
云南河口不同林龄人工橡胶林土壤CO2浓度的变化规律及其影响因素
2015-04-29 00:44李涛李芹王树明李春赵东兴张勇高梅
热带作物学报 2015年1期
李涛 李芹 王树明 李春 赵东兴 张勇 高梅

摘 要 以云南河口不同林龄人工橡胶林作为研究对象,对抗逆高产和胶木兼优橡胶林土壤CO2浓度及相关影响因素进行2 a的动态观测。结果表明:(1)从季节变化来看,土壤CO2浓度具有明显的季节性变化特征,每年春季开始上升,夏秋季最高,冬季最低;(2)从垂直土壤深度来看,土壤CO2浓度随着土壤深度的递增而升高,在0~100 cm土壤范围内,表现为100 cm>80 cm>60 cm>40 cm>20 cm;(3)土壤CO2浓度与温度、水分和土壤有机质之间存在着显著或非显著正相关关系,表明温度、水分、有机质等是影响土壤CO2浓度变化的主要因素。
关键词 橡胶林;土壤CO2浓度;温度;含水量;有机质
中图分类号 S718.5 文献标识码 A
The Variation and Influence Factors of Soil CO2
Concentration in Planted Hevea brasiliensis
with Different Ages in Hekou,Yunnan
LI Tao,LI Qin*,WANG Shuming,LI Chun
ZHAO Dongxing,ZHNAG Yong,GAO Mei
Honghe Institute of Tropical Agriculture,Hekou,Yunnan 661300,China
Abstract Planted Hevea brasiliensis with different ages with high latex yield and stress tolerance, and also with quality timber in Hekou,Yunnan was used to monitor soil CO2 concentration and related influencing factors for two years. The results showed:(1)The soil CO2 concentration changed obviously with season passed. It rose from spring, reached the highest in summer and autumn, and declined to the lowest in winter;(2)In the vertical soil profile, the soil CO2 concentration of the two studied sites rose with soil depth increased, and showed as 100 cm >80 cm>60 cm>40 cm>20 cm within 0-100 cm limits;(3)There was a positive correlation or a nonsignificant correlation between the soil CO2 concentration with temperature,moisture content and soil organic matter. The major factors which influenced the soil CO2 concentration were the temperature, moisture content, soil organic matter and so on.
