
Rhizopusoryzae LS-1利用丢糟水解液发酵生产乳酸
2015-06-05 09:51田艳花任海伟刘娜丽
食品工业科技 2015年1期
田艳花,任海伟,刘娜丽
(1.山西药科职业学院食品工程系,山西太原 030031;2. 兰州理工大学生命科学与工程学院,甘肃兰州 730050)
RhizopusoryzaeLS-1利用丢糟水解液发酵生产乳酸
田艳花1,任海伟2,*,刘娜丽1
(1.山西药科职业学院食品工程系,山西太原 030031;2. 兰州理工大学生命科学与工程学院,甘肃兰州 730050)
研究了一株诱变米根霉RhizopusoryzaeLS-1利用丢糟水解液发酵生产乳酸的可行性。首先考察R.oryzaeLS-1利用葡萄糖和木糖的糖代谢差异特性,并通过正交实验优化丢糟水解液发酵生产乳酸的工艺参数。结果表明,R.oryzaeLS-1能代谢利用葡萄糖和木糖,且二者存在协同互补作用,有利于乳酸生成和糖酸转化,可用于木质纤维原料的乳酸生产。丢糟水解液发酵生产乳酸的实验表明,氯化铵是R.oryzaeLS-1适宜的氮源。在接种量为3.0%、pH为6.5、发酵时间为96h、CaCO3添加量为80g/L的条件下,乳酸生成浓度为13.27g/L,糖利用率为79.61%。说明诱变菌株R.oryzaeLS-1具备发酵丢糟水解液制备乳酸的潜力。
丢糟水解液,米根霉,乳酸发酵
乳酸是一种重要的多用途有机酸,学名2-羟基丙酸,被广泛用于食品、医药、饲料、化工等领域。尤其引人关注的是,乳酸聚合而成的聚乳酸(PLA)作为无毒、无刺激、高强度的生物相容性高分子材料能用做制造生物可降解塑料、绿色包装材料和药用修复材料等,应用前景广阔[1]。
乳酸生产有化学合成法、酶转化法以及微生物发酵法。其中,微生物发酵法因其原料来源广泛、生产成本低、产品光学纯度高、安全性好等优点已成为主要的乳酸生产方法。目前报道的乳酸发酵微生物有丝状真菌(Rhizopusoryzae、R.stolonifer、R.ritici,R.elegans)、细菌(lactic acid bacteria,Bacillusspecies)和基因工程菌(Pichiastipitis,Corynebacteriumglutamicum,Candidautilis,Escherichiacoli)等[2]。其中,乳酸菌(Lactobacillus,Lactococcus,Streptococcus,Leuconostoc,Enterococcus)对营养环境和生长培养基的要求严格,只能在氨基酸、维生素和其他生长因子具备的培养基中生长,且产物多为乳酸的外消旋混合物,使得生产和分离纯化成本提高。与之相比,米根霉(Rhizopusoryzae)发酵生产的乳酸具有光学纯度高、营养需求简单、好氧发酵及产物容易提纯等优点,是工业生产高光学纯度乳酸的理想菌株[3]。
木质纤维素主要由纤维素、半纤维素和木质素组成,可降解转化为富含葡萄糖和木糖的混合糖基质用于乳酸发酵生产[4],是乳酸工业化生产的重要原料,如农作物秸秆(麦秸、玉米秸等)[5-6]、玉米芯[7]、木材[8]、废弃报纸[9]、蔗渣[10]等。中国是白酒生产和消费大国。白酒酿造副产物——丢糟不仅产量巨大,资源丰富,而且纤维素和半纤维素含量高达60%以上,是一种亟待开发的木质纤维素资源。若能将其降解糖化后用于发酵生产乳酸,不仅能拓宽乳酸的生产原料来源,降低原料成本,而且对减少工业废弃物污染具有积极意义。
本文以1株诱变的米根霉(Rhizopusoryzae)LS-1为发酵菌种,首先研究了R.oryzaeLS-1的葡萄糖和木糖代谢特性,并以丢糟水解液为原料,选取乳酸浓度和总糖利用率为评价指标,考察了氮源种类、发酵时间、接种量、pH和CaCO3添加量等参数对乳酸发酵工艺的影响,探索丢糟用于乳酸发酵的可行性,为丢糟利用和白酒酿造企业循环发展提供理论依据。
1 材料与方法
1.1 材料与仪器
