
鸡IGF1R、IGFBP-3基因SNP位点之间互作效应分析
2015-03-15 12:32:06金崇富陈长宽杨智青陈应江
江苏农业学报 2015年5期
金崇富, 陈长宽, 杨智青, 时 凯, 陈应江
(江苏沿海地区农业科学研究所,江苏 盐城 224002)
胰岛素样生长因子(Insulin-like growth factor,IGF)是人体最重要的生长因子之一。IGF家族由2个多肽类生长因子(IGF1、IGF2)、2类受体(IGF1R、IGF2R)和7种结合蛋白(IGFBP1~IGFBP7)组成,它们是动物生长轴上重要的因子,具有强烈的促生长和分化作用[1]。2类IGF受体的结构和功能差异很大,其中第1类IGF受体(IGF1R)是IGF功能的主要介导者。Moe等[2]研究了IGF1R基因编码区单核苷酸多态性与日本鹌鹑体质量的相关性,发现不同基因型对10周龄体质量有极显著的影响。高凤华等[3]对东北农业大学肉鸡品系的IGF1R基因进行遗传多态性分析,发现不同基因型对5周龄体质量有显著影响。雷明明等[4]研究发现,C17337024G/T突变位点与生长和屠体性状显著相关,与初生质量显著相关。
IGFBP-3主要存在于血液中,它对IGF1和IGF2具有很高的亲和力,是血液中最为丰富的IGFBP结合形式。血液中95%的IGF1和IGF2都与IGFBP-3结合,因此,IGFBP-3对于IGF作用的发挥起着重要作用[5]。另外,IGFBP-3可以增强 IGF1的效应,能促进IGF1在骨骼上的合成代谢效应。胡沈荣等[6]对西农萨能奶山羊IGFBP-3基因进行了遗传多态性分析,发现西农萨能奶山羊IGFBP-3的Xsp-I基因座不同基因型间的第二胎产羔数有显著差异,且初生质量也存在显著差异(P<0.05)。刘哲等[7]对奶牛IGFBP-3基因进行了多态性分析,分析结果表明AA型12月龄体长和体高均极显著高于AB型。
数量性状是由许多微效基因或称多基因(Polygene)的联合效应控制的,数量性状直接影响畜牧产品的产量及经济效益。如果仅仅从单基因入手进行分子辅助育种,往往不能达到最终的育种目标。在分析多基因调控的数量性状时,需要同时对多个位点进行分析,这样才能得到基因与性状之间的真实相关性,在利用DNA标记进行辅助选择时才可能获得更大的遗传进展[8]。本研究以盐城地方草鸡为研究对象,对IGF1R、IGFBP-3基因进行互作效应分析,以期能较全面地研究盐城地方草鸡生产性状的分子遗传机制。
1 材料与方法
1.1 试验材料
从江苏沿海地区农业科学研究所生态养殖基地采集210只母鸡血样,同时记录母鸡生长性状(初生质量及4、8、12、16周龄体质量)和繁殖性状(开产日龄、开产体质量、开产蛋质量、300日龄产蛋数)。翅静脉采集血样1.5 ml,肝素钠抗凝,-20℃保存。DNA提取试剂盒(Omega blood DNA kit)法提取基因组DNA。
1.2 引物设计
根据GenBank中鸡IGF1R基因(NC_006097)和IGFBP-3基因(NC_006089)序列,设计IGF1R基因第2外显子引物IGR2-1和IGR2-2,以及IGFBP-3基因第1和2外显子引物IBP1和IBP2(表1)。引物由上海生物工程有限公司合成。

