
红花种质资源形态性状遗传多样性分析
2015-04-29 05:10郭丽芬张跃徐宁生高梅李文昌胡尊红邓剑川刘旭云
热带作物学报 2015年1期
郭丽芬 张跃 徐宁生 高梅 李文昌 胡尊红 邓剑川 刘旭云

摘 要 为促进红花品种改良及种质资源的高效利用,以筛选出的68份优异种质为材料,对19个形态性状进行调查分析,研究其遗传多样性。利用主成分分析和聚类分析,结果表明:国内外不同地理来源红花资源群体间的遗传多样性丰富,数量性状上存在较大变异。主成分分析以5个主成分因子反映了13个数量性状的全部信息。累计贡献率达85.08%,各主成分包含的信息具有一定的相关性,表明主成分分析结果与参试资源和性状指标的选择均有关系。将主要农艺性状归纳为产量构成因子,生长势因子和子粒因子。采用DPS 7.0软件,进行系统聚类分析,根据19个性状在红花种质资源间的不同表现,在遗传距离13.29处将68份供试材料聚为7大类群。聚类结果表明,不同地理来源的种质资源多样性较丰富,聚类分析中部分地理来源相同或遗传背景相似的资源能够聚在同一类群,但也有一些地理来源及遗传背景不一致的种质资源也聚在同一类群,少部分资源品种表现差异大,表明品种类群间的性状分化与地理分布既有一定的联系又不绝对相关,即不同地区的材料聚类呈现一定的地理分布规律。研究结果表明:供试的红花种质资源具有丰富的遗传多样性,其中多样性指数最高的是分枝总数为15.779 2,其次为单株有效果球数、单株产量和分枝长度,依次为15.673 9、10.617 5和10.415 5,多样性指数越高,表明遗传多样性越丰富。实践证明在杂交或其他方法育种中,选配亲本材料应依据主成分的排序,具体分析与全面评价每个亲本材料综合指标的优劣,依据红花育种目标,结合聚类分析,合理地选配组合。对于红花品种改良及资源的进一步开发利用具有重要意义。
关键词 红花;形态性状;主成分分析;聚类分析;遗传多样性
中图分类号 S681.9 文献标识码 A
The Genetic Diversity Analysis of Both Morphologic
Traits and Agrinomic Traits of Safflower
(Carthamus tinctorius L.)Germplasm Resources
GUO Lifen1, ZHANG Yue2, XU Ningsheng1, GAO Mei1, LI Wenchang1*,
HU Zunhong1, DENG Jianchuan1, LIU Xuyun1*
1 Industrial Crop Research Institute, Yunnan Academy of Agricultural Sciences, Kunming,Yunnan 650205, China
2 Shuanglong Subdistrict Office, Panlong, Kunming, Yunnan 650224, China
Abstract Both morphologic traits and agronomic traits of 68 outstanding safflower varieties were evaluated by both principal component analysis and cluster analysis to promote the breeding of new cultivars and effective application of germplasms resources. The genetic diversity of safflower evaluated was very rich, and the variation in quantitative character among the varieties was great. Five principal component could reflect fully 13 quantitative characters with cumulative rate 85.08%. The results of the principal component analysis correlated with morphologic traits and agronomic traits evaluated. The agronomic traits were divided into 3 groups: yield component factors, growing factors, kernel factors. 68 safflower varieties were divided into 7 groups by cluster analysis of 19 traits at 13.29 genetic distance by software DPS 7.0. There were plenty of variation among varieties from different places. Some varieties from the same place could be divided into the same group, in which there were some varieties with huge difference from different places. It meant morphology cluster was somewhat related to the original place. The trait with the highest index of genetic diversity was the number of branches(15.779 2), the effective number of fruits per plant(15.673 9), the yield per plant(10.617 5) and the length of branches(10.415 5). It is important to select parents with good quality by the results of both the principal component analysis and the analysis to breed new cultivars, which will be helpful for both the application of safflower germplasm resources and the breeding of new safflower cultivars.