丢糟,甘肃金徽酒股份有限公司提供。鲜酒糟自然风干后粉碎过50 目筛备用。基本成分为(以干基计):蛋白质质量分数为16.42%±0.03%,淀粉质量分数为14.81%±0.09%,粗脂肪质量分数为3.56%±0.02%,纤维素质量分数为53.17%±1.26%,半纤维素质量分数为12.86%±0.06%。
米根霉(Rhizopusoryzae)LS-1(CICC41411的诱变菌种) 兰州理工大学生命科学与工程学院保藏。
HH-4数显恒温水浴锅 国华电器有限公司;SHD-III 型循环水式多用真空泵 保定高新区阳光科教仪器厂;TDL-5-A离心机 上海安亭科学仪器厂;GZX-9240MBE 数显鼓风干燥箱 上海博讯实业有限公司医疗设备厂;Cary 50 紫外可见分光光度计 上海精密科学仪器有限公司;SHZ-82气浴恒温振荡培养箱 江苏省金坛市荣华仪器制造有限公司。
1.2 培养基
菌种保藏斜面培养基:马铃薯葡萄糖琼脂(PDA)培养基;种子培养基(g/L):葡萄糖 50;(NH4)2SO44;KH2PO40.6,MgSO4·7H2O 0.25,ZnSO4·7H2O 0.44,CaCO310(添加1/10体积的10% CaCO3悬浮液单独灭菌,用于调节pH);生长培养基(g/L):葡萄糖120,(NH4)2SO41,KH2PO40.5,MgSO4·7H2O 0.5,ZnSO4.7H2O 0.04,CaCO325。
1.3 实验方法
1.3.1 分析方法 乳酸测定采用对羟基联苯比色法;木糖测定采用间苯三酚法;总糖(以还原糖计)测定采用DNS法。
1.3.2 液体种子培养方法 LS-1米根霉在PDA斜面培养基上32℃培养5~7d,制备孢子悬浮液。250mL三角瓶中装入50mL种子培养基,接种5mL孢子悬液,在32℃、220r/min条件下振荡培养48h,得到液体种子,计算孢子浓度为5×106个/mL。
1.3.3 糖代谢特性研究 设置不同的C/N比值,考察碳源种类分别为葡萄糖、木糖或混合糖(m葡萄糖∶m木糖=1∶1)条件下米根霉的糖代谢特性。糖浓度分别为90、100、110、120、130、140g/L,培养过程中选择氮源为氯化铵,氯化铵添加量以N浓度计为3.5g/L。培养条件为:250mL三角瓶中装入50mL未加碳源的生长培养基,接种量为5%,32℃摇床培养72h,转速220r/min。发酵结束后计算糖酸转化率和糖利用率。计算公式如下:
糖利用率(%)=初始糖质量(g)-终了糖质量(g)/初始糖质量(g)×100
(1)

(2)
1.3.4 丢糟水解液发酵制备乳酸的工艺参数优化
1.3.4.1 单因素实验 参考文献[11]方法制备丢糟水解液,经测定葡萄糖浓度为15.28g/L,木糖浓度为4.73g/L,总糖浓度为22.19g/L。
采用250mL三角瓶装液50mL进行发酵。准确量取50mL丢糟水解液,依次添加KH2PO4、MgSO4·7H2O和ZnSO4·7H2O,使其浓度分别为0.5g/L、0.5g/L和0.04g/L。灭菌后接入种子培养液进行乳酸发酵,发酵温度32℃,摇床转速为220r/min。发酵结束后,取发酵样液5000r/min、4℃离心10min,取适量上清液测定乳酸浓度和总糖浓度,并按照公式(1)计算糖利用率。
a.氮源种类对乳酸发酵的影响
在总糖浓度为100g/L、碳源为葡萄糖和木糖(m葡萄糖∶m木糖=1∶1)的条件,考察硫酸铵(AS)、氯化氨(AC)、尿素(UR)、蛋白胨(PE)和酵母抽提物(TE)等5种氮源对乳酸浓度和糖利用率的影响,上述5种氮源的添加量均以N浓度计(3.5g/L)。
筛选出适宜的氮源种类后,在相同碳源(同上)和氮源为氯化氨(AC添加量以N浓度计为3.5g/L)条件下,进行乳酸发酵工艺参数的单因素和正交优化实验。
b.pH对乳酸发酵的影响
固定CaCO3添加量70g·L-1,发酵时间96h,接种量2.5%,研究不同pH(4.5、5.0、5.5、6.0和6.5)对米根霉发酵的影响。
c.接种量对乳酸发酵的影响
固定CaCO3添加量70g·L-1,pH6.0,发酵时间96h,研究不同接种量(1.0%、1.5%、2.0%、2.5%和3.0%)对米根霉发酵的影响。
d.发酵时间对乳酸发酵的影响
