
供氮方式及水平对米老排苗期生长动态的影响1)
2015-03-06 09:16闫彩霞杨锦昌尹光天李荣生邹文涛
东北林业大学学报 2015年5期
闫彩霞 杨锦昌 尹光天 李荣生 邹文涛
(中国林业科学研究院热带林业研究所,广州,510520)
责任编辑:任 俐。
施肥是苗木培育过程中促进苗木营养生长、提高苗木产量和质量、优化造林效果的重要途径[1-2]。作为植物生长所需的大量营养元素,氮素是植物体内许多重要有机化合物的组分[3],对苗木生长有重要影响,氮供给充足的植株生长较快[4-5],缺乏时常使植株的生长受到抑制,同时是影响苗木生长发育、产量和品质的重要因素。合理的施肥方法能够提高苗木氮素利用效率、产量、品质,增强生存竞争力,节省肥料,对保护生态环境也非常重要,因此,其一直是林木营养的热点[6]。
等量施肥法是在生长期内重复施用相同剂量的肥料[7],又称衡量施肥或传统施肥。阶段递增施肥是在连续的生长阶段以递增函数的方式确定施肥量的方法。修正指数施肥法是基于植物稳态矿质营养平衡理论、按照修正指数施肥模型来确定施肥量的方法。三者的施肥总量和频率相似,但施肥量的周期加载率不同[8]。修正指数施肥法在苗木组织养分吸收和储藏上比传统施肥法效率更高[9-10]。朱存福等[11]研究了不同施肥方法和水分胁迫对台湾相思(Acacia confusa)苗木生长的影响,发现常规浇水时苗木总生物量由大到小的顺序为指数施肥、平均施肥、直线施肥、CK(不施肥),而平均施肥在水分胁迫的条件下效果最佳。Chen et al.[12]比较了传统施肥(50 C和100 C)和指数施肥(50 E、100 E、200 E、400 E)对米老排(Mytilaria laosensis Lecomte)生长和养分吸收动态的影响,认为100 C对增加植株生物量和养分质量分数效果更佳。通过适当增加初期施入量和减少后期施入量,修正指数施肥法能克服指数施肥法初期肥量不足和后期肥量积累引发生理肥害的缺陷,因此,它比指数施肥法更有优势。但国内外对指数施肥方法在林业实践上的报道日益增多,但对修正指数施肥方法的研究少之又少。