Key words Rubber plantation; Soil CO2 concentration; Temperature; Moisture content; Organic matter
doi 10.3969/j.issn.1000-2561.2015.01.002
土壤系统中的CO2主要来源于植物根系呼吸及土壤有机质的氧化分解[1-2],土壤CO2一部分参与到土壤化学反应或者扩散到土壤水中[3],另一部分经由土壤呼吸作用排放到大气中,已有研究结果表明,土壤呼吸是土壤碳库和大气碳库之间碳流通的主要方式之一[4],据估计全球土壤CO2排放量在68~100 pg/a之间[5-6],是整个生物圈的主要碳源之一。碳循环在全球生态系统中至关重要,许多发达国家及政府部门都加大了对人工林碳吸收、碳储存途径的研究力度。中国大面积种植巴西橡胶树已有50多年的历史,目前天然橡胶种植面积已达114万hm2,年产量达86.5多万t,种植面积和产胶量均居世界43个产胶国家的第四位,已经建立起较为完善的天然橡胶产业体系。橡胶林也是南方热作垦区重要的人工生态林之一[7],具有森林型生态系统的特点,进行着大量物质、能量循环和转移,是实现热区资源可持续利用的一个重要人工生态系统[8]。橡胶树除生产橡胶和提供优质木材外,还有长期不为人们注意的生态服务功能,如橡胶树可固定大气中的C和缓解温室效应,橡胶林一生的生命周期里可固定大气中的C为318.7 t/hm2[9],中国橡胶林每年从大气中固定的CO2量为6.51 t/h,采用C税法估算其价值达每年19.6万元/hm2[10]。
20世纪80年代以后对胶园生态系统的研究主要集中在养分及养分循环、胶园小气候等方面,而对大气调节作用方面的研究报道较少。有学者对西双版纳橡胶林土壤呼吸季节变化及其影响因子进行了研究,显示该地区土壤呼吸速率季节变化与橡胶树生长节律基本一致,且与温度因子、湿度因子有相关关系[11],土壤温湿度是影响橡胶林土壤呼吸作用形成季节变化的主要原因之一[12]。但中国作为世界上少有的几个大面积人工种植橡胶的国家[13],在这方面的研究远远不够,尤其对橡胶林土壤呼吸作用的研究报道极少。因此,笔者研究橡胶林土壤CO2浓度的季节变化规律及其分布特征,对不同橡胶林、不同深度土壤CO2浓度进行长期动态观测,以此更好地了解天然人工橡胶林碳循环途径及其变化规律,为研究人工橡胶林对全球碳循环的影响提供基础数据,也为中国橡胶产业的固碳减排工作提供一定的科学理论依据。
1 材料与方法
1.1 研究区概况
试验选择云南河口红河综合试验站抗逆高产和胶木兼优2个不同林龄人工橡胶林作为研究对象(图1)。其中,胶木兼优橡胶林于2003年4月定植,主要品种为178、509、516、524、8-79、628等12个,种植面积约4.73 hm2;抗逆高产橡胶林于2010年6月定植,主要品种为云研77-4、热研7-33-97、热研8-79、热垦525、热垦628和湛试327-13共6个,种植面积约12.07 hm2。试验区属热带雨林季风性气候,最高温40.9 ℃,最低温1.9 ℃,年均温22 ℃,年均降水1 587.3 mm,年均湿度84%,日照1 605 h;土壤以灰褐色粘土为主,土壤分布均匀,土层较厚,土壤基本理化性状平均值为:pH4.24、有机质30.75 g/kg、有效磷8.24 mg/kg、速效钾49.80 mg/kg、碱解氮94.65 mg/kg、交换钙100.1 mg/kg和交换镁13.3 mg/kg。