固定CaCO3添加量70g·L-1,pH6.0,接种量2.5%,研究发酵时间(12、24、36、48、60、72、84、96和108h)对米根霉发酵的影响。
e.CaCO3添加量对乳酸发酵的影响
固定pH6.0,接种量2.5%,发酵时间96h,研究CaCO3添加量(40、50、60、70和80g/L)对米根霉发酵的影响。
1.3.4.2 正交实验优化设计 在单因素实验的基础上,选取CaCO3添加量、pH、接种量和发酵时间4个因素进行L9(34)正交实验,各取3个水平,优化发酵参数,并进行验证实验。

表1 正交实验因素水平表Table 1 Factors and levels of the orthogonal tests
1.4 数据统计分析
实验数据表示为平均值±标准差,采用SPSS软件处理,置信水平为95%时,p<0.05说明数据在统计学上存在显著性差异。
2 结果与讨论
2.1 米根霉糖代谢特性研究
木质纤维素的降解产物是以葡萄糖和木糖为主的混合糖,为更好地将其作为发酵基质,研究了米根霉发酵葡萄糖、木糖或混合糖(葡萄糖∶木糖=1∶1)的糖代谢情况。由图1可见,葡萄糖或木糖单独作为碳源时的糖利用率和糖酸转化率基本相同,但混合糖发酵的糖酸转化率和糖利用率均明显提高,尤其糖酸转化率成倍增长。因为,二者代谢途径存在差异,葡萄糖代谢主要与糖酵解途径相关,纯葡萄糖培养条件下,不断下降的体系pH对菌体生长产生了抑制作用,使菌体生物量偏低,造成糖利用率和糖酸转化率偏低;而米根霉代谢木糖时会产生较高的胞内还原力(NADH/NAD+)和ATP,代谢反应以促进细胞生物量的合成以及细胞大分子组分的合成为主,指向生物量积累的能量代谢旺盛[12-13]。另一方面,与葡萄糖相比较,米根霉发酵木糖所需要的C/N比较低,积极有效的呼吸作用比葡萄糖基质更重要,而木糖代谢过程中的氧化还原途径与较高的呼吸通量有关系[14]。因此,米根霉R.oryzaeLS-1不仅可以发酵葡萄糖和木糖,且二者共存时还可以提高糖酸转化率,对乳酸发酵具有协同促进作用。

图1 不同碳源条件下的米根霉糖代谢特性Fig.1 Sugar metabolism of Rhizopus oryzae LS-1 for different carbon sources
2.2 米根霉发酵丢糟水解液产乳酸工艺研究
2.2.1 氮源种类对丢糟水解液发酵乳酸的影响 氮源不仅对微生物合成细胞物质至关重要,而且是一种成本较高的培养基组分。为了降低发酵成本,研究了硫酸铵(AS)、氯化氨(AC)、尿素(UR)、蛋白胨(PE)和酵母抽提物(TE)五种氮源对乳酸发酵的影响。氮源量均以N浓度计(3.5g/L)。
由图2可知,无氮源添加时,乳酸浓度仅为2.52 g·L-1,糖利用率为12.35%;添加氮源后均能不同程度地提高乳酸浓度和糖利用率。因为丰富的N源为生物大分子如蛋白质、脂肪等提供了充足的原料,从而促进米根霉糖酸转化率的提高。然而,不同氮源对乳酸发酵的影响各有差异,其中氯化铵对应的乳酸浓度和糖利用率最高,说明与其它四种氮源相比,氯化铵能更多地促进乳酸合成。该结果与硫酸铵是最适宜氮源的研究结果不一致,这可能是由于培养基成分不同所致[15],因为培养基营养成分会通过影响乳酸菌形态来影响发酵过程和乳酸产量。另外,总体比较而言,尽管有机氮源富含矿物质、维生素和糖分,但无机氮源更有利于促进乳酸生成。从培养基成本和氮源作用效果等角度衡量,本实验选用氯化铵作为培养基中的氮源。

图2 氮源种类对乳酸发酵的影响Fig.2 Effects of nitrogen sources on the fermentation of lactic acid
