
乙型肝炎病毒感染过程中的免疫应答
2015-06-01 10:16何佳奇路立业黄敦武
安徽医学 2015年3期
何佳奇 路立业 黄敦武
乙型肝炎病毒感染过程中的免疫应答
何佳奇 路立业 黄敦武
乙型肝炎病毒;固有免疫;特异性免疫
肝炎病毒感染,特别是乙型肝炎病毒(hepatitis B virus,HBV)和丙型肝炎病毒(hepatitis C virus,HCV)感染给人类健康带来了严重的威胁。目前全球约有3.5亿的HBV感染者和约1.8亿的HCV感染者,约57%的肝硬化患者和78%的原发性肝癌患者和HBV或HCV慢性感染有关[1]。我国作为HBV感染的高发国家,研究HBV的每个致病环节至关重要,本文主要针对成人HBV感染过程中的免疫应答的现有研究进行综述,并探讨HBV和宿主免疫应答之间的动态关系。
HBV是嗜肝病毒科正嗜肝病毒属的一员。在电镜下观察,HBV感染者血清中存在3种形式的颗粒:大球形颗粒(Dane颗粒)、小球形颗粒和管形颗粒。大球形颗粒为完整的HBV颗粒,是病毒复制的主体;后两者为HBsAg组成的空心包膜,无传染性。HBV基因结构独特而精密,由不完全的环状双链DNA组成,复制过程非常特殊。在HBV感染的肝细胞核内有稳定的共价闭合环路DNA(covalently closed circular DNA,cccDNA),其复制过程中有一个反转录步骤,当HBV进入肝细胞后,HBV DNA进入细胞核形成cccDNA,然后以cccDNA为模板合成前基因组mRNA,前基因组mRNA进入胞浆作为模板合成负链DNA,再以负链DNA为模板合成正链DNA,经过外膜包装后出胞完成复制过程[2]。
HBV感染的自然病程是复杂多变的,同时受到很多因素的影响,包括病毒因素(HBV基因型、病毒变异和病毒复制的水平)、宿主因素(性别、年龄、免疫状态)和其他外源性因素(同时感染其他嗜肝病毒或HIV、嗜酒等)。临床上HBV感染包括从症状不明显的肝炎到急性有症状的肝炎,甚至急性爆发型肝炎,从非活动性HBsAg携带状态到慢性肝炎、肝硬化等各种状况,有15%~40%的慢性HBV感染者会发展为肝硬化和晚期肝病[3]。母婴传播导致的新生儿HBV感染者,约90%会发展成为慢性感染,一般经历免疫耐受、免疫清除和非活动或低(非)复制阶段。而青少年和成人期感染HBV者,仅5%~10%发展成为慢性,一般无免疫耐受期[4]。本文综述内容主要针对成人感染HBV自然病程中的免疫应答。
1 急性HBV感染患者的免疫应答

Kupffer细胞是肝内一个非常庞大的细胞群,几乎占到肝细胞总数的15%[9]。在给HBV转基因小鼠注射一种肝脏特异性的疟疾病原株,大量的Kupffer细胞被激活,小鼠体内的疟原虫和HBV感染均得到一定的控制[10]。因此其在肝内固有免疫和诱导特异性免疫方面的作用不容小视。另外Kupffer细胞可以通过分泌细胞因子IFN-γ和趋化因子CXCL9、CXCV10来诱导HBV特异性T细胞的聚集和分化[11]。
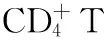
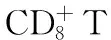
在动物实验中,被HBV感染的黑猩猩体内至少90%的病毒是在肝脏损伤出现前被清除的。在给两只成年黑猩猩注入HBV血清后,猩猩体内的病毒DNA载量都在第8周达到高峰,然后迅速下降。而两只黑猩猩血清ALT在10~12周时才开始上升,分别在16周和21周达到高峰[19]。可见非溶细胞作用的病毒清除机制在宿主清除体内HBV病毒过程中占据着十分重要的作用。而且该实验还发现在急性HBV感染时,这种非溶细胞病毒清除机制也能有效控制HBV cccDNA。这种非溶细胞病毒清除机制则是宿主清除HBV的最理想状态,即在不损伤自身重要器官的同时清除体内的病毒。
1.2.4 B淋巴细胞和体液免疫 B淋巴细胞受HBV抗原刺激后,分化为浆细胞,产生大量特异性抗体,介导体液免疫应答,并通过分泌细胞因子,参与免疫调节[23]。虽然体液免疫也是一种非溶细胞免疫反应,其不同于非溶细胞病毒清除机制(可以清除肝细胞内病毒),体液免疫只能清除循环中的HBV,而对肝细胞内的HBV无效。中和性抗体可以阻止病毒侵犯正常的靶细胞,HBV感染者体内的中和性抗体主要是抗HBsAg和抗PreS1,它们可以长期的控制自限性HBV感染者体内的痕量HBV,并保护这些宿主免于再次感染HBV。虽然抗HBsAg在感染的早期已经产生,但因中和大量的HBsAg,所以在感染早期不易被检测。普遍认为抗HBeAg和抗HBcAg没有中和作用,不能保护宿主再受侵犯。病毒抗原的中和抗体和非中和抗体都可以和吞噬细胞表面的Fc受体结合来调节这些吞噬细胞的抗原递呈能力。另外,抗HBsAg和抗PreS1还可以激活补体系统来参与宿主的免疫反应。但是大量免疫复合物的形成可以使部分HBV感染者出现肝外损伤,如急性乙型肝炎早期出现的血清病样表现,慢性乙型肝炎患者中发生的膜性肾小球肾炎伴肾病综合征、结节性多动脉炎等[24]。