1.2 方法
每个研究点设置不同土壤深度即20、40、60、80、100 cm,共计5个不同深度土层,监测每个点每个不同土层的CO2浓度。具体在2个橡胶林下分别挖掘1m深的土坑,垂直土壤剖面上按不同深度要求凿洞,把PVC管制作的“L”型土壤CO2收集管埋放于内,水平管为土壤气体收集管,其上钻有通气孔与土壤气体交换;竖管为导气管,导气管延伸至地面,导管开口上插有塞子防止土壤气体与大气交换(图2)。使用加拿大BW公司生产的GasAlertMicro 5 IR CO2检测仪直接测定,每个土层深度的CO2浓度重复测定3次,取平均值作分析统计,观测周期为每月2次,持续2 a。
1.3 测定项目
橡胶林内空气的温度、湿度用德国Testo 435-2多功能测量仪直接测定;土壤有机质采用重铬酸钾氧化-外加热法[14];橡胶林下土壤温度使用水银温度计插入土中测定;土壤含水量采用烘干法,即105 ℃烘干8 h后称重测定,计算公式为:土壤含水量=[(湿土重量-烘干后重量)/湿土重量]×100%。
2 结果与分析
2.1 土壤CO2浓度
2.1.1 土壤CO2浓度的季节变化规律 从图3、4可知,2个试验点的土壤CO2浓度均具有明显的季节性变化特征,从3月份开始随着春季的到来,气温回升,光照增强,降雨量增多,土壤呼吸作用随之增强,土壤CO2浓度开始缓慢上升,并在夏季(6~8月份)达到最大值,秋季开始下降,冬季最低。其中夏季最大值达到48 500 mg/kg,冬季最小值为4 500 mg/kg,夏季土壤CO2的浓度是其它季节的2~3倍,说明橡胶林土壤CO2的浓度具有明显的季节性变化规律。由图5可知,从每年的4月份开始,一方面随着雨季的到来,气温回升,降雨量增多,橡胶树及地表植被光合作用增强,土壤呼吸作用也随之增强,新陈代谢活动旺盛,根系呼吸作用释放出的CO2增多;另一方面夏季时在高温高湿的气候条件下,土壤微生物活动能力最强,土壤中有机物的氧化分解速度变快,释放出的CO2更多,使得橡胶林土壤CO2浓度在夏季显著上升。
2.1.2 土壤CO2浓度的垂向变化特征 从垂直土壤深度看,随着土壤深度递增,2个试验点的土壤CO2浓度均出现升高现象(图3、4),表现为深层土土壤CO2浓度大于表层土土壤CO2浓度,在0~100 cm土壤范围内,土层越深,其CO2浓度越高。在夏秋季节变化更明显,且各个不同深度土层的CO2浓度变化梯度增大,20、40和60 cm这3个相邻土层的变化幅度最大,变化范围相差5 000~10 000 mg/kg,但近底部80和100 cm这2个相邻土层的变化幅度一年四季中均变化较小,平均变化范围相差1 000~3 000 mg/kg,总体上趋于稳定;而冬春季时,各个相邻土层的CO2浓度变化梯度不明显,相差仅2 000~3 000 mg/kg。
2.2 土壤CO2浓度与气温、土温、土壤含水量和土壤有机质的关系