2.2.2 pH对丢糟水解液发酵乳酸的影响 微生物生存环境中的pH对微生物的活动影响很大,主要作用在于引起细胞膜电荷变化,从而影响微生物对营养物质的吸收;影响代谢过程中酶的活性;改变生长环境中营养物质的可给性以及有害物质的毒性。因此,适宜的pH能使菌体获得最佳培养效果。由图3可以看出,随着体系pH不断升高,乳酸浓度和总糖利用率均逐渐增加,说明pH升高有利于乳酸的生成。当pH为6.0时,乳酸产量最高为10.82g/L,对应的总糖利用率也达到峰值75.33%。当pH升至6.5时,乳酸浓度显著降低,总糖利用率也随之减小,这可能是长时间高pH环境严重影响了菌体的生长及发酵产酸过程。但另一方面,发酵过程中产生的乳酸也会造成体系pH自动下降,对糖利用效率和乳酸的合成产生较大影响。因此,适当高的初始pH不仅能缩短菌体生长的迟滞期,而且还能使发酵体系维持较长时间的良好环境,避免过早发生产物抑制现象,有利于米根霉的发酵代谢。故发酵初始pH确定为6.0,这与Martak J等[16]米根霉生长最适pH范围5.0~6.0的研究结果相一致。
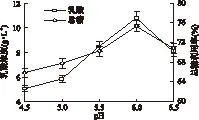
图3 初始pH对乳酸浓度和总糖利用率的影响Fig.3 Effects of pH on lactic acid concentration and total sugars utilization
2.2.3 接种量对丢糟水解液发酵乳酸的影响 接种量是由发酵时菌体生长繁殖的速度决定的,通常较大的接种量可以缩短生长周期,使产物合成提前,但接种量过大也可能使菌体生长过快,造成溶氧量不足,从而影响产物合成。由图4可知,随着接种量的增加,乳酸浓度和总糖利用率均逐渐升高,二者变化趋势基本一致,当接种量为2.5%时的乳酸浓度和总糖利用率均达到最大值,分别为9.06g/L和62.90%,当接种量提高到3.0%时,二者反而降低。实验过程中观察发现,接种量的大小影响菌体形态。随着接种量的不断提高,菌丝体越易聚集成较大的絮状菌团。因为过高的接种量容易导致过高的菌丝体生长密度,进而导致发酵液粘度过高,而相对较低的搅拌转速(220r/min)所产生的剪切力,也不足以克服菌丝之间由Ca2+介导的静电引力,从而使得菌丝体聚集成较大的絮状菌团。絮状菌团导致发酵体系的传质效率降低,乳酸产量下降。另一方面,尽管过高的接种量会缩短延迟期时间,菌体进入快速生长期,但菌体大量繁殖,也会产生过多的代谢副产物,反而抑制乳酸的形成。Bai等[17]研究发现,接种量过小,米根霉呈现丝状菌丝形态,接种量过高,菌丝发生聚集形成菌块,造成氧气和营养成分的传质受限引起菌会内部发生自溶现象。因此,选定接种量为2.5%。
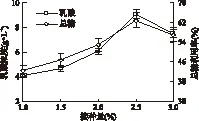
图4 接种量对乳酸浓度和总糖利用率的影响Fig.4 Effects of inoculum size on lactic acid concentration and total sugars utilization
2.2.4 发酵时间对丢糟水解液发酵乳酸的影响 由图5可知,发酵开始96h内,总糖利用率和乳酸浓度逐渐增加,分别提高了75.86%和52.36%;总糖浓度下降明显,说明乳酸主要在总糖不断消耗的过程中产生。随着时间的推移,一方面营养物质消耗殆尽,菌体早衰,发酵后劲不足;另一方面,大的菌球颗粒或聚集也会引起传氧传质阻力急剧增加,从而影响乳酸产量和总糖利用率。故确定发酵时间为96h。

图5 发酵时间对乳酸浓度和总糖利用率的影响Fig.5 Effects of fermentation time on lactic acid concentration and total sugars utilization
2.2.5 CaCO3添加量对丢糟水解液发酵乳酸的影响 乳酸发酵过程中,发酵产生的乳酸会使反应体系pH快速下降,较低的pH会对菌体正常生长和产酸过程产生强烈抑制,需加入一定的中和剂来控制pH。CaCO3是最为常用的中和剂,作用温和,不会对细胞造成损伤,有利于菌体生长。同时,溶解释放的Ca2+在菌体生长和菌球形成过程起重要作用,适量Ca2+有利于米根霉菌丝体的生长及菌球的形成。由图6可知,未添加CaCO3时,生成的乳酸浓度仅为2.12g/L,总糖利用率为20.83%,可能受到低pH和乳酸反馈抑制的影响。当CaCO3添加量为40g/L时,乳酸浓度迅速上升至5.82g/L,总糖利用率也提高至60.47%。