表1 引物信息Table 1 The information of the primers
1.3 PCR-SSCP 分析
将3μl PCR产物加入7μl变性缓冲液中,98℃变性10 min,然后迅速冰浴5 min。将变性PCR产物点样加入到10%聚丙烯酰胺凝胶中,140 V电压电泳12 h。在电泳结束后,进行银染显色并拍照记录结果。
1.4 IGF1R、IGFBP-3 基因综合标记
两基因位点间互作用以下模型进行方差分析:yijk=μ + Gi+ Gj+ Gij+ eijk,式中,yijk为性状测定值,μ 为群体平均值,Gi、Gj为基因型效应,Gij为位点i和位点j间的互作效应,e为随机残差效应。
2 结果与分析
2.1 IGF1R、IGFBP-3基因 SSCP引物的 PCR扩增
采用 PCR-SSCP法分析 IGR2-1、IGR2-2和IBP1、IBP2引物扩增片段。结果显示:IGR2-1扩增片段有3种带型,定义为AA、AB和BB;IGR2-2扩增片段有3种带型,定义为CC、CD和DD;IBP1扩增片段有3种带型,定义为EE、EF和FF;IBP2扩增片段有3种带型,定义为GG、GH和HH(图1)。
2.2 IGF1R、IGFBP-3基因序列分析结果
测序结果表明,引物IGR2-1的PCR产物片段的BB型与AA型相比在第26 333 bp处发生了G→A突变,引物IGR2-2的PCR产物片段的DD型与CC型相比在第26 336 bp处发生了G→A突变,引物IBP1的PCR产物片段的FF型与EE型相比在第160 bp处发生了T→G突变,引物IBP2-1的PCR产物片段的HH型与GG型相比在第1 087 bp处发生了C→T突变(图2)。

图1 引物IGR2-1(a)、IGR2-2(b)、IBP1(c)和IBP2(d)的SSCP分析结果Fig.1 SSCP analysis of PCR product using primers IGR2-1(a),IGR2-2(b),IBP1(c)and IBP2(d)

图2 鸡不同基因型PCR产物片段的序列比较Fig.2 Sequence alignment of PCR product segments of different genotypes in chickens
2.3 IGF1R、IGFBP-3基因群体遗传学分析
G26333A位点AA基因型为优势基因型,基因型频率为0.745;等位基因A为优势等位基因,等位基因频率为0.855。G26336A位点CC基因型为优势基因型,基因型频率为0.555;等位基因C为优势等位基因,等位基因频率为0.755。T160G位点FF基因型为优势基因型,基因型频率为0.495;等位基因F为优势等位基因,等位基因频率为0.703。C1087T位点GG基因型为优势基因型,基因型频率为0.415;等位基因G为优势等位基因,等位基因频率为0.615(表2)。
