
氮素处理对2种丁香的生长和光合特性的影响1)
2015-03-06 09:16王美杰胡举伟逄好胜张会慧金销王美菊王晶英
东北林业大学学报 2015年5期
王美杰 胡举伟 逄好胜 张会慧 金销 王美菊 王晶英
(东北林业大学,哈尔滨,150040) (黑龙江省科学院自然与生态研究所) (东北林业大学)
责任编辑:程 红。
丁香属(Syringa)植物属于木犀科灌木或小乔木,喜光耐阴,耐寒耐旱,适应性较强,是黑龙江园林绿化中不可缺少的树种。氮素营养是植物的生命元素,根系吸收的氮素营养以铵态氮与硝态氮2种氮素形态为主[1]。由于铵态氮与硝态氮的理化性质不同,植物体的吸收及利用能力也不同[2],所以不同氮形态对植物的生长及生理特性的影响也有明显差异[3-4]。研究表明,不同氮素处理可影响植物氮代谢有关酶的活性[5]、含氮化合物的合成[6]、养分的吸收[7]等生理活动,综合表现为对植物生长和光合性能的影响,如目前关于不同氮素处理对落叶松幼苗(Larix gmelinii)[8]、桑树(Morus alba)[9]和水曲柳(Fraxinus mandschurica)[10]等多种植物光合和生理特性的影响均有报道,但关于丁香的这方面的研究较少。光合作用是研究植物生理生态过程对环境响应的基础[11]。本试验研究了不同氮素水平和形态比例对紫丁香(Syringa oblata)和小叶丁香(Sytinga microphylla)的生长及光合参数对光强的响应,以期为紫丁香和小叶丁香的合理施肥提供理论依据。
1 材料与方法
1.1 试验材料
试验于2014年5—7月在东北林业大学植物生理学实验室进行,供试材料为1年生的紫丁香和小叶丁香幼苗,苗高为30 cm左右,于2014年5月移入盆中,采用经洗涤的细沙作为栽培基质,挑选生长近似的紫丁香和小叶丁香幼苗分为2批,每批5组,每组3盆,每盆4株,生长期间浇Hoagland完全营养液,每3 d浇一次。于2014年6月15日先用清水洗涤栽培基质,在Hoagland营养液的基础上,一批进行不同氮素水平处理,固定m(铵态氮)∶m(硝态氮)=1∶1,分别设5个氮素水平(0、10、20、30、40 mmol·L-1),以1、2、3、4、5表示,设处理1为对照组;一批进行不同氮素形态比例的处理,固定营养液中的总氮量为20 mmol·L-1,m(铵态氮)∶m(硝态氮)分别为4∶0、3∶1、2∶2、1∶3、0∶4,以6、7、8、9、10表示。每3 d浇灌一次营养液,处理2周后进行各项指标测定。
1.2 测定指标与方法
试验期间,分别于处理前和处理最后一天对各处理每株幼苗进行苗高(H)和地径(D)的测定,每株测定2次取平均值,2次测量的差值记为各处理幼苗的株高和地径的增量。
光响应曲线测定:试验处理2周后,应用LI-6400便携式光合系统分析仪(LI-COR,Inc,USA)对不同氮素处理两周以后的2种丁香进行测定,测定时间选择在09:00—12:00。叶片选取上数第5片叶片。测定时,在自然CO2摩尔质量分数条件下(约375μmol·mol-1),叶片温度控制在(29±0.5)℃,利用仪器配备的红、蓝人工光源(Li6400-02B LED),设定光有效辐射强度(PAR)由高到低依次为1 000、800、600、400、200、100、50、20、0μmol·m-2·s-1,每个光强下适应2 min后测定,按照光照强度(PFD)由高到低的顺序分别测定,每个处理重复测定3组叶片;仪器自动记录净光合速率(Pn)、蒸腾速率(Tr)指标。