2.2.1 土壤CO2浓度与气温的关系 土壤微生物呼吸和植物根系呼吸是构成土壤呼吸的2个主要组分[15-16],土壤系统中68%的CO2由土壤微生物呼吸释放,气温的变化是影响土壤微生物活动性的主要因素之一[17]。微生物活动能力随温度的升高而加强,并加速了土壤中有机物质的氧化分解,产生更多CO2。
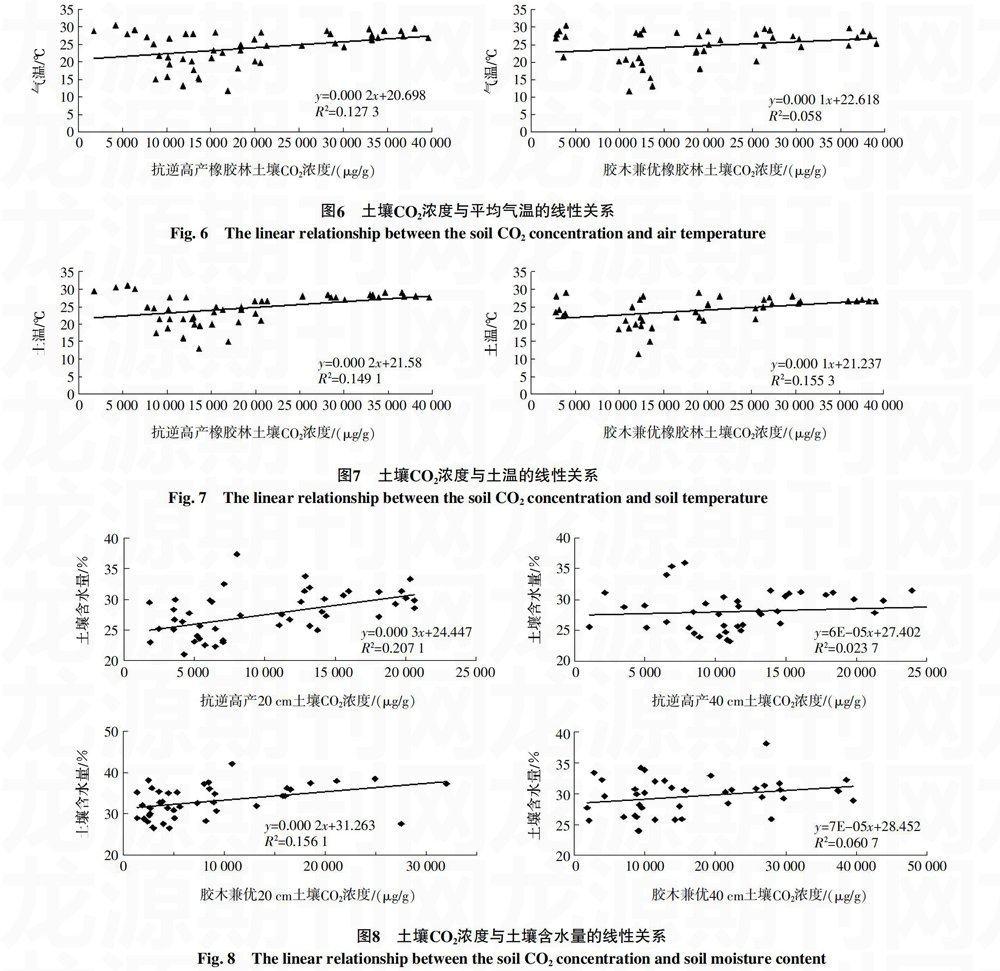
利用Excel、SPSS软件对橡胶林土壤CO2浓度的平均值与气温作相关性分析及显著性检验(图6),得知抗逆高产橡胶林土壤CO2浓度与气温的相关系数R=0.357*,P=0.013;胶木兼优橡胶林土壤CO2浓度与气温的相关系数R=0.241,P=0.120。说明温度在0~35 ℃范围内,抗逆高产橡胶林土壤CO2浓度与气温之间存在显著正相关关系(p<0.05),胶木兼优橡胶林土壤CO2浓度与气温之间存在非显著正相关关系(p>0.05)。
2.2.2 土壤CO2浓度与土温的关系 橡胶林土壤CO2浓度的平均值与土温作相关性分析及显著性检验可知(图7),抗逆高产橡胶林土壤CO2浓度与土温的相关系数R=0.386**,P=0.007;胶木兼优橡胶林土壤CO2浓度与土温的相关系数R=0.394**,P=0.009。说明抗逆高产和胶木兼优橡胶林土壤CO2浓度与土温之间均存在极显著正相关关系(p<0.01)。土温变化缓慢,外界环境对土壤温度的变化影响不大,土温比气温稳定一些,更能反映温度对CO2浓度的影响,已有研究结果表明,土壤温度能较准确地反映温度对土壤微生物的影响,是影响土壤呼吸的重要因素之一[18]。
2.2.3 土壤CO2浓度与土壤含水量的关系 橡胶林20和40 cm土壤CO2浓度与土壤含水量作相关性分析及显著性检验可知(图8),抗逆高产橡胶林土壤CO2浓度与含水量的相关系数分别为20 cm处R=0.455**、P=0.001,40 cm处R=0.153、P=0.301;胶木兼优橡胶林土壤CO2浓度与含水量的相关系数分别为20 cm处R=0.395**、P=0.010,40 cm处R=0.247、P=0.115。说明橡胶林20 cm深度土壤CO2浓度与土壤含水量之间存在极显著正相关关系,而40 cm深度的为非显著正相关关系,由此表明橡胶林土壤CO2浓度与土壤含水量之间的相关性随着土壤深度增加而减弱,仅在0~20 cm深度范围内存在极显著正相关关系,更深土层为非显著正相关关系。
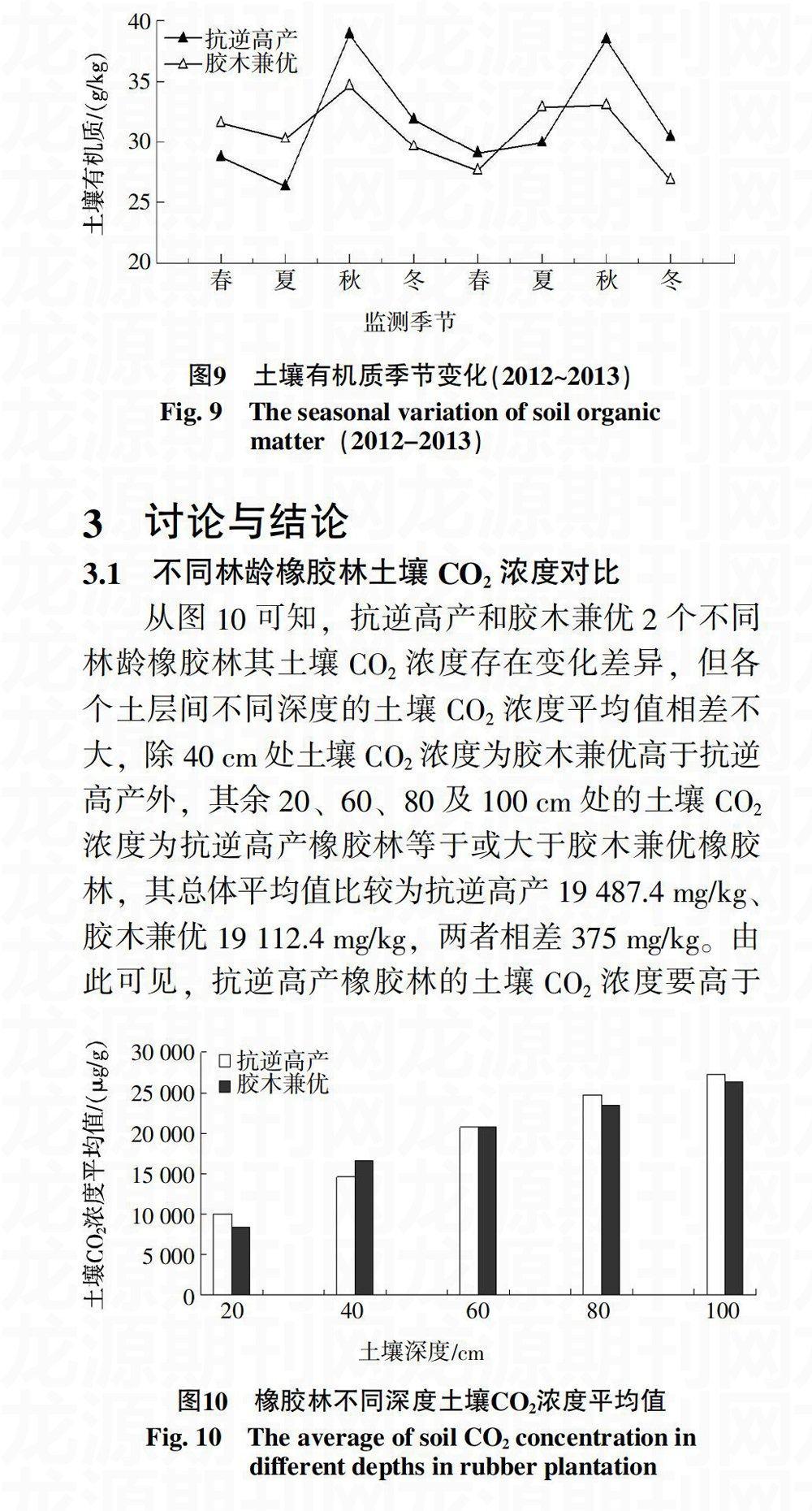
2.2.4 土壤CO2浓度与土壤有机质的关系 从图9可知,橡胶林土壤有机质含量呈季节性变化规律,抗逆高产橡胶林土壤有机质含量平均值为31.7 g/kg,胶木兼优橡胶林土壤有机质含量平均值为30.8 g/kg,说明该区域内土壤有机质含量较高,从数值上比较抗逆高产橡胶林土壤有机质含量略高于胶木兼优橡胶林,但两者相差不大。利用SPSS软件对各个季节土壤CO2浓度和有机质含量的平均值作相关性分析及显著性检验,得知抗逆高产橡胶林的相关系数R=0.554,P=0.154;胶木兼优橡胶林的相关系数R=0.274,P=0.511。说明橡胶林土壤CO2浓度与土壤有机质之间均存在非显著正相关关系。
3 讨论与结论
3.1 不同林龄橡胶林土壤CO2浓度对比