当CaCO3添加量由40g/L提高到70g/L时,乳酸浓度继续保持高增长,达到10.62g/L,且增幅高于总糖利用率,说明该范围更有利于乳酸浓度的积累,同时观察到该阶段的米根霉形态为大小均一的小球体,这种形态有利于促进乳酸生成量的提高[1],而且Ca2+对乳酸脱氢酶(LDH)有激活作用,可从一定程度上加快乳酸的生成[18],说明CaCO3的适当增加能提高菌体对糖的利用效率和乳酸积累。随着CaCO3添加量的继续增加(80g/L),乳酸浓度反而下降,总糖利用率仅微幅增长,基本保持恒定。因为CaCO3的过量添加会对菌体造成包埋,不利于菌体与培养基接触而影响发酵进程,影响溶氧传递系数,甚至造成局部厌氧环境,导致乙醇等副产物的生成[17]。因此,适量添加CaCO3会促进菌体繁殖生长和乳酸生成。综合考虑pH调节效果、乳酸产量和菌体形态等因素,确定CaCO3添加量为70g/L。

表2 正交实验设计与结果Table 2 L9(34) Orthogonal design and experimental results
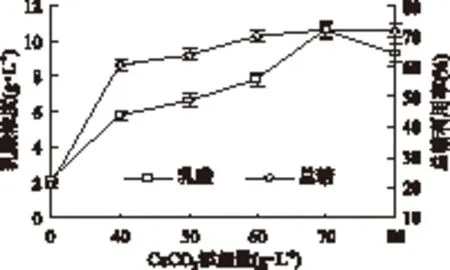
图6 CaCO3添加量对乳酸浓度和总糖利用率的影响Fig.6 Effects of calcium carbonate addition on lactic acid concentration and total sugars utilization
2.3 正交实验优化
在上述单因素实验结果基础上,选取一定区间内对米根霉发酵产乳酸效果影响较显著的CaCO3添加量、pH、接种量和发酵时间4个因素作为研究对象,选取乳酸浓度和总糖利用率2个评价指标进行分析,结果见表2~表4。
从表2可以看出,各因素对乳酸浓度的影响大小顺序为D>C>A>B,最优组合为 A2B3C3D2;各因素对总糖利用率的影响大小顺序为C>A>D>B,最优组合为A2B3C3D2,不同指标条件下的最优组合一致。从表3和表4方差分析可知,上述四个因素对乳酸浓度和总糖利用率的影响均不显著。
为了更准确判定最优发酵参数,选取表2中正交最优组合A2B3C3D2条件进行五次重复实验(取平均值),发现该条件下的乳酸浓度和总糖利用率分别为12.47g/L和76.92%,略低于表2中的直观优组合7(A3B1C3D2)。鉴此,再次选择这两个组合(A3B1C3D2和A2B3C3D2)进行五次重复验证实验(取平均值)。结果发现,组合A2B3C3D2条件对应的乳酸浓度和总糖利用率较高,分别为13.27g/L和79.61%,高于组合A2B3C3D2条件对应的12.81g/L和77.58%,但二者差异不显著(p<0.05)。
综合上述优化实验结果,从发酵参数控制、提高乳酸浓度和总糖利用率等角度考虑,确定最优发酵条件为A2B3C3D2,即CaCO3添加量80g/L、pH6.5、接种量3.0%和发酵时间96h。

表3 乳酸浓度的影响方差分析Table 3 Variance analyses statistical data on the factors of lactic acid concentration
注:p<0.05表示差异性显著(下同)。

表4 总糖利用率的影响方差分析Table 4 Variance analyses statistical data on the factors of the reducing sugars utilization rate
3 结论
米根霉诱变菌株R.oryzaeLS-1的糖代谢研究表明,米根霉R.oryzaeLS-1能分别代谢利用葡萄糖和木糖生产乳酸,且二者共存时对发酵过程具有协同促进作用,能显著提高乳酸生成浓度和糖酸转化率,这就为利用木质纤维素中的木糖和葡萄糖发酵生产乳酸提供了理论依据。利用LS-1米根霉发酵丢糟水解液实验表明,发酵过程中选取氯化铵作为氮源最为适宜,在接种量为3.0%、pH为6.5、发酵时间为96h、CaCO3添加量为80g/L的条件下,乳酸发酵效果最好,浓度达到13.27g/L,糖利用率为79.61%。表明米根霉R.oryzaeLS-1具备发酵丢糟水解液制备乳酸的潜力,这也为丢糟生物质的转化利用奠定了理论基础。