米老排别名马蹄荷、壳菜果、山油桐、三角枫等,金缕梅科壳菜果属常绿阔叶大乔木,是中国、老挝、越南的特有种,在我国天然分布于广东西部、广西西南部和云南东南部。近年来,鉴于其良好的开发利用价值和广泛的应用前景,米老排日益受到人们关注,并在生产上不断得到推广应用[13]。目前国内外对于米老排的研究主要集中在分类学[14-17]、木材特性[18-20]、混交林营造技术[21-22]和人工林生态功能[23-25]等方面,鲜见有关于不同施肥方法及施用量对米老排苗期生长影响的报道。本试验采用3种施肥方法,即等量施肥、阶段递增施肥和修正指数施肥法,并设置不同氮素施用量水平对米老排进行苗期施肥试验,通过定期测量苗高、地径和生物量等指标,分析研究不同施肥方法对米老排苗期营养生长动态的影响,旨在为提高米老排苗期产量及质量、优化苗圃生产施肥技术和改善其造林效果提供参考。
1 材料与方法
1.1 试验材料
试验于2013年5—11月份在中国林业科学研究院热带林业研究所温室大棚中进行。供试米老排幼苗为实生苗,种子于2012年采自我国广西靖西。
2013年5月末播种发芽后选取长势良好、大小均匀一致、苗高约10 cm的幼苗,用去离子水洗净后,移栽于规格为10 cm×8 cm×10 cm(上口×下口×高)的塑料盆中(体积为600 mL);培养基质为经高压灭菌的混合基质(V(泥炭)∶V(蛭石)∶V(珍珠岩)=3∶2∶2)。为了防止水肥流失,盆内套有双层白色塑料袋。
参试肥料为普罗丹高质量分数水溶性复合肥(Planta,质量分数w(N)∶w(P)∶w(K)=20∶20∶20,加拿大安大略省宾顿市植物产品有限公司),以去除潜在的由施肥、灌溉条件对苗木生长和生理产生的交互影响。其主要养分质量分数分别为N20%,P2O520%,K2O20%,螯合铁(Fe)0.10%,螯合锰(Mn)0.05%,螯合锌(Zn)0.05%,螯合铜(Cu)0.05%,硼(B)0.02%,钼(Mo)0.000 5%,螯合体(EDTA)1.00%。
1.2 试验设计与措施
移苗前,根据Timmer et al.[26]的方法确定基质的最大持水量,以最大持水量的75%作为移苗时的初始水量;此后每周浇水,浇水量视天气和苗木生长状况进行适当调整,保证所有苗木水分条件一致。移苗时,先用自来水冲洗幼苗,再用去离子水将幼苗冲洗干净,将其移栽到塑料盆中;为减少边际效应,定期移动苗盘。试验期间,温室未使用人工光源,日平均温度28℃,日平均湿度67%。