从图10可知,抗逆高产和胶木兼优2个不同林龄橡胶林其土壤CO2浓度存在变化差异,但各个土层间不同深度的土壤CO2浓度平均值相差不大,除40 cm处土壤CO2浓度为胶木兼优高于抗逆高产外,其余20、60、80及100 cm处的土壤CO2浓度为抗逆高产橡胶林等于或大于胶木兼优橡胶林,其总体平均值比较为抗逆高产19 487.4 mg/kg、胶木兼优19 112.4 mg/kg,两者相差375 mg/kg。由此可见,抗逆高产橡胶林的土壤CO2浓度要高于胶木兼优橡胶林,抗逆高产橡胶林于2010年6月定植,树龄3 a,而胶木兼优橡胶林于2003年4月定植,树龄10 a,由于两者所处的地理位置、土壤环境及气候类型大致相同,在不考虑橡胶品系的前提下,可进一步说明在不同林龄人工橡胶林中,幼林地土壤CO2浓度平均值要比开割林地高。
造成这一差异的原因可能是抗逆高产橡胶林为幼林地,其土壤系统经过几年时间的闲置修整,林带空间开阔,光照、雨水充足,林下生物多样性丰富,地上杂草丛生,植被种类多,土壤微生物活动强烈,使得其土壤中的营养元素、矿物质得到了恢复和补充,有利于土壤呼吸作用释放出更多的CO2;而开割林地阴蔽度较高,林下光照不足,其他植物生长受限,而且因为产胶量的需要,本地区每年4月份即开始割胶直至11月份结束,在近8个月的时间里每天进行割胶,橡胶林需不断合成胶乳,吸收了土壤中的大部分营养物质,使得土壤变得贫瘠,生物多样性下降,林下枯落相对较少,有机物氧化分解释放出的CO2少。
3.2 橡胶林不同深度土壤CO2浓度变化差异
在0~100 cm垂直土壤范围内,2个试验点的土壤CO2浓度均随着土壤深度的递增而升高,土层越深其CO2浓度越高,深层土壤CO2浓度大于表层土,表现为100 cm>80 cm>60 cm>40 cm>20 cm,而且不同深度变化幅度不同,各个土层的CO2浓度在冬春季变化幅度较小,梯度不明显;在夏秋季变化幅度较大,梯度最明显。原因可能是20、40和60 cm深度的土壤属于表层土,土壤疏松多孔,通透性良好,密度较低,土壤中的CO2容易向低浓度CO2的空气中扩散,而且受外界环境变化的影响较大,而80 cm和100 cm深度的土壤其深藏于地表之下,土壤通透性较差,密度大,土壤温度、含水量相对稳定,受外界环境干扰较小。土壤中的CO2是植物根系呼吸、微生物活动及大气扩散输入的混合[16],CO2的浓度受土壤的通气性、土壤生物化学过程的强度、气象条件及植被的共同影响[17]。此外,土壤CO2浓度还会受到土壤微生物活性的影响,而植物凋落物、光合产物向地下部分的分配及根系分泌物也会影响到土壤微生物的活性,从而影响土壤呼吸强度[19-21]。
3.3 橡胶林土壤CO2浓度影响因素
土壤呼吸作为一个复杂的生物学过程,其产生的CO2浓度受环境、气象及生物活性等多重因素的影响,通过对橡胶林土壤CO2浓度与气温、土温、土壤含水量以及土壤有机质之间作相关性分析及显著性检验,得知人工橡胶林土壤CO2浓度与气温、土温、土壤含水量以及土壤有机质之间均存在着正相关关系,有的表现为显著正相关,有的表现为非显著正相关。其中,与土温、20 cm深度土壤含水量之间存在极显著或显著正相关,与气温、40 cm深度土壤含水量之间存在非显著正相关。造成这种差异的原因可能是土壤CO2浓度同时受到气温、降雨、土壤含水量、土壤有机质、地表植被和生物活性等多个影响因子的共同控制,而不仅是某一个因子决定。就土壤含水量来说,监测数据显示20 cm深度土壤含水量波动幅度较大,变化呈季节性规律较为明显,所以与土壤CO2浓度显著相关;而40 cm深度土壤含水量比20 cm深度的土壤含水量要稳定一些,季节变化规律不明显并趋于平缓,所以与土壤CO2浓度非显著相关。通过分析表明温度、水分是影响土壤CO2浓度变化的主要因素,已有的研究报道也揭示了温度和水分是影响土壤呼吸最为显著的因素[22-25]。