[1]HongWei Yen,TsiaJu Chen,WeiChin Pan,etal. Effects of neutralizing agents on lactic acid production byRhizopusoryzaeusing sweet potato starch[J]. World J Microbiol Biotechnol,2010,26(3):437-441.
[2]杨登峰,潘丽霞,关妮,等.产高纯度L-乳酸大肠杆菌基因工程菌的初步研究[J].现代食品科技,2010,26(2):126-128,171.
[3]Kenji Okano,Tsutomu Tanaka,Chiaki Ogino,etal. Biotechnological production of enantiomeric pure lactic acid from renewable resources:recent achievements,perspectives,and limits[J]. Appl Microbiol Biotechnol,2010,85(3):413-423.
[4]邹水洋,郭祀远,肖凯军.生物转化木质纤维素原料生产乳酸的研究进展[J].现代食品科技,2008,24(4):394-400.
[5]Katsuichi Saito,Yasuhiro Hasa,Hideyuki Abe. Production of lactic acid from xylose and wheat straw byRhizopusoryzae[J]. Journal of Bioscience and Bioengineering,2012,114(2):166-169.
[6]Zhangwei Xue,Limin Wang,Jiansong Ju,etal. Efficient production of polymer-grade L-lactic acid from corn stover hydrolyzate by thermophilicBacillussp. strain XZL4[J]. Springer Plus,2012,1:43.
[7]Dong-Mei Bai,Shi-Zhong Li,Z. Lewis Liu,etal. Enhanced L(+)-Lactic acid production by an adapted strain ofRhizopusoryzaeusing corncob hydrolysate[J]. Appl Biochem Biotechnol,2008,144(1):79-85.
[8]Adenise L. Woiciechowski,Carlos R. Soccol,Luis P. Ramos,etal. Experimental design to enhance the production of L-(+)-lactic acid from steam-exploded wood hydrolysate usingRhizopusoryzaein a mixed-acid fermentation[J].Process Biochemistry,1999,34(9):949-955.
[9]Enoch Y. Park,Pham Ngoc Anh,Naoyuki Okuda. Bioconversion of waste office paper to L(+)-lactic acid by the filamentous fungusRhizopusoryze[J]. Bioresource Technology,2004,93(1):77-83.
[10]郭洋,闫巧娟,江正强,等.米根霉利用蔗糖和蔗渣发酵生产乳酸的研究[J].食品与发酵工业,2009,35(12):24-27.
[11]任海伟,张飞,张轶,等. 白酒糟水解液摇瓶发酵生产木糖醇的工艺优化[J].农业工程学报,2012,28(10):287-292.
