白中的融合蛋白所产生抗体对试验结果的影响,将福氏志贺菌培养,收集菌体裂解后进行SDS-PAGE蛋白电泳,然后转NC膜,与不同稀释度的小鼠抗血清(对照为阴性抗血清)作用1h,然后加入二抗孵育1h,最后DAB显色,确定OmpA抗血清的特异性。
1.2.7 OmpA蛋白序列系统发生分析 根据NCBI数据库公布的OmpA蛋白氨基酸序列,利用DNAMan软件进行同源性分析;采用MEGA软件Neighbour-Joining方法构建系统发生树。
2 结果
2.1 重组载体的构建
通过PCR扩增获得约1 000bp左右片段,与目的基因大小一致(图1-A1)。将PCR获得的目的基因片段与pET-32a质粒进行双酶切处理后,T4-DNA连接酶连接,转化克隆菌株E.coliDH5α。提取重组质粒,BamHⅠ与HindⅢ双酶切处理,获得大小与预测一致的1 047bp片段(图1A2)。此外,序列测序结果证实目的基因与公布的序列相同。
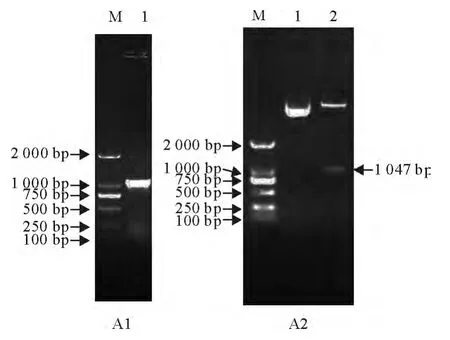
图1 福氏志贺菌OmpA基因重组质粒构建Fig.1 Recombinant plasmid construction of S.flexneri OmpA gene
2.2 OmpA的表达检测与蛋白的纯化、复性
将检测成功构建的重组质粒,转化表达菌株E.coliBl21,获得OmpA表达菌株,经IPTG诱导后,蛋白电泳获得约57ku条带,其中包含OmpA蛋白约37ku,以及pET-32a质粒的融合蛋白标签20.4 ku。蛋白电泳结果显示OmpA重组蛋白大小与理论预期数值一致。此外,利用SDS-PAGE电泳切胶纯化,以及尿素浓度梯度复性的方法获得OmpA蛋白(图2)。
2.3 OmpA蛋白抗血清效价检测
将OmpA蛋白免疫小鼠,获得的抗血清经酶联法检测,发现其抗体效价达到1∶1 600(图3)。
2.4 OmpA蛋白抗血清特异性检测
采用Western blot方法发现不同稀释度OmpA抗血清出现明显条带;而对照阴性抗血清无对应条带(图4)。表明OmpA抗血清可与OmpA蛋白特异性结合。
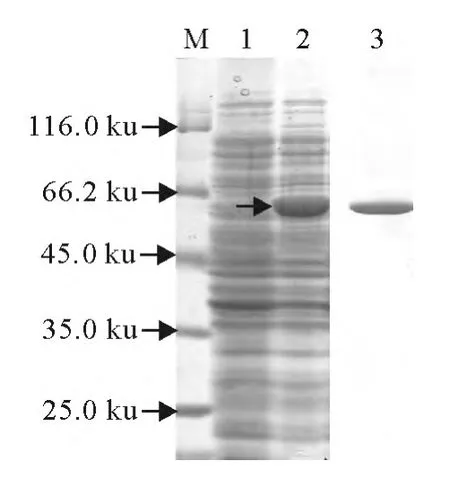
图2 OmpA蛋白表达、纯化与复性Fig.2 The expression,purification and renaturation of OmpA protein

图3 OmpA多克隆抗体效价的检测Fig.3 Detection of the OmpA polyclonal antibody titer
2.5 OmpA蛋白的氨基酸序列分析
利用DNA Man软件,对选取的10种致病性菌株OmpA蛋白的氨基酸序列进行同源性分析。结果表明:不同细菌OmpA蛋白的C-端存在较高的同源性;不同种的志贺菌,如福氏志贺菌,痢疾志贺菌(Shigelladysenteriae),宋内志贺菌(Shigellasonnei),鲍氏志贺菌(Shigellaboydii)存在更高的同源性(图5)。
采用MEGA5.02软件构建的OmpA系统进化树发现四种志贺菌的亲缘关系较近,福氏志贺菌与痢疾志贺菌亲缘关系更近;动物致病菌副猪嗜血杆菌(Haemophilusparasuis)和多杀性巴氏杆菌(Pasteurellamultocida)亲缘关系近。