Key words Safflower(Carthamus tinctorius L.);Morphologic traits;Principal component analysi;Cluster analysis;Genetic diversity analysis.
doi 10.3969/j.issn.1000-2561.2015.01.014
红花(Carthamus tinctorius L. 2n=24)属菊科( Compositae)红兰花属(Carthamus),又名黄兰、红兰花、草红花、红花菜,为一年生草本双子叶植物。红花的原产地为大西洋东部,非洲西北部的加那利群岛及地中海沿岸[1]。种植区分布在20°S~47°N的范围之间,在沿海地带和半干旱地区均有分布[2]。红花在我国已有2 100多年的栽培和药用历史,主要集中在新疆、四川、云南、河南、甘肃、河北、山东、浙江、江苏等省(区)。中国红花种植面积约6.6万hm2,主要是药用,部分油药兼用。早在20世纪30年代,甘肃省敦煌县就将红花作为油料作物栽培,直到20世纪50年代,由于推广高含油量的优良品种,红花才成为一种新的油料作物得以发展。在国外,红花是一种重要的油料作物,红花子油是世界公认的具有保健、美容功效的功能性食用油,在国际上被称为“绿色食品”,其亚油酸含量是已知植物油中最高的,达80%,号称“亚油酸之王”,并富含维生素E和其他营养成分,长期食用具有良好的保健作用[3-5]。
用主成分分析法和聚类分析法是遗传育种和品种资源研究中普遍应用的两种方法[6],已在豌豆[7]、油菜[8]、黍稷[9]、蚕豆[10]等种质资源研究中得到广泛应用。目前,关于红花种质资源形态性状遗传多样性分析的报道较少,本研究通过对世界不同地理来源的红花种质资源进行系统的鉴定,对其主要形态性状进行遗传多样性分析,为充分了解红花种质资源遗传多样性地理分布特点和种质资源群间的遗传关系,挖掘优异种质材料,旨在为拓宽红花品种改良与种质资源创新基础提供参考依据。
1 材料与方法
1.1 材料
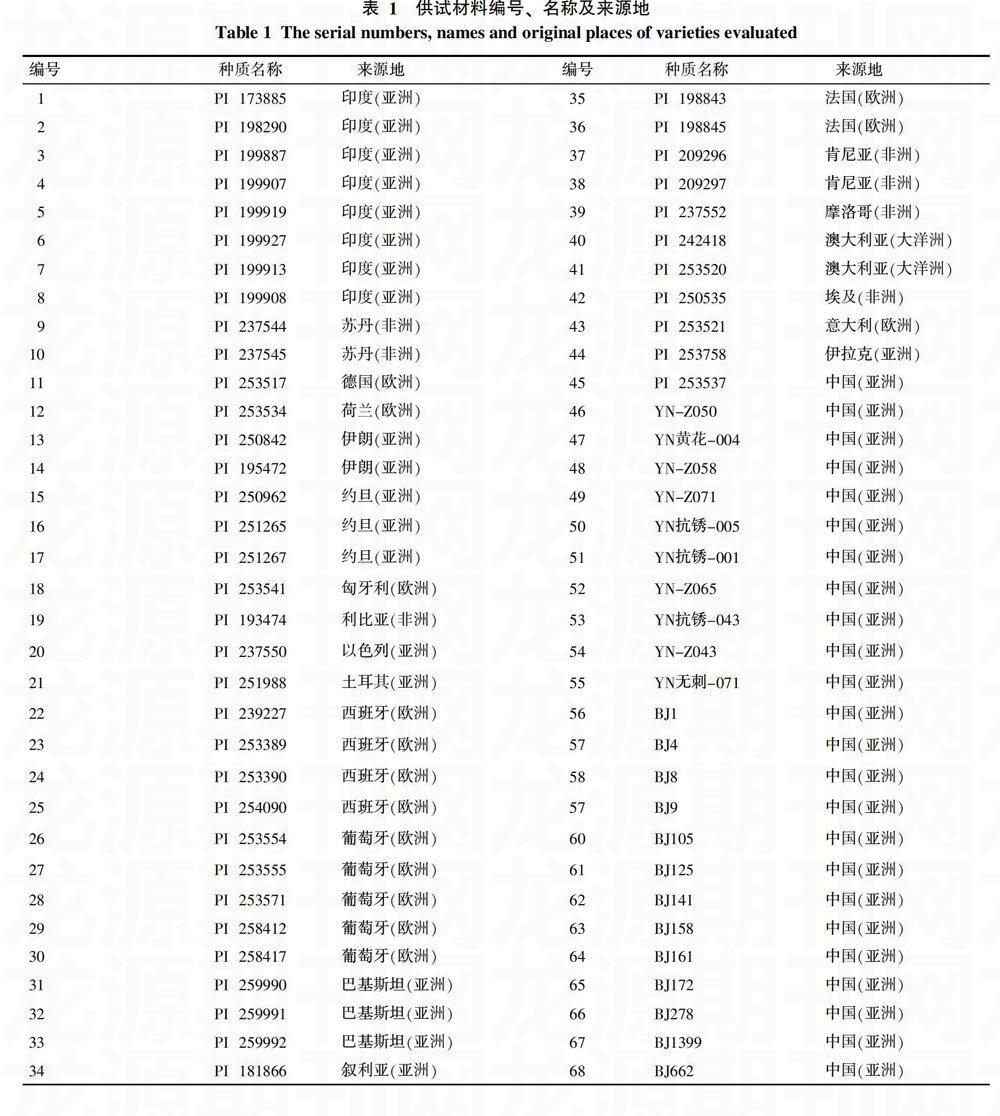
从已收集保存48个国家的3 000余份红花种质资源中,结合多年田间试验筛选出的68份优异种质为材料,研究其形态性状遗传多样性。供试材料编号、名称及来源地见表1。
1.2 方法
1.2.1 试验设计 试验于2008~2009年在云南省元谋县实验农场进行,试验点位于海拔1 118.4 m,E101°52′,N25°44′;年均气温21.9 ℃≥10 ℃,年积温7 786 ℃,终年基本无霜(持续时间350 d),年降雨量615.1 mm,雨量占年降水量的90%以上;该区光热资源丰富,年日照时数2 670 h,日照百分率62%,全年太阳总辐射量641.8 kJ/㎝2[11-12]。土壤类型为沙壤土,肥力中等,有灌溉条件。试验采用随机区组设计,3次重复;每个小区种植4行,行长1.5 m,行距40 cm,株距15 cm,小区面积2.4 m2。其它田间管理措施同大田一致。