光合拟合模型的选择:植物的各种光响应参数不能直接测量,只能通过一些经验模型或机理模型的拟合来得到。虽然叶片光合作用对环境因子的响应可以用光合作用、蒸腾作用、气孔导度的耦合模型模拟[12],但即使是使用其中的生化模型最终也要用直角双曲线或相近的非直角双曲线来描述光合作用或电子传递对光强的响应。本研究采用双曲线模型来进行拟合。直角双曲线的表达式为:
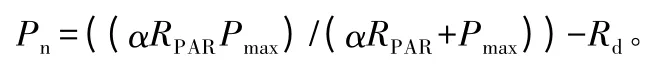
式中:Pn为净光合速率;α为表观量子效率;RPAR为光合有效辐射;Pmax为最大光合速率;Rd为暗呼吸速率。
数据处理:采用WPSExcel和DPS软件进行数据处理。
2 结果与分析
2.1 氮素水平对2种丁香的影响
2.1.1 对生长的影响
由表1可以看出:处理4紫丁香的株高增量最大,为1.79 cm,比对照组增加了94.6%,差异显著,但与处理3、5差异不显著;处理4的地径增量最大,为4.08×10-2cm,比对照组增加了25.9%,差异显著,但与处理3、5差异不显著。处理5的小叶丁香株高增量最大,为2.90 cm,比对照组增加了33.6%,差异显著,但处理1、2、3、4差异不显著,处理2、3、4、5差异不显著;处理5的地径增量最大,为3.39×10-2cm,较对照组增加了11.8%,差异显著,但与处理3、4差异不显著。

表1 不同氮素水平处理下紫丁香和小叶丁香的生长量
2.1.2 对光合特性的影响
由图1可以看出,不同氮素水平处理下紫丁香的Pn、Tr均随PFD的增加而升高,两者达到光饱和点后均趋于平稳。处理4的Pn、Tr值均高于其他4组,两者达到光饱和点后均趋于平稳。由此,m(铵态氮)∶m(硝态氮)=1∶1、氮素水平为30 mmol·L-1的处理可以提高紫丁香的光合能力。而小叶丁香的Pn、Tr均随PFD的增加而升高,两者达到光饱和点后均趋于平稳。处理5的Pn、Tr值均高于其他4组。由此,可以得出:m(铵态氮)∶m(硝态氮)=1∶1,氮素水平为40 mmol·L-1,可以提高小叶丁香的光合能力。经分析可以得出:当2种氮素形态比例是固定的时候,紫丁香最强的光合能力表现在氮素水平为30 mmol·L-1时,小叶丁香最强的光合能力表现在氮素水平为40 mmol·L-1时。这说明,紫丁香和小叶丁香对氮素水平的利用是有一定要求的,并非越高越好。
由表2可以看出:处理4的紫丁香表观量子效率(α)的值最大,为6.90,比对照组提高了182%;其最大净光合速率(Pmax)也是最大,为6.75μmol·m2·s-1。处理5的小叶丁香的α值最大,为5.03,比对照组提高了120%;其Pmax也为最大,为14.21。由此可知,改变施用氮肥的浓度可以改变2种丁香的气体交换参数,选择最优的施肥浓度,不仅可以避免对氮的浪费,也可以提高其光合产量。而2种丁香的暗呼吸速率与净光合速率成正相关,处理4的暗呼吸为最大。由此可见,适量的氮营养的吸收可促进植物代谢,因而对紫丁香和小叶丁香的暗呼吸也有促进作用。

图1 不同氮素水平处理下紫丁香和小叶丁香光合气体交换参数的响应曲线

表2 不同氮素水平对紫丁香和小叶丁香叶片气体交换参数的影响
2.2 氮素形态对2种丁香的影响
2.2.1 对生长特性的影响
由表3可以看出,紫丁香处理9的株高增量最大著,;为处理1.7 9 9的 c m地,径比增对量照最组大增,为加了2.8 235×.120%-2,c差m,异比不对显照组增加了25.8%(P>0.05)。小叶丁香处理8的株高增量最大,为3.07 cm,比对照组增加了46.5%,差异显著;处理8的地径增量最大,为3.25×10-2cm,比对照组增加了71%,差异显著。