采用随机区组设计,设置3种施肥方式(即等量施肥(C)、阶段递增施肥(A)和修正指数施肥(ME)),每种施肥方式设置50、100、200、400、600 mg·株-15个施氮量水平,并以不施肥为对照(CK),总共16个处理水平,每个施肥处理24株,重复4次,共计1 536株。
试验开始时,随机抽取60株,测得幼苗平均初始N、P、K含量分别为7.58、0.65、6.71 mg·株-1。移苗后第3周开始施肥,每周施肥1次,共16次,具体施肥方案见表1。
等量施肥公式为:

式中:NT为设置的氮素施用量;t为施肥次数;Nt为每次氮素施用量。
阶段递增施肥公式为:
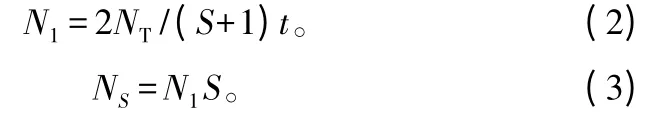
式中:N1为第1阶段施肥量;NS为第S阶段施肥量;NT为设置的氮素施用量;t为施肥次数;S为施肥阶段数。
修正指数施肥采用Birge[9]描述的方法确定施肥量,具体如下:

式中:r为从苗木在施肥处理前最初阶段的养分质量分数Ns增加至最终氮质量分数(NT+Ns)的相对增加率;NT为设置的氮素施用量;t为施肥次数。
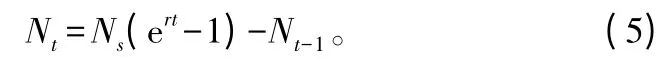
式中:Nt为第t次施肥量;Nt-1为含前一次施氮量的氮添加的累积量。
修正施肥即氮补偿,是通过增加初始氮添加和减少生长末期施用量来实现的,公式如下:

式中:NC(氮补偿)为由公式(5)得出的最后2次施氮量的差值,并按上述公式补偿到根系中;N0为补偿期间最后的氮素添加量。
1.3 取样调查与指标测定
试验期间,分别于6—10月份每个月初(即第0、5、10、15、20周)时,调查并记录所有参试苗木株高、地径(由于移苗时米老排幼苗嫩茎较脆弱,不易于用游标卡尺测量地径,故6月份时未测地径)。从7月份起每月测定一次生物量,每小区选取4个平均标准株,将幼苗用去离子水洗净,按根、茎、叶分别置于烘箱中105℃杀青20 min,然后70℃下烘48 h后称其干质量。根据以下公式计算苗木质量指数(QI)[27]。

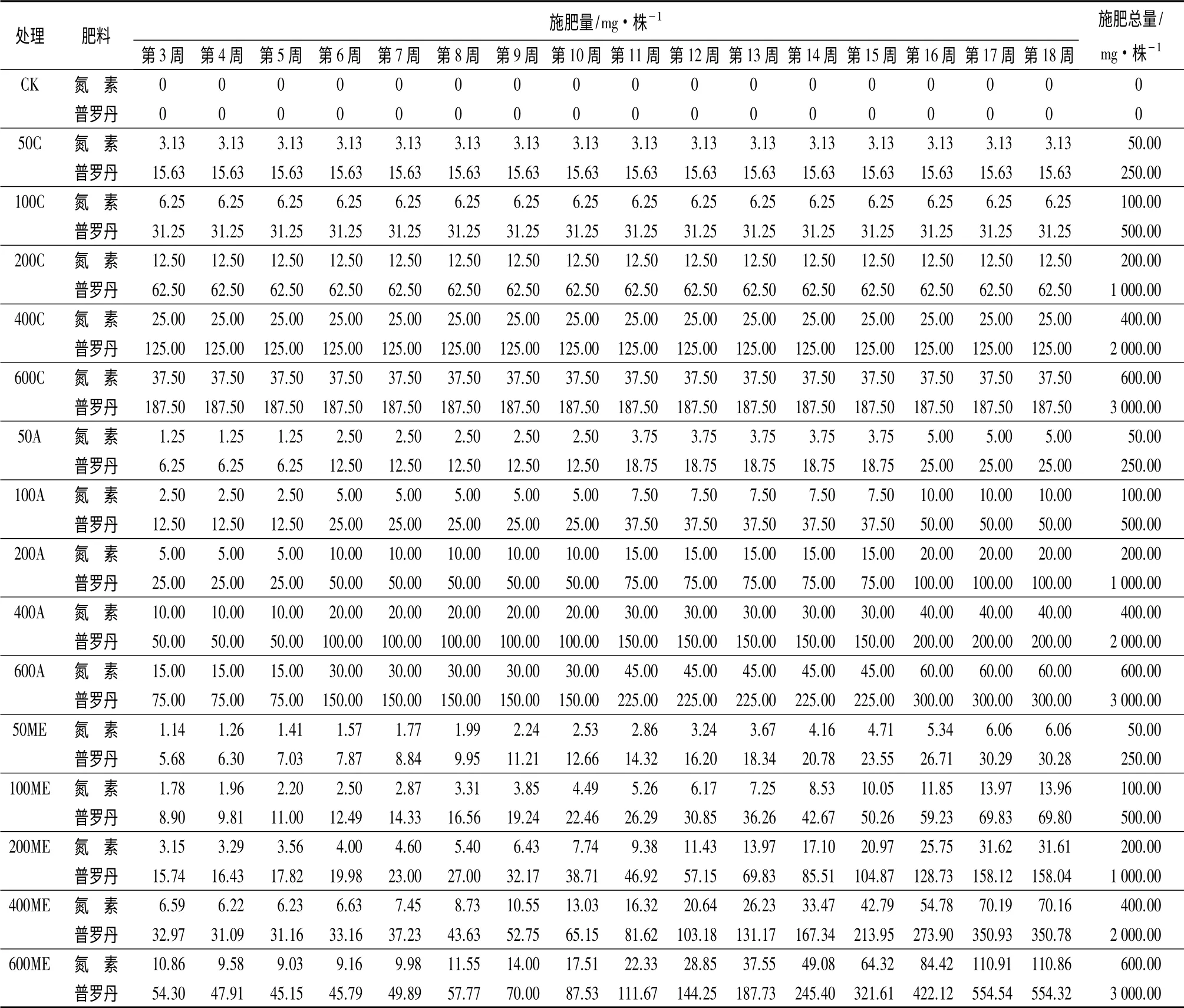
表1 米老排苗期施肥方案
1.4 数据分析