1.2.2 性状调查 性状的观察与记录用《红花种质资源描述规范和数据标准》中的统一标准进行[13]。田间目测观察叶缘、叶形、叶刺、花色等形态性状;成熟时每个小区随机取样10株室内考种,调查株高、茎粗、第一分枝高度、最末分枝高度、分枝长度、分枝总数、单株有效果球数、顶果球直径、果球着粒数、千粒重、单株产量、小区产量、种子形状、种子壳型等。统计分析的性状分为两类:一类是质量性状,包括叶缘、叶形、叶刺、花色、种子形状、种子壳型6个性状;另一类是数量性状,包括生育期、株高、茎粗、第一分枝高度、最末分枝高度、分枝长度、分枝总数、单株有效果球数、顶果球直径、果球着粒数、千粒重、单株产量、小区产量等13个性状。计算平均值、标准差、最小值、最大值、变异幅度、变异系数和遗传多样性指数。
1.3 数据处理及统计分析
利用Excel2007计算供试材料13个数量性状的总体平均数(X)、标准差(s)和变异系数(CV%)。根据总体平均数(X)和标准差(s)计算结果将所有材料划分为10级,按第1级[Xi<(X-2s)]到第10级[Xi>(X+2s)], 每0.5 s为1级,Xi为第i级中的数据,每1级的相对频率(Pi)用于计算多样性指数。遗传多样性指数即Shannon-Weiner index(H′)的计算公式[14-15]如下:H′=PilnPi,式中n为某一性状表型级别的数目,Pi为某性状第i级别内材料份数占总份数的百分比。采用DPS7.0软件进行主要农艺性状的主成分分析和聚类分析。聚类分析时将原始数据进行标准化处理,采用欧氏距离和离差平方和法进行数据分析[16-17]。
2 结果与分析
2.1 红花种质资源质量性状遗传多样性分析
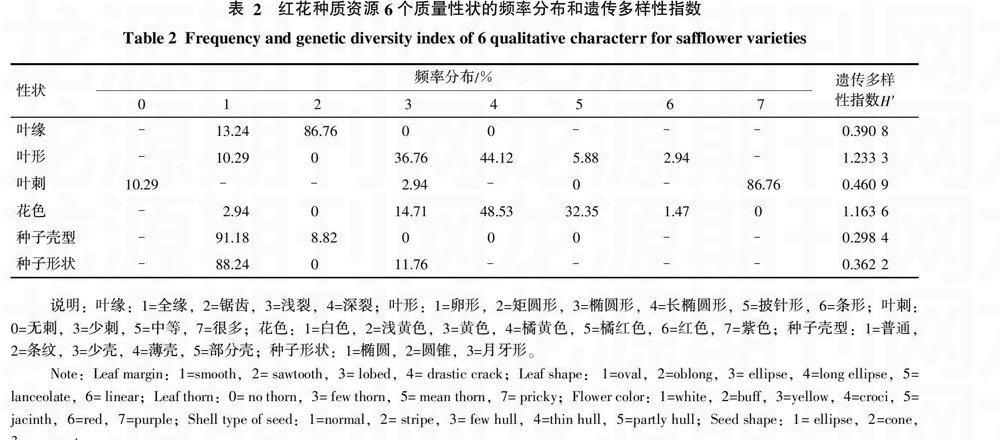
对68份红花种质资源的6个质量性状进行统计分析,结果(表2)显示,红花种质资源叶缘多为锯齿(86.76%),叶形多为长椭圆形(44.12%),叶刺(86.76%)为很多,花色则多为橘黄色和橘红色(分别为48.53%和32.35%),种子壳型多为普通(91.18%), 种子形状(88.24%)为椭圆。遗传多样性分析可知,叶形性状表现出较丰富的遗传多样性,遗传多样性指数最高为1.233 3,其次是花色遗传多样性指数为1.163 6;叶形在5种变异类型中以长椭圆形和椭圆形为主,卵形次之,少数为披针形和条形。花色在5种变异类型中以橘黄色和橘红色为主,黄色次之,少数为白色和红色。
2.2 红花种质资源数量性状遗传多样性分析
对13个数量性状进行统计分析,结果表明(表3),红花种质资源数量性状差异明显,变异范围大,遗传多样性丰富。13个性状的平均变异系数为25.41%。生育期、株高、茎粗、最未分枝高度、顶果球着粒数、顶果球直径、千粒重的变异系数小于20%;分枝总数、单株有效果球数的变异系数大于50%;其他4个性状的变异系数在20%~50%之间。说明参试材料的生育期、株高、茎粗、最未分枝高度、顶果球着粒数、顶果球直径、千粒重等一致性较强,变异范围不大;相比之下,分枝总数、单株有效果球数、单株产量、第一分枝高度、小区产量、分枝长度等性状的变异范围更大。