图4 Western blot检测OmpA多克隆抗体特异性Fig.4 Identification of the specificity of OmpA polyclonal antibodies by Western blot
3 讨论
志贺菌是细菌性痢疾最常见的病原菌,分为福氏志贺菌,痢疾志贺菌,宋内志贺菌,鲍氏志贺菌[11]。其中福氏志贺菌属于高度传染性的急性肠道传染病,其通过侵袭并穿透黏膜,破坏结肠上皮层,引起细菌性痢疾[1-2]。OmpA蛋白在介导福氏志贺菌黏附侵染宿主细胞,以及诱导宿主的机体免疫上起到重要作用。研究证实OmpA的突变改变福氏志贺菌膜的结构,从而减弱其毒力,影响细菌对宿主的黏附[6]。此外,OmpA免疫小鼠可显著增加小鼠特异性的抗体水平,提高机体对福氏志贺菌的抵抗能力[7]。而获得抗体是OmpA功能与作用研究的重要手段。多克隆抗体以其成本低、周期短及效果好得到广泛运用[12]。本试验利用分子与免疫学方法获得OmpA蛋白原核表达载体,纯化OmpA蛋白,并成功制备多克隆抗体,为揭示OmpA蛋白功能及相关疾病的发生机制研究奠定基础。
OmpA蛋白具有重要的生物学功能,在遗传进化上具有相对保守性。本试验对OmpA序列系统发生分析研究发现不同种的致病菌存在同源性,特别C-端同源性较高,可能与其执行重要的生理学功能有关;并且不同种志贺菌具有更高的同源性。因而,其免疫宿主产生的抗血清可能对不同种类致病菌具有交叉免疫保护作用,为OmpA蛋白疫苗的研制提供依据。

图5 OmpA蛋白氨基酸序列比对Fig.5 Multiple alignment of amino acid sequences of OmpA protein
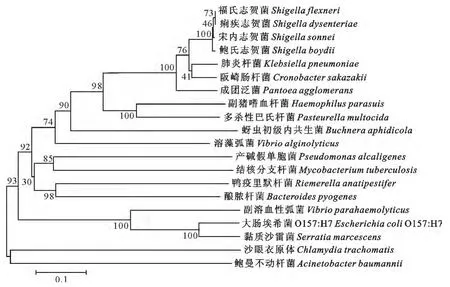
图6 MEGA软件构建的OmpA氨基酸序列系统进化树Fig.6 Phylogenetic tree based on OmpA amino acid sequences by MEGA
[1]George D T,Behm C A,Hall D H,et al.Shigellaflexneriinfection inCaenorhabditiselegans:cytopathological examination and identification of host responses[J].PLoS One,2014,9(9):e106085.
[2]朱 阵,王 婧,张继瑜,等.福氏志贺菌2型多重PCR检测方法的建立 [J].中国畜牧兽医,2014,41(8):34-38.
[3]Dragoi A M,Agaisse H.The serine/threonine kinase STK11 promotesShigellaflexneridissemination through establish-ment of cell-cell contacts competent for tyrosine kinase signaling[J].Infect Immun,2014,82(11):4447-57.
[4]王 俊,祁 伟,王淑香,等.天津地区福氏志贺菌整合子携带及耐药性研究 [J].中国病原生物学杂志,2014,9(2):146-149.
[5]陈春琳,俱 雄,万 健,等.革兰阴性菌外膜蛋白A研究进展[J].生物技术,2014,24(2):98-103.
[6]Ambrosi C,Pompili M,Scribano D,et al.Outer membrane protein A (OmpA):a new player inShigellaflexneriprotru-sion formation and inter-cellular spreading [J].PLoS One,2012,7(11):e49625.
[7]Pore D,Mahata N,Pal A,et al.Outer membrane protein A(OmpA)ofShigellaflexneri2ainduces protective immune response in a mouse model[J].PLoS One,2011,6(7):e22663.
[8]Bhowmick R,Pore D,Chakrabarti M K.Outer membrane protein A (OmpA)ofShigellaflexneri2ainduces TLR2-mediated activation of B cells:involvement of protein tyrosine kinase,ERK and NF-κB [J].PLoS One,2014,9(10):e109107.
[9]Pore D,Chowdhury P,Mahata N,et al.Purification and characterization of an immunogenic outer membrane protein ofShigellaflexneri2a[J].Vaccine,2009,27:5855-5864.
[10]刘 祥.溶藻弧菌附着定植因子ACFA原核载体构建、表达条件优化及多克隆抗体制备 [J].华北农学报,2015,30(1):35-41.
[11]Lee J H,Park H,Park Y H.Molecular mechanisms of host cytoskeletal rearrangements byShigellainvasins[J].Int J Mol Sci,2014,15(10):18253-18266.
[12]刘 祥,陈春琳,牟 欢,等.重组人骨硬化蛋白的表达、纯化及多克隆抗体制备[J].生物技术,2014,24(6):68-72.
猜你喜欢
天津农学院学报(2024年1期)2024-04-07 11:20:02
食用菌(2023年6期)2023-11-28 06:03:32
环球时报(2022-09-20)2022-09-20 15:18:57
今日农业(2021年11期)2021-11-27 10:47:17
今日农业(2020年24期)2020-12-15 16:16:00
食品科学(2018年10期)2018-05-23 01:27:28
兽医导刊(2016年12期)2016-05-17 03:51:50
西南医科大学学报(2015年1期)2015-08-22 13:01:46
中国当代医药(2015年9期)2015-03-01 02:01:59
现代检验医学杂志(2015年4期)2015-02-06 02:02:06