2 慢性HBV感染患者的免疫应答


慢性HBV感染患者的免疫功能,特别是特异性T细胞的功能和数量都较正常人明显受损,Treg明显增多,抑制性免疫机制异常活跃。所以在多数患者体内存在持续的HBV复制,同时伴有肝组织中大量非特异性淋巴细胞侵润(这可能是慢性HBV感染者引起肝脏损伤的主要原因),导致肝脏组织炎症及肝细胞坏死、再生,并逐渐出现肝纤维化等病理变化,仅有个别HBV感染者可以通过自身免疫清除体内的HBV,大多数患者都逐渐演变为终末期肝病。
[1] Barbara R. Pathogenesis of chronic viral hepatitis: differential roles of T cells and NK cells[J]. Nat Med,2013,19(7):859-868.
[2] Don G, Alfred MP. Hepatitis B virus infection-natural history and clinical consequences[J]. N Engl J Med, 2004,350(11):1118-1129.
[3] Fattovich G. Natural history and prognosis of hepatitis B[J].Semin Liver Dis,2003,23(1):47-58.
[4] Su WJ,Liu CC,Liu DP,et al.Effect of age on the incidence of acute hepatitis B after 25 years of a universal newborn hepatitis B immunization program in Taiwan[J].J Infect Dis, 2012,205(5):757-762.
[5] Samuel CE.Antiviral actions of interferons[J].Clin Microbio Rev,2001,14(4):778-809.
[6] Cui GY,Diao HY.Recognition of HBV antigens and HBV DNA by dendritic cells[J]. Hepatobiliary Pancreat Dis Int,2010,9(6):584-592.
[7] Kimura K,Kakimi K,Wieland S,et al.Activated intrahepatic antigen-presenting cells inhibit hepatitis B virus replication in the liver of transgenic mice[J].J Immunol,2002,169(9): 5188-5195.
[8] George K,Vassilis V,Elias K.Role of Kupffer cells in the pathogenesis of liver disease[J]. World J Gastroenterol,2006,12(46): 7413-7420.
[9] Pasquetto V,Guidotti LG,Kakimi K,et al.Host-virus interactions during malaria infection in hepatitis B virus transgenic mice[J].J Exp Med,2000,192(4):529-536.
[10] Kakimi K,Lane TE,Chisari FV,et al.Cutting edge: inhibition of hepatitis B virus replication by activated NK T cells does not require inflammatory cell recruitment to the liver[J]. J Immunol,2001,167(12): 6701-6705.
[11] Bray SM,Vujanovic L,Butterfield LH.Dendritic cellvbased vaccines positively impact natural killer and regulatory T cells in hepatocellular carcinoma patients[J].Clin Dev Immunol,2011;2011:249281.
[12] Mizukoshi E,Sidney J,Livingston B,et al.Cellular immune responses to the hepatitis B virus polymerase[J].J Immunol,2004,173(9): 5863-5871.
[13] Guidotti LG,Chisari FV. Immunobiology and Pathogenesis of Viral Hepatitis[J]. Annu Rev Pathol, 2006, 1:23-61.
[14] Thimme R,Wieland S,Steiger C,et al.CD8(+) T cells mediate viral clearance and disease pathogenesis during acute hepatitis B virus infection[J].J Virol,2003,77(1):68-76.
[15] Zhang JY,Zhang Z,Lin F,et al.Interleukin-17-producing CD4(+) T cells increase with severity of liver damage in patients with chronic hepatitis B[J].Hepatology,2010,51(1):81-91.
[16] Wijaya I,Hasan I.Reactivation of hepatitis B virus associated with chemotherapy and immunosuppressive agent[J].Acta Med Indones,2013, 45(1): 61-66.
[17] Sprengers D,Molen RG,Kusters JG,et al.Analysis of intrahepatic HBV-specific cytotoxic T-cells during and after acute HBV infection in humans[J].J Hepatol,2006,45(2):182-189.
[18] Han SH.Extrahepatic manifestations of chronic hepatitis B[J].Clin Liver Dis,2004,8(2):403-418.
[19] Duan XZ,Wang M,Li HW,et al.Decreased frequency and function of circulating plasmocytoid dendrtic cells(pDC) in hepatitis B virus infected humans[J].J Clin Immunol,2004,24(6):637-646.
[20] Ciurea A,Hunziker L,Klenerman P, et al.Impairment of CD4(+) T cell responses during chronic virus infection prevents neutralizing antibody responses against virus escape mutants[J]. J Exp Med,2001,193(3):297-305.
[21] Carolina B,Paola F,Caterina V,et al.Characterization of Hepatitis B Virus (HBV)-Specific T-Cell Dysfunction in Chronic HBV Infection[J].J Virol,2007,81(8):4215-4225.
[22] Rigopoulou EI,Suri D,Chokshi S,et al.Lamivudine plus interleukin-12 combination therapy in chronic hepatitis B: antiviral and immunological activity[J]. Hepatology,2005,42(5):1028-36.
[23] Wherry EJ.T cell exhaustion[J].Nat Immunol,2011,12(6): 492-499.