13个性状的平均遗传多样性指数为10.024 9,其中,分枝总数的遗传多样性指数最高为15.779 2,生育期最低为2.656 0两者相差13.123 2;分枝总数、单株有效果球数、单株产量、分枝长度、茎粗、顶果球直径的遗传多样性指数比其他7个性状高,说明其遗传多样性更丰富。
2.3 主成分分析
对红花13个主要数量性状采用DPS 7.0软件进行主成分分析,结果表明,前5个主成分累计贡献率达85.08%。在所有主成分构成中,包含了13个性状的绝大部分信息(表4),主成分一贡献率最大,为40.90%,其次为主成分二、三、四、五,贡献率依次为21.62%、9.50%、7.63%、5.44%。
由表4可知,第一主成分特征值为5.32,贡献率为40.90%。在第一主成分的特征向量中,载荷较高且符号为正的性状以单株有效果球数的值最大为0.412 4,其后依次是分枝总数、单株产量、小区产量、茎粗、分枝长度和顶果球着粒数,其特征向量值依次为0.412 3、0.377 5、0.321 7、0.318 0、0.281 2和0.262 2。此类性状为产量构成因子;符号为负的性状有第一分枝高度、最未分枝高度、千粒重、生育期和顶果球直径,以第一分枝高度的值最大为-0.325 3、其后依次为-0.150 5、-0.146 1、-0.122 3和-0.002 6,由各载荷数值可看出,在一定范围内,随着单株有效果球数的增加产量会有所提高,但超出一定范围内,单株有效果球数过大,第一分枝高度过高就会直接影响千粒重,即影响产量性状的最佳发挥。因此,在高产育种工作中,应注意单株有效果球数、第一分枝高度、千粒重、生育期的选择,第一主成分的相关值不能过高。
第二主成分特征值为2.81,贡献率为21.62%。第二主成分的特征向量中,所有性状的载荷数值符号均为正,对应特征向量中以株高的值最大,其次是最未分枝高度、第一分枝高度、生育期、顶果球直径、分枝长度、茎粗,此类性状主要与植株的生长势有关,可称为生长势因子。其载荷值较高而且均为正,说明株高越高,生育期越长,最未分枝高度和第一分枝高度越高,生长势越强;其余6个性状载荷数值较小,表明在育种工作中,应注意株高、最未分枝高度、第一分枝高度、生育期的选择,否则株高、最未分枝高度、第一分枝高度过高,导致分枝总数少,单株有效果球数少,顶果球着粒数较少,生育期晚直接影响产量,因此,在品种选择上应以生育期适中、生长势适中的为好。
第三主成分特征值为1.24,贡献率为9.50%。从载荷数值看千粒重的值最大,其后依次是生育期、分枝长度和顶果球着粒数,这些性状主要反映了红花品种的子粒大小特点,顶果球直径的载荷值较高为-0.587 6,其与千粒重表现为负相关;由各载荷数值可看出,千粒重过高是影响产量的重要因素,如追求过高的千粒重,会直接导致顶果球直径、单株有效果球数、单株产量、小区产量的降低,不利于产量的提高。因此,在育种过程中,应根据选育目标有针对性的进行性状选择。
第四主成分特征值为0.99,贡献率为7.63%。由各载荷数值可看出,第四主成分主要反映顶果球着粒数,从载荷数值的大小可知,顶果球着粒数增加,但是子粒小,千粒重明显下降,另外,由于株高过矮,导致分枝总数较少、单株有效果球数减少、单株产量降低,从而影响产量。所以,在育种过程中应根据选育目标,注意选择植株高度适中的品种。