卡方适合性检验结果表明,群体除了在C1087T位点偏离Hardy-Weinberg平衡状态外(P<0.05),在其余位点都处于Hardy-Weinberg平衡状态。
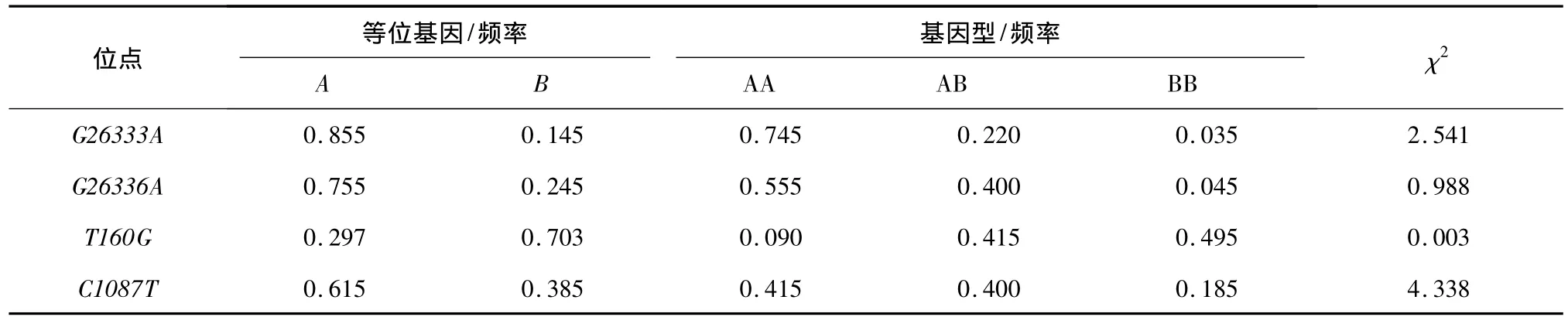
表2 IGF1R、IGFBP-3基因基因型和等位基因频率Table 2 Genotype and allele frequency of IGF1R and IGFBP-3 genes
2.4 IGF1R基因 G26336位点和 IGFBP-3基因C1087T位点与生产性能的关联分析
由表3可知,DD基因型个体在8周龄时体质量显著高于CC基因型(P<0.05),DD基因型个体在12周龄时体质量显著高于CC和CD基因型(P<0.05)。由表4可知,GH基因型个体的开产日龄显著低于HH基因型(P<0.05)。
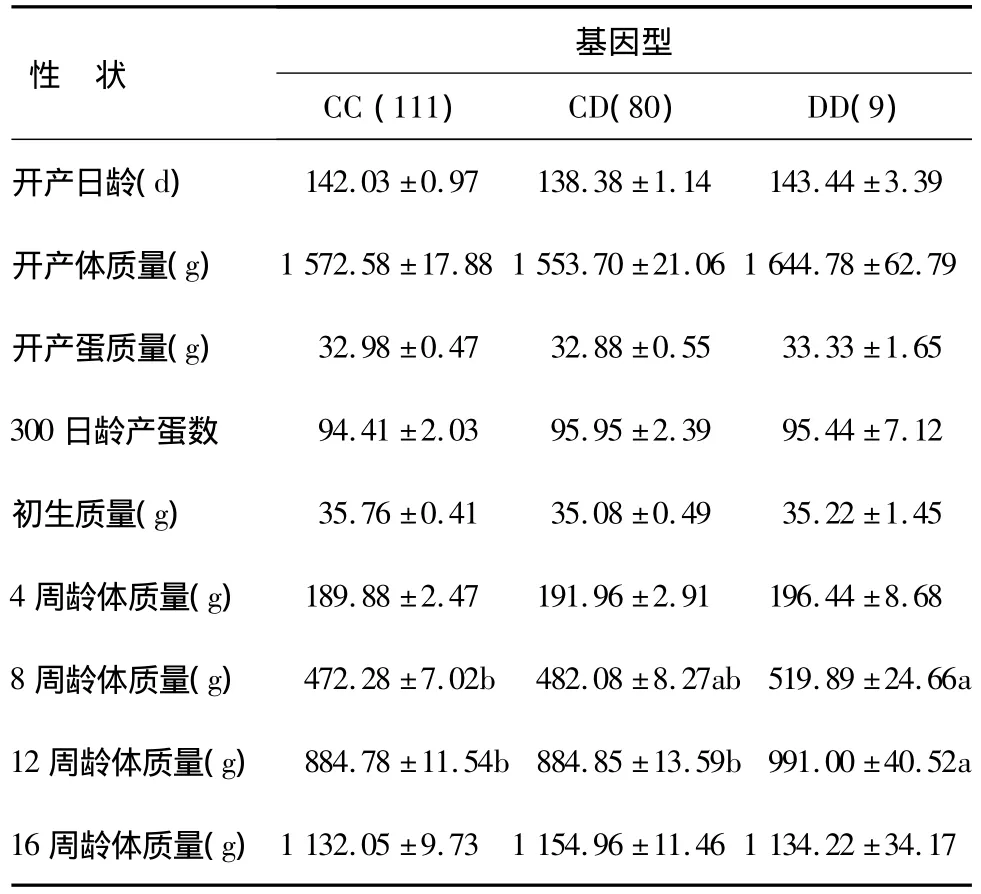
表3 G26336位点基因型与鸡生产性能的相关性Table 3 Association of G26336 genotypes with production performance in chickens

表4 C1087T位点基因型与鸡生产性能的相关性Table4 Association of C1087T genotypes with production performance in chickens
2.5 IGF1R基因和IGFBP-3基因对繁殖性状的互作效应
IGF1R基因与IGFBP-3基因对繁殖性状的互作效应见表 5。由表 5可知,G26333A×T160G、G26333A×C1087T、G26336A×T160G和G26336A×C1087T对开产日龄、开产体质量、开产蛋质量及300日龄产蛋数均无显著影响(P>0.05)。故IGF1R基因与IGFBP-3基因对盐城地方草鸡的繁殖性状不存在互作。

表5 IGF1R基因和IGFBP-3基因对鸡繁殖性状的互作效应Table 5 Interaction effect of IGF1R and IGFBP-3 genes on reproductive traits in chickens
2.6 IGF1R基因和IGFBP-3基因对生长性状的互作效应
IGF1R基因和IGFBP-3基因对生长性状的互作效应见表6。由表6可见,IGF1R基因和IGFBP-3基因对盐城地方草鸡生长性状存在一定互作,IGF1R基因G26336A位点与IGFBP-3基因C1087T位点对12周龄体质量存在互作效应。
2.7 IGF1R和IGFBP-3基因型组合对生产性状的互作效应分析
G26336A与C1087T位点基因型组合对生长性状的互作效应见表7。G26336A和C1087T位点共检测到9种基因型组合。由于DD/HH个体数较少(1只),不参与多重比较。CC/GG和CC/HH基因型组合个体的开产日龄极显著高于CD/GH(P<0.01),CC/GH基因型组合个体的初生质量显著高于CD/GG(P<0.05),DD/GG基因型组合个体的8周龄体质量显著高于CC/HH(P<0.05),DD/GG基因型组合个体的12周龄体质量极显著高于CC/GG、CC/GH、CC/HH、CD/GG、CD/GH 和CD/HH(P <0.01)。