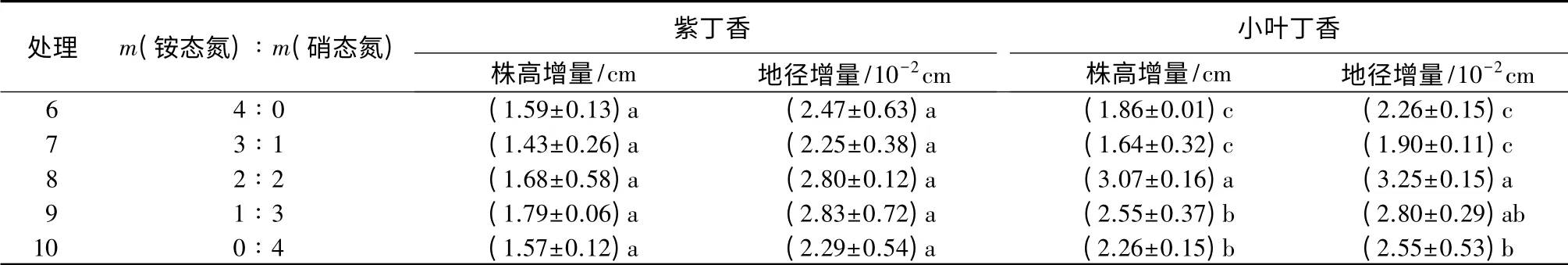
表3 不同氮素形态处理下紫丁香和小叶丁香的生长量特性
2.2.2 对光合特性的影响
由图2可以看出,不同氮素形态处理下紫丁香的Pn、Tr均随PFD的增加而升高,两者达到光饱和点后均趋于平稳。处理9的Pn、Tr值均高于其他4组,两者达到光饱和点后均趋于平稳。由此,氮素水平为20 mmol·L-1,m(铵态氮)∶m(硝态氮)=1∶3的处理可以提高紫丁香的光合能力。不同氮素形态处理下小叶丁香的Pn、Tr均随PFD的增加而升高,两者达到光饱和点后均趋于平稳。处理8的Pn、Tr值均高于其他4组。由此,可以得出:氮素水平为20 mmol·L-1,m(铵态氮)∶m(硝态氮)=1∶3时提高了紫丁香的光合能力;氮素水平为20 mmol·L-1,m(铵态氮)∶m(硝态氮)=1∶1的处理可以提高小叶丁香的光合能力。
由图2可以得出:当氮素水平是固定的时候,紫丁香最强的光合能力表现在m(铵态氮)∶m(硝态氮)=1∶3时,小叶丁香最强的光合能力表现在m(铵态氮)∶m(硝态氮)=1∶1时。这可以说明,相对多的硝态氮更能提高紫丁香的光合能力,而施用等量的铵态氮和硝态氮则可以提高小叶丁香的光合能力。

图2 不同氮素形态处理下紫丁香和小叶丁香光合气体交换参数的响应曲线

表4 不同氮素比例对紫丁香和小叶丁香叶片气体交换参数的影响
由表4可以看出,处理9的紫丁香α值最大,其 Pmax和Rd也为最大值,其对强光的利用能力也是最大的。混合施用氮肥,较高浓度的硝态氮显著提高了紫丁香的α值,比对照组提高了85%。处理8的小叶丁香α值最大,其Pmax和Rd也为最大值。施用等比例的铵态氮和硝态氮显著提高了小叶丁香的α值,比对照组提高了79%。而2种丁香的Rd与Pn成正相关,由此可以说明氮的吸收对紫丁香和小叶丁香的暗呼吸有促进作用。紫丁香和小叶丁香的光饱和点与补偿点并不因氮源的不同而改变。而2种丁香的暗呼吸速率与净光合速率成正相关,由此可以说明植株对不同氮素形态的吸收对2种丁香的暗呼吸有促进作用。
3 结论与讨论
有研究表明,增大施氮水平可显著提高落叶松的光合水平。在本试验的氮素水平的试验处理中,紫丁香和小叶丁香对不同处理表现出的生长和光合特性响应表明,紫丁香和小叶丁香在铵态氮和硝态氮等量的情况下,对氮肥的吸收是随浓度的增长而变化的,但紫丁香对氮肥的吸收能力弱于小叶丁香。同时,各种处理中,暗呼吸速率对于氮的吸收量呈现正相关,也表明,氮的吸收对紫丁香和小叶丁香的光合作用和呼吸作用的影响显著。
光合作用表观量子效率(α)表示每吸收一个光量子能引起CO2净同化的数目。在自然条件下α的值远小于理论上限,对于长势良好的作物一般在0.04~0.07[13],而直角双曲线给出的拟合结果会偏高。植物叶片的光饱和点与光补偿点反映了植物对光照条件的要求,光补偿点是衡量植物利用弱光能力大小的重要指标,光饱和点是衡量植物利用强光能力大小的指标[14]。光补偿点较低、光饱和点较高的植物对光环境的适应性较强;而光补偿点较高、光饱和点较低的植物对光照的适应性较弱[15]。暗呼吸速率主要受植物生长状态和温度的影响。而有研究表明氮的吸收对地肤(Kochia scoparia)暗呼吸有显著的促进作用[16]。