第五主成分特征值为0.71,贡献率为5.44%。对应的特征向量中以生育期的值最大为-0.663 1,由各载荷数值可看出,生育期较晚,会直接导致分枝总数减少、单株有效果球数减少、果球着粒数减少,直接影响产量。因此,在实际工作中,应注意选择生育期适中的品种才能提高产量。
2.4 聚类分析
采用DPS 7.0软件,根据13个性状在68份红花种质资源间的不同表现,以欧氏距离为遗传距离,聚类分析采用离差平方和法,在遗传距离13.29处将供试材料聚为7大类群(图1),各品种类群的均值列于表5。
分析结果表明,第Ⅱ类群包含18份材料,这一类材料特征是叶缘均为锯齿,叶形以长椭圆形居多,卵形和椭圆形次之,叶刺全部为很多,花色以橘黄色居多,橘红色次之,种子壳型为普通,种子形状椭圆形,株高适中,千粒重平均值较高,在同一性状的7个种质群中处于第1位,此类群材料可依据育种目标作为大粒杂交亲本加以利用;第Ⅵ类群包含9份材料,表现叶缘均为锯齿,叶刺为很多,叶形以长椭圆形居多,少数为披针形,花色以橘黄色居多,少数为黄色,种子壳型以条纹壳较多,普通壳次之。种子形状全部为椭圆形,生育期适中、分枝总数多、单株有效果球数多、顶果球着粒数多、单株产量高、小区产量高等特点,可选择这类材料作为杂交亲本,以培育高产品种;第Ⅶ类群包含2份材料为62和66;这两份材料表现叶刺为很多,花色均为橘黄色,叶缘均为锯齿,叶形以长椭圆形、卵形各占一半,种子壳型为普通,种子形状椭圆形;生育期适中、株高适中、分枝长度、分枝总数、单株有效果球数、顶果球着粒数、单株产量和小区产量平均值在7个类群中为最高,这两份材料可作高产育种种质利用。
聚类结果表明,不同地理来源的种质资源有比较宽的遗传基础,聚类分析中部分地理来源相同或遗传背景相似的资源能够聚在同一类群,但也有一些地理来源及遗传背景不一致的材料也聚在同一类群。表明不同地理来源的红花种质资源在外部性状特征间具有丰富的多样性[18]。如第Ⅴ类群7份品种资源中有2份来自印度,2份来自葡萄牙,1份来自匈牙利,1份来自巴基斯坦,1份来自中国,表明其亲缘关系复杂,证实了不同地理来源品种的丰富多样性,为红花品种改良及资源的进一步开发利用提供参考依据。
3 讨论与结论
遗传多样性是生物多样性的核心和重要组成部分,任何一个物种都具有其独特的基因库和遗传组织形式,物种的多样性也就是显示了基因遗传的多样性[19]。种质资源的遗传多样性是生物进化和育种工作的基础,通过形态性状的遗传多样性研究,从整体上了解红花资源的丰富程度,为优良基因资源的发现和利用提供重要信息[20]。在红花育种中实践证明,新品种选育有赖于优良基因的发现和利用,因此,红花品种改良的关键是种质资源的有效利用[21]。