表6 IGF1R基因和IGFBP-3基因对鸡生长性状的互作效应Table 6 Interaction effect of IGF1R and IGFBP-3 genes on growth traits in chickens
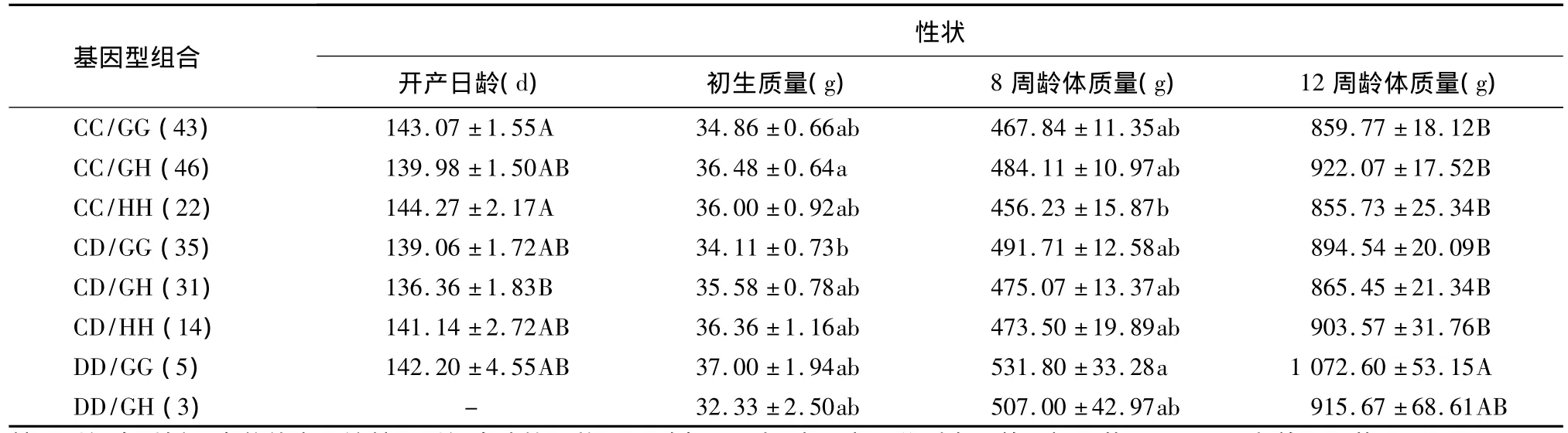
表7 IGF1R基因G26336A位点和IGFBP-3基因C1087T位点不同基因型组合与鸡生长性状的相关性Table 7 The relationship between different genotype combinations of IGF1R and IGFBP-3 genes and growth traits in chickens
基因型组合后括号内数值表示该基因型组合鸡的只数。同列中不同大、小写字母分别表示差异极显著(P<0.01)和差异显著(P<0.05)。
3 讨论
在IGF1R基因G26336A位点上,DD基因型个体的8周龄体质量显著高于CC基因型个体(P<0.05),12周龄体质量显著高于CC和CD基因型个体(P<0.05);从4、8、12周龄体质量看,具有DD>CD>CC的趋势;等位基因D是体质量性状的增效基因,等位基因C是体质量性状的减效基因。在IGFBP-3基因C1087T位点上,GH基因型个体的开产日龄显著低于HH基因型(P<0.05)。
培育具有综合性能的优良家禽品种已成为现代家禽育种的重要目标。由于家禽许多经济性状是多基因控制的数量性状,在育种中通过分子标记选择含有多个目标基因的个体,从中再选出优良的个体,实现有利基因的聚合,使品种具有更优良的生产性能。目前,通过分子标记将多个影响家禽生产性状的基因聚合在一起的研究报道不多。赵秀华[9]在京海黄鸡群体中对IGFBP-1、IGFBP-2和STAT5基因进行了三基因互作效应分析,结果显示三基因合并效应>两基因合并效应>单基因效应,而且组合基因型效应不是单个基因型效应的简单相加,其效果要优于单个基因型的效应。李国辉等[10]以白耳鸡为试验对象,检测GH、POU1F1和PRL基因单核苷酸多态性,分析这3个基因的单基因效应与聚合基因型效应。生产上可通过选择影响生产性状的2个基因的SNP,进行分子标记辅助育种,这比用单个基因进行选择的风险小。多基因的基因型组合效应不是单基因的基因型效应的简单相加,而是高于最好的单基因的基因型效应。这提示我们,在分析多基因调控的数量性状时,需要同时对多个位点组成的基因型组合进行分析,这样才能得到基因与性状之间的真实相关性。
本研究IGF1R基因G26336A位点与IGFBP-3基因C1087T位点基因型组合中,CC/GG和CC/HH个体开产日龄最大;CC/GH个体初生质量最大;DD/GG个体8周龄体质量最大;DD/GG个体12周龄体质量最大,均值为 (1 072.60±53.15)g,极显著地高于其他基因型组合个体。提示G26336A×C1087T基因型组合中剔除CC/GG和CC/HH型个体可减小开产日龄,选留CC/GH型个体可提高初生质量,选留DD/GG型个体可提高8周龄和12周龄体质量。