植物对氮素形态的喜好因遗传种性、生态适应性和土壤环境的不同而不同。茶树(Camellia sinensis)[17]、云南松(Pinus yunnanensis)[18]对铵态氮的吸收效率明显高于硝态氮,单纯供应铵态氮2种植物的生物量最大。越橘(Vaccinium corymbosum)也明显偏好铵态氮,施用铵态氮促进了植株生长,也提高了其净光合速率[19]。而高山松(Pinus densata)和油松(P.tabulaeformis)明显偏好硝态氮,在以铵态氮为氮源时,2个树种的生物量积累和光合活性都显著下降[18]。但是,具体树种的适宜氮形态配比至今并未掌握其规律。大量的试验均表明,硝态氮和铵态氮具有联合效应,混合施加时有利于维持细胞的电性平衡和pH的平衡。同时供应时,几乎所有植物都生长得较好[20]。
氮素形态几乎影响了光合作用的各个环节,包括影响叶片叶绿素含量、光合速率、暗反应主要酶活性以及光呼吸等,直接或间接影响着光合作用。氮素形态对植物叶片的光合性能的影响与植物的种类有关。有研究发现铁核桃(Juglans sigillata)实生苗在硝态氮处理下的光合色素含量以及净光合速率明显低于铵态氮处理[21],并且在红松(Pinus koraiensis)幼苗的研究中也表现出相似的结果[22],而柚(Citrus maxima)在硝态氮处理下的净光合速率明显高于铵态氮[23]。
在氮素形态的试验处理中,紫丁香和小叶丁香对不同处理表现出的生长特点表明,紫丁香的各个处理的差异不显著,即紫丁香的生长对氮素形态没有特别的偏好;而光合特性响应表明,紫丁香和小叶丁香对于硝态氮和铵态氮的混合施用的吸收程度远高于单独施用某种氮肥。同时,各种处理中,暗呼吸速率对于氮的吸收量呈现正相关,也表明氮的吸收对紫丁香和小叶丁香的光合作用和呼吸作用的影响显著。
综上所述,在施氮总量一定的条件下,施用氮素的水平为30 mmol·L-1更有利于紫丁香的生长和光合作用;在10、20、30、40 mmol·L-1施氮处理中,水平为40 mmol·L-1时更有利于小叶丁香的生长和光合作用。在氮肥量一定的条件下,施用m(铵态氮)∶m(硝态氮)=1∶3的氮肥,更有利于紫丁香的光合作用;施用m(铵态氮)∶m(硝态氮)=2∶2的氮肥更有利于小叶丁香的光合作用。即紫丁香更偏好硝态氮,小叶丁香对2种形态的氮营养的没有特别偏好;从生长量而言,不同的氮素形态对紫丁香没有显著影响,但小叶丁香更倾向于硝态氮。不同的丁香应采用不同的氮肥施用方案,才能更利于丁香的生长,才能发挥丁香的环境和经济效益。
[1]Burke I C,Lauenroth W K,Parton W J.Regional and temporal variation in net primary production and nitrogen mineralization in grasslands[J].Ecology,1997,78(5):1330-1340.
[2]张延春,陈治锋,龙怀玉,等.不同氮素形态及比例对烤烟长势、产量及部分品质因素的影响[J].植物营养与肥料学报,2005,11(6):787-792.
[3]Xie H L,Jiang R F,Zhang F S,et al.Effect of nitrogen form on the rhizosphere dynamics and uptake of cadmium and zinc by the hyperaccumulator Thlaspi caerulescens[J].Plant and Soil,2009,318(1/2):205-215.
[4]刘赵帆,张国斌,郁继华,等.氮肥形态及配比对花椰菜产量、品质和养分吸收的影响[J].应用生态学报,2013,24(7):1923-1930.