本文通过对以上19个性状的遗传多样性分析表明,红花种质资源存在广泛的性状变异,性状变异的表现是遗传因素与环境因素共同作用的结果,在栽培种植条件一致和材料数较多的情况下,可真实地表现资源间的遗传多样性。分析结果表明多样性指数越高遗传多样性越丰富,而丰富的多样性为作物育种和遗传改良奠定了基础。主成分分析可以详细地描述不同物种及其样本之间存在差异的主要原因。利用主成分分析法,前5个主成分因子反映了13个数量性状的全部信息,累计贡献率达85.08%,各主成分包含的信息具有一定的相关性,而不同性状在5个主成分中具有明显不同的载荷值,表明主成分分析结果与参试资源和性状指标的选择均有关系。通过以上分析结果表明,产生差异的原因可能由于所选择的群体和研究的性状不同。这与李艳花等[22]研究结果相似,在杂交育种和亲本选配时,可根据育种目标选择主成分互补的材料配制组合。
聚类分析方法是研究不同物种及其样本之间相似性的有效方法,不仅可以评价不同物种及其样本之间的性状类型分类,还可以研究不同种质材料的遗传差异[23]。本研究采用DPS 7.0软件,根据13个性状在红花种质资源间的不同表现,在遗传距离13.29处将68份供试材料聚为7大类群,同一地区的大部分品种聚在同一类群中。这与先前研究地方红花种质资源的形态标记聚类分析[24]比较有异同,同的是所选性状可正确有效地区分红花种质资源差异,了解种质资源间性状的相似性状况,有利于更准确地选择特异资源材料。异的是根据红花种质资源各性状形态指标的编码结果进行分析,并根据形态标记的相似性对红花品种资源进行系统聚类分析,能在一定程度上反映种质资源之间亲缘关系的远近。研究结果表明,供试资源品种的遗传多样性较丰富,说明品种类群间的遗传分化与地理分布有一定的联系又不绝对相关。这与贾晓艳等[25]研究结果一致,说明地理因素对品种有很大影响。但是,也有部分来自不同国家或地区的资源品种表现相似而聚为一类,这有可能是由于不同来源,不同地理环境、海拔高度的品种资源种植在同一环境条件下有些品种性状上可能出现具有相似的综合生态适应反应;或者尽管地理上相距较遥远,但是资源材料产地的生态条件相似。这与崔艳华等[26]研究结果相似,即不同地区的材料聚类呈现一定的地理分布规律。同时少部分来自不同地方的材料因生态条件相似而聚在了一类。根据育种学理论[27],不同类群间组配杂交组合有可能产生可供育种利用的遗传变异,故选配亲本时应以各类群间性状差异进行选择,而不能仅靠地理来源选择。
本研究针对红花资源主要农艺性状进行了系统的分析评价,分析结果表明红花品种间遗传差异较大,生物多样性较丰富。实践证明,在杂交或其他方法育种中,选配亲本材料可以依据主成分的排序,从红花种质资源中发掘有用基因,具体分析与全面评价每个亲本材料综合指标的优劣,依据育种目标,结合聚类分析,合理地选配组合,为促进红花品种的遗传改良或选育出高产、优质的新品种提供可靠依据。
参考文献
[1] 王兆木. 世界红花种质资源评价与利用[M]. 北京: 中国科学技术出版社, 1993: 26-58.
[2] 谭美莲, 严明芳, 汪 磊,等. 世界特种油料种质资源保存概况[J]. 植物遗传资源学报, 2011, 12(3): 339-345.
[3] 黎大爵. 世界红花种质资源的生物多样性及可持续利用[C]. 第三届生物多样性保护与利用高新科学技术国际研讨会论文集,2003: 22-24.
[4] 吴应祥, 黎大爵. 红花[M]. 农业出版社, 1982: 146.
[5] 张宗文. 红花遗传资源的研究与利用[J]. 植物遗传资源科学, 2000, 1(1): 7-10.