[1] 张 力,李树浓.胰岛素样生长因子-1[J].国外医学生理、病理科学与临床分册,1997,17(2):174-176.
[2] MOE H H,SHIMOGIRIT,KAMIHIRAGUMA W,et al.Analysis of polymorphisms in the insulin-like growth factor 1 receptor(IGF1R)gene from Japanese quail selected for body weight[J].Animal Genet,2007,38(6):659-61.
[3] 高凤华,卞立红,李 辉,等.鸡IGF1R基因多态性与生长和体组成性状的相关性研究[J].东北农业大学学报,2009,40(1):77-83.
[4] 雷明明,彭 霞,张细权.鸡IGFI-型受体基因单苷酸多态与鸡生长性能的相关分析[C]//中国畜牧兽医学会.第十次全国畜禽遗传标记研讨会论文集,2006:121-124.
[5] 高 雪,徐秀容,许尚忠.牛类胰岛素生长因子结合蛋白3基因PCR-SSCP分析[J].农业生物技术学报,2006,14(4):474-477.
[6] 胡沈荣,于 娇,蓝贤勇,等.西农萨能奶山羊IGFBP-3多态与产羔数、体尺性状的关联分析[J].西北农林科技大学学报:自然科学版,2007,35(11):1-5.
[7] 刘 哲,吴建平,马彦男,等.奶牛IGFBP-3基因部分序列PCR-SSCP多态性与产奶量和生长性能的相关性[J].农业生物技术学报,2009,17(3):445-450.
[8] 王金玉,陈国宏.数量遗传与动物育种[M].南京:东南大学出版社,2004.
[9] 赵秀华.京海黄鸡IGFBP-1、IGFBP-2和STAT5基因遗传效应及表达规律的研究[D].扬州:扬州大学,2012.
[10]李国辉,张学余,苏一军,等.3个基因SNPs及基因聚合对白耳鸡产蛋数的遗传效应[J].江西农业大学学报,2010,32(4):661-667.
猜你喜欢
动物营养学报(2022年11期)2022-12-08 08:23:14
特产研究(2021年6期)2021-12-08 03:16:38
智慧健康(2021年17期)2021-07-30 14:38:32
河南农业科学(2019年9期)2019-09-24 11:49:32
现代检验医学杂志(2016年5期)2016-08-20 03:16:54
农村百事通(2016年6期)2016-05-14 00:27:38
法医学杂志(2015年4期)2016-01-06 12:36:40
现代检验医学杂志(2015年6期)2015-02-06 01:44:02
实验动物与比较医学(2014年5期)2014-02-28 14:53:10
分子诊断与治疗杂志(2013年3期)2013-07-08 02:17:11