[25] Seeger C,Mason WS.Hepatitis B virus biology[J].Microbiol Mol Biol Rev,2000, 64(1): 51-68.
[26] Milich D,Liang TJ.Exploring the biological basis of hepatitis B e antigen in hepatitis B virus infection[J].Hepatology,2003,38(5):1075-1086.
[27] Anthony TT,Sarene K,Vera G,et al.Understanding the immunopathogenesis of chronic hepatitis B virus: an Asian prospective[J].J Gastroen Hepatol,2008,23(6):833-843.
[28] Schurich A,Khanna P,Lopes AR,et al.Role of the coinhibitory receptor cytotoxic T lymphocyte antigen-4 on apoptosis-prone CD8 T cells in persistent hepatitis B virus infection[J]. Hepatology,2011,53(5):1494-1503.
[29] Gu XB,Yang XJ,W JH.PD-1 expression on CTL may be related to more severe liver damage in CHB patients with HBV genotype C than in those with genotype B infection[J].J Viral Hepat,2013,20(4):e1-2.
[32] Das A,Matthew H,Nathan D,et al.Functional skewing of the global CD8 T cell population in chronic hepatitis B virus infection[J].J Exp Med,2008,205(9):2111-2124.
[33] Duramad O,Fearon KL,Chan JH,et al.IL-10 regulates plasmacytoid dendritic cell response to CpG-containing immunostimulatory sequences[J].Blood,2003,102(13):4487-4492.
(2014-11-10 收稿 2014-12-25 修回)
答题卡填写后沿此虚线剪下寄回(答题卡复印无效)

2015年《安徽医学》继续医学教育答题卡(1-3期)
注:学习3期为1个单元(3分),全年共12分。如答题卡空间不够,可附页答题;作者寄答案时,请详细写出联系电话、联系地址。
110003 辽宁沈阳 中国人民解放军第202医院传染科
路立业,llyjfj202@163.com
10.3969/j.issn.1000-0399.2015.03.042
猜你喜欢
传染病信息(2022年2期)2022-07-15
南京师大学报(自然科学版)(2022年2期)2022-06-20
中国典型病例大全(2022年13期)2022-05-10
昆明医科大学学报(2022年2期)2022-03-29
昆明医科大学学报(2022年1期)2022-02-28
小学生时代·综合版(2016年8期)2016-05-14
小星星·阅读100分(高年级)(2015年11期)2015-11-28
大自然探索(2015年11期)2015-09-10
医学研究杂志(2015年12期)2015-06-10
初中生世界·八年级(2009年8期)2009-11-03