[6] 裴鑫德. 多元统计分析及其应用[M]. 北京: 北京农业大学出版社, 1990: 213-235.
[7] 贺晨帮,宗绪晓. 豌豆种质资源形态标记遗传多样性分析[J]. 植物遗传资源学报, 2011, 12(1): 42-48.
[8] 段利云, 王通强, 阳标仁,等. 甘蓝型油菜主要农艺性状的主成分和聚类分析[J]. 山地农业生物学报, 2007, 26(5): 381-385.
[9] 胡兴雨, 陆 平, 贺建波,等. 黍稷农艺性状的主成分分析与聚类分析[J]. 植物遗传资源学报, 2008, 9(4): 492-496.
[10] 徐东旭, 姜翠棉, 宗绪晓,等. 蚕豆种质资源形态标记遗传多样性分析[J]. 植物遗传资源学报, 2010, 11(4): 399-406.
[11] 张建平. 元谋干热河谷区蒸发量减少原因的灰色关联分析[J]. 云南地理环境研究, 1994, 6(2): 68-74.
[12] 何毓荣, 徐建忠, 黄成敏,等. 金沙江干热河谷区变性土的特征及系统分类[J]. 土壤学报, 1995, 32(增刊): 102-103.
[13] 杨建国, 刘旭云, 严兴初, 等.红花种质资源描述规范和数据标准[M]. 北京: 中国农业出版社, 2007.
[14] 白新桂. 数据分析与实验优化设计[M]. 北京: 清华大学出版社, 1986.
[15] 蔡一林, 刘志斋, 王天宇,等. 国内部分玉米地方品种的品质与农艺性状的表型多样性分析[J]. 植物遗传资源学报, 2011, 12(1): 31-36.
[16] 段会军, 张彩英, 王省芬,等. 河北省大豆品种主要农艺性状及聚类分析[J]. 河北农业大学学报, 2003, 26(2): 5-9.
[17] 王瑞珍, 赵朝森, 程春明,等. 我国中部湖南湖北两省野生大豆种群表型多样性分析[J]. 江西农业学报, 2009, 21(12): 1-4.
[18] 廖 丽, 白昌军, 郭晓磊,等. 竹节草种质资源形态多样性研究[J]. 热带作物学报, 2011, 32(11): 2 042-2 047.
[19] 葛 颂. 生物遗传多样性研究的原理及方法[M]. 北京:科学出版社, 1994.
[20] 郭丽芬, 张 跃, 徐宁生,等. 红花种质资源主要数量性状的灰色关联度分析[J]. 西南农业学报, 2013, 26(1): 328-333.
[21] 郭丽芬, 徐宁生, 张 跃,等. 云南红花种质资源主要农艺性状的遗传多样性分析[J]. 植物遗传资源学报, 2012, 13(2): 219-225.
[22] 李艳花, 杜成章, 陈 红,等. 重庆大豆地方资源多样性评价及群体表型特点研究[J]. 植物遗传资源学报, 2013, 14(6): 44-49.
[23] 游明安. 长江下游大豆地方品种的聚类分析[J]. 中国油料,1994, 16(4): 36-45.
[24] 郭丽芬, 张 跃, 胡尊红,等. 地方红花种质资源的形态标记聚类分析[J]. 河南农业科学, 2013, 42(2): 41-45, 97.
[25] 贾晓艳, 张彩英. 河北省大豆推广品种遗传多样性分析[J]. 植物遗传资源学报, 2006, 7(3): 310-315.
[26] 崔艳华, 邱丽娟, 常汝镇, 等. 黄淮夏大豆遗传多样性分析[J]. 中国农业科学, 2004, 37(1): 15-22.
[27] 杨春燕, 闫 龙,张孟臣. 河北省大豆地方品种遗传基础[J]. 植物遗传资源学报, 2009, 10(4): 560-565.
猜你喜欢
长江蔬菜·学术版(2016年12期)2017-01-12
农家科技下旬刊(2016年9期)2016-12-15
大学教育(2016年11期)2016-11-16
大经贸(2016年9期)2016-11-16
中小企业管理与科技·上旬刊(2016年10期)2016-11-15
考试周刊(2016年84期)2016-11-11
中国市场(2016年33期)2016-10-18
科技视界(2016年20期)2016-09-29
企业导报(2016年9期)2016-05-26