综上表明,不同林龄人工橡胶林土壤CO2浓度的季节变化与温度(土温、气温)、土壤含水量和土壤有机质之间存在着显著或非显著正相关关系,说明橡胶林土壤CO2浓度的季节性变化规律与树龄、温度(土温、气温)、降雨量及生物活动密切相关。符合现有研究结论即橡胶林土壤呼吸与温度和土壤含水量都有明显的季节变化,而且变化趋势基本一致[26]。由此可见,温度、降水、凋落物和生物活性对CO2浓度变化都起着重要影响作用,气候条件、环境因子、水分条件等使地表植被生长状况及土壤微生物活动量随季节的变化而变化,进而影响土壤CO2浓度的变化,使土壤CO2浓度呈现季节性变化规律。
参考文献
[1] Witkamp M, Frank M. L. Evolution of CO2 from litter, humus and subsoil of a pine stand[J]. Pedobiologia. 1969, 9: 358-365.
[2] Fritz P, Reardon E J, Barker J. et al. The carbon isotope geochemistry of a small groundwater system in Northeastern Ontario[J]. Water Resources Research, 1978, 14: 1 059-1 067.
[3] Davidson E A, Belk E, Boone R D. Soil water content and temperature as independent or confounded factors controlling soil respiration in a temperature mixed hardwood forest[J]. Global Change Biology, 1998, 4: 217-227.
[4] Weber M G. Forest soil respiration after cutting and burning in immature aspen ecosystems[J]. For Ecol Manage, 1990, 31: 1-14.
[5] Hendry M J, Lawrence J R, Zanyk B N, et al. Microbial production of CO2 in the unsaturated dgeologic media in a mesoscale model[J]. Water Resour Res, 1993, 29: 973-984.
[6] Raich J W, Schlesinger W H. The global carbon dioxide fluxin soil respiration and its relation ship to vegetation and climate[J]. Tellus, 1992, 44: 81-89.
[7] 曹建华, 蒋菊生. 橡胶林生态系统养分循环研究进展[J]. 热带农业科学, 2007, 27(3): 48-56.