[5]Gashaw L,Mugwira L M.Ammonium-N and nitrate-N effects on the growth and mineral compositions of triticale,wheat,and rye[J].Agronomy Journal,1981,73(1):47-51.
[6]陈锦强,李明启.不同氮素营养对黄麻叶片的光合作用、光呼吸的影响及光呼吸与硝酸还原的关系[J].植物生理学报,1983,9(3):251-259.
[7]Engels C,Marschner H.Influence of the form of nitrogen supply on root uptake and translocation of cations in the xylem exudate of maize(Zea mays L.)[J].Journal of Experimental Botany,1993,44(11):1695-1701.
[8]郭盛磊,阎秀峰,白冰,等.供氮水平对落叶松幼苗光合作用的影响[J].生态学报,2005,25(6):1291-1298.
[9]许楠.黑龙江省桑树氮肥运筹及光合调控机理的研究[D].哈尔滨:东北林业大学,2013.
[10]孙海龙.氮磷营养对水曲柳苗木光合作用和氮同化的影响[D].哈尔滨:东北林业大学,2005.
[11]张中峰,黄玉清,莫凌,等.岩溶植物光合—光响应曲线的两种拟合模型比较[J].武汉植物学研究,2009,27(3):340-344.
[12]于强,王天铎,刘建栋.玉米株型与冠层光合作用的数学模拟研究Ⅰ:模型与验证[J].作物学报,1998,24(1):7-15.
[13]Long SP,Humphries S,Falkowski P G.Photoinhibition of photosynthesis in nature[J].Annual Review of Plant Biology,1994,45(1):633-662.
[14]叶子飘.光合作用对光和CO2响应模型的研究进展[J].植物生态学报,2010,34(6):727-740.
[15]杨兴洪,邹琦,赵世杰.遮荫和全光下生长的棉花光合作用和叶绿素荧光特征[J].植物生态学报,2005,29(1):8-15.
[16]曹勋,郑新军,李彦,等.氮过量吸收对地肤暗呼吸速率及相对生长速率的影响[J].生态学杂志,2013,32(8):2002-2007.
[17]Ruan Jianyun,Gerendás J,Härdter R,et al.Effect of nitrogen form and rootzonepH on growth and nitrogen uptake of tea(Camellia sinensis)plants[J].Annals of Botany,2007,99(2):301-310.
[18]Yao Buqing,Cao Jing,Zhao Changming,et al.Influence of ammonium and nitratesupply on growth,nitrate reductase activity and N-use efficiency in anatural hybrid pine and its parents[J].Journal of Plant Ecology,2011,4(4):275-282.
[19]李亚东,赵爽,张志东,等.不同氮素形态配比对越橘生长、产量及叶片元素含量的影响[J].吉林农业大学学报,2008,30(4):477-480.
[20]葛会敏,樊卫国.不同形态氮源及其配比对石灰性黄壤上纽荷尔脐橙光合特性的影响[J].中国生态农业学报,2013,21(4):401-408.
[21]樊卫国,葛慧敏,吴素芳,等.不同形态氮素及其配比对铁核桃实生苗叶片光合特性和养分含量的影响[J].果树学报,2013,30(3):437-443.
[22]陈永亮,刘明河,李修岭.不同形态氮素配比对红松幼苗光合特性的影响[J].南京林业大学学报:自然科学版,2005,29(3):77-80.
[23]孙谷畴,曾小平,赵平,等.不同氮源下生长的柚树叶片光合参数对高浓度CO2驯化作用的比较[J].生态学报,2003,23(1):14-21.
猜你喜欢
河南师范大学学报(自然科学版)(2022年3期)2022-04-14
雪豆月读·低年级(2021年6期)2021-09-10
华北水利水电大学学报(自然科学版)(2020年6期)2021-01-29
农业环境科学学报(2020年10期)2020-11-11
江苏农业科学(2020年12期)2020-07-20
西安科技大学学报(社会科学版)(2019年1期)2019-09-10
北方音乐(2017年1期)2017-03-02
小学生作文选刊·低年级版(2016年7期)2016-08-06
新课程·中旬(2014年10期)2014-12-26
小学生·多元智能大王(2014年2期)2014-03-21