[12]余洋,徐晴,李霜.米根霉在氮源限制下的木糖代谢和关键酶特性[J].微生物学报,2013,53(11):1189-1194.
[13]窦畅,徐晴,宋萍,等.米根霉利用木糖与葡萄糖的代谢差异[J].微生物学报,2011,51(4):468-473.
[14]Ronald H W Maas,Jan Springer,Gerrit Eggink,etal. Xylose metabolism in the fungusRhizopusoryzae:effect of growth and respiration on L(+)-lactic acid production[J]. J Ind Microbiol Biotechnol,2008,35(6):569-578.
[15]Wanying Yao,Xiao Wu,Jun Zhu,etal. Miller. Utilization of protein extract from dairy manure as a nitrogen source byRhizopusoryzaeNRRL-395 for L-lactic acid production[J].Bioresource Technology,2010,101(11):4132-4138.
[16]Ján Marták,tefan Schlosser,Erika Sabolov,etal. Fermentaion of lactic acid with Rhizopus arrhizus in a stirred tank reactor with a periodical and feed operation[J]. Process Biochemistry,2003,38(11):1573-1583.
[17]Dong-Mei Bai,Min-Ze Jia,Xue-Ming Zhao,etal. L(+)-
Production of lactic acid from distillers grains hydrolysates byRhizopusoryzaeLS-1
TIAN Yan-hua1,REN Hai-wei2,*,LIU Na-li1
(1.Food Engineering Department of Shanxi pharmaceutical Vocational College,Taiyuan 030031,China;2.School of Life Science and Engineering,Lanzhou University of Technology,Lanzhou 730050,China)
Distillers grains,one of the lignocellulosic biomasses riched in cellulose and hemicellulose,was used as an economically attractive carbohydrate feedstock for production of lactic acid by saccharification and microbial fermentation processes. The aim of this study was to investigate the possibility of lactic acid production from distillers grains hydrolysates by the mutant strainRhizopusoryzaeLS-1 and to optimize the biological conversion of reducing sugars into lactic acid to evaluate the culture conditions. The effects of factors such as inoculations size,CaCO3addition,pH value and fermentation time on the lactic acid concentration and the reducing sugars utilization rate were researched by the method of orthogonal experimental design. In addition,the sugar metabolism ofR.oryzaeLS-1 was also studied. The results show thatR.oryzaeLS-1 has the capability to utilize xylose or glucose as carbon resource,furthermore there are synergic and complementary actions during the coexistence of both sugars. The cooperated operations of metabolism was beneficial to the bioconversion of lignocellulosic biomass for lactic acid production. Fermentation results show that ammonium chloride is the most favorable nitrogen source for lactic acid production,the optimal fermentation conditions are inoculation size of 3.0% seed culture,CaCO3addition of 80g·L-1,fermentation time of 96h and culture pH of 6.5. Under optimal conditions,13.27g/L lactic acid was produced and the sugar conversion rate reached 79.61%. This study provided an encouraging means of producing lactic acid from lignocellulosic resource such as the low-cost distillers grains. It was concluded that the fermentation technics for the production of lactic acid from distiller grains was worthy to be developed on a large scale.
Rhizopusoryzae;distiller grains hydrolysates;lactic acid fermentation
2014-03-04
田艳花(1981-),女,硕士,讲师,研究方向:食品生物技术。
*通讯作者:任海伟(1983-),男,硕士,讲师,研究方向:农副产物资源转化利用。
国家自然科学基金项目(51366009);甘肃省杰出青年科学基金项目(1210RJDA016);兰州理工大学“红柳青年教师培养计划”项目(Q201207)。
TS209
A
1002-0306(2015)01-0192-07
10.13386/j.issn1002-0306.2015.01.031
猜你喜欢
阅读(中年级)(2022年9期)2022-10-08
红蜻蜓·低年级(2022年2期)2022-03-19
红蜻蜓·低年级(2021年12期)2021-12-19
科技与创新(2016年22期)2017-03-30
科技视界(2016年20期)2016-09-29
西南农业学报(2016年5期)2016-05-17
特产研究(2016年3期)2016-04-12
中国酿造(2016年12期)2016-03-01
广东饲料(2016年8期)2016-02-27
中国酿造(2014年9期)2014-03-11