[8] Choudhury M, Sarma A C, Pal T K, et al. Available nutrient status of the rubber(Hevea brasiliensis)growing soils of Meghalaya, Mizoram, Manipur and southern parts of Assam[J]. Indian Journal of Hill Farming, 2001, 14(2): 76-80.
[9] Sivakumaran S, Yew F K. Carbon S equest ration in Rubber: Impli cation and Model to Fund Continued Cultivation[J]. IRRDB, Bogor-In dones ia, 2000, 9: 12-14.
[10] 蒋菊生, 王如松.橡胶林固定CO2和释放O2的服务功能及其价值估计[J]. 生态学报, 2002, 22(9): 1 545-1 551.
[11] 周文君, 沙丽清.西双版纳橡胶林土壤呼吸季节变化及其影响因子[J]. 山地学报, 2008, 26(3): 317-325.
[12] 卢华正, 沙丽清. 西双版纳热带季节雨林与橡胶林土壤呼吸的季节变化[J]. 应用生态学报, 2009, 20(10): 2 315-2 322.
[13] 吴志祥, 谢贵水, 陶忠良, 等. 不同树龄橡胶树土壤有机碳含量与储量特征[J]. 热带作物学报, 2009, 30(2): 135-141.
[14] 鲍士旦. 土壤农化分析(第3版)[M].北京: 中国农业出版社, 2000: 257-282.
[15] Singh B, Nordgren A, Lofvenius M O, et al. Tree root and soil heterotrophic respiration as revealed by girdling of boreal Scots pine forest: extending observations beyond the first year[J]. Plant, Cell and Environment, 2003, 26: 1 287-1 296.
[16] Fritz P M, Ozeto A A, Reardon E J. Practical consideration on carbon isotope studies on soil carbon dioxide[J]. Chemical Geology(Isotope Geoscience Section), 1985, 58: 89-95.
[17] 俞锦标, 李春华, 赵培道, 等. 贵州普定县岩溶地区土壤空气中CO2含量分布及溶蚀作用的研究[J]. 中国岩溶, 1985, 4(4): 325-331.
[18] 单建平, 陶大立. 国外对树木细根的研究动态[J]. 生态学杂志, 1992, 11(4): 46-49.
[19] Li Y, Xu M, Zou X, et al. Soil CO2 efflux and fungal and bacterial biomass in a plantation and a secondary forest in wet tropics in Puerto Rico[J]. Plant and Soil, 2004, 2689(1): 151-160.
[20] Boone R D, Nadelhoffer K J, Canary J D, et al. Roots exert a strong influence on the sensitivity of soil respiration[J]. Nature, 1998, 396: 570-572.
[21] Hogberg P, Nordgren A, Buchmann N, et al. Large-scale forest girdling shows that current photosynthesis drives soil respiration[J]. Nature, 2001, 411: 789-791.
[22] Schlentner R E, Van Cleve K. Relationships between CO2 evolution from soil, substrate temperature, and substrate moisture in four mature forest types in interior Alaska[J]. Can J For Res, 1984, 15: 97-106.
[23] Fang C, Moncrieff J B, Gholz H L, et al. Soil CO2 efflux and its spatial variation in a Florida slash pine plantation[J]. Plant and Soil, 1998, 205: 135-146.
[24] Goulden M L, Wofsy S C, Harden J W, et al. Sensitivity of boreal forest carbon balance to soil thaw[J]. Science, 1998, 279: 214-217.
[25] Ohashi M, Gyokusen K, Saito A. Measurement of carbon dioxide evolution from a Japanese cedar(Cryptomeria japonica D. Don)forest floor using an open-flow chamber method[J]. For Ecol Manage, 1999, 123: 105-114.
[26] 房秋兰, 沙丽清. 西双版纳热带季节雨林与橡胶林土壤呼吸[J]. 植物生态学报, 2006, 30(1): 97-103.
猜你喜欢
热带作物学报(2022年6期)2022-07-08
现代青年·精英版(2021年11期)2021-12-16
森林工程(2018年4期)2018-08-04
时代农机(2018年11期)2018-03-17
农民致富之友(2016年22期)2017-02-07
天津农业科学(2016年12期)2017-01-11
现代农业研究(2016年11期)2017-01-10
安徽农学通报(2016年20期)2016-12-26
浙江农业学报(2016年7期)2016-06-15
电源技术(2016年9期)2016-02-27