采用Excel 2007和SPSS 18.0软件对试验数据进行统计分析并绘制图表。方差分析用于检验不同施肥方法对各生长指标产生的影响,当P<0.05时,用Duncan法进行多重比较。
2 结果与分析
2.1 不同的施肥方式及施肥量对米老排苗期高生长的影响
不同的施肥方式及施肥量对米老排株高生长产生了显著影响(图1)。试验期间,对照处理苗木的株高呈经典的慢—快—慢的Logistic生长曲线。等量施肥方式下,试验前期不同施肥量对株高影响差别不大;7月份各施肥处理苗木的株高增长速度最快,其后低剂量施肥幼苗株高增速有所减缓,高剂量处理(400 C和600 C)幼苗表现更显著。对于阶段递增施肥方式,试验前期不同施肥量下幼苗高生长动态与等量施肥方式相似;试验后期,600 A米老排高生长减缓最为突出。低剂量修正指数施肥(50 ME和100 ME)的苗高生长趋势在试验期间呈“J”型,也就是说在这些苗木在试验后期依然保持高速增长状态;高剂量修正指数施肥苗高增长在9月份出现分化,高生长减缓由大到小的顺序为600 ME、400 ME、200 ME,再次验证了过量施肥对植株生长的毒害作用。
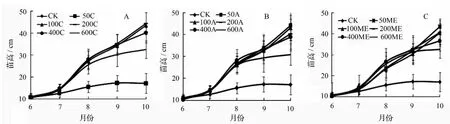
图1 不同施肥方式及施肥量下米老排苗高生长曲线
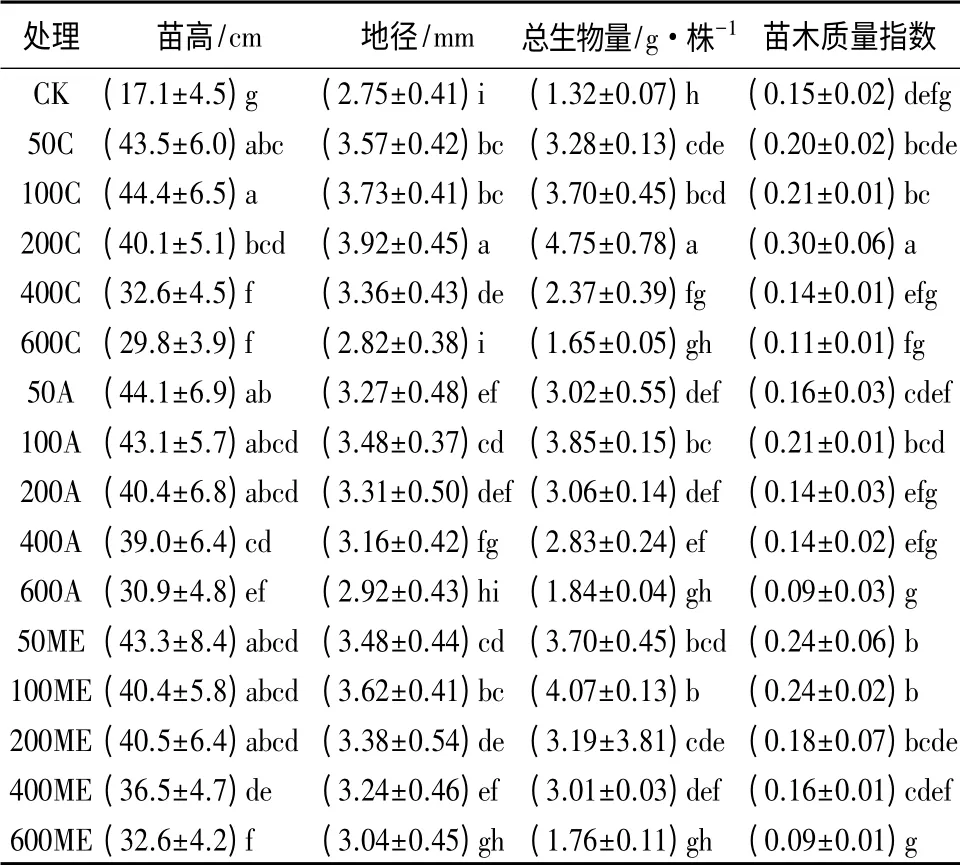
表2 不同的施肥方式及施肥量对收获时米老排生长指标的影响
从表2中可以看出,施肥结束时,对于苗高而言,除400 mg·株-1施肥量的苗木在阶段递增和修正指数施肥方式下与等量施肥显著差异外,其他施肥量在不同施肥方式间的差异均不显著。不同施肥量对应的最佳施肥方式也不同。当苗木施氮量为50 mg·株-1或400 mg·株-1时,阶段递增施肥方式均大于其他方式对株高生长的促进效果;100 mg·株-1时,以等量施肥效果最好;200 mg·株-1或600 mg·株-1时,修正指数施肥方式下米老排高生长表现最佳。
2.2 不同的施肥方式及施肥量对米老排苗期地径生长的影响
施肥方式及施肥量不同,对米老排地径生长产生的影响不同。由图2可知,在测量期间,对照苗木的直径增长速度随着试验时间的延长而减缓。等量施肥方式下,中等剂量处理苗木的直径生长持续高于其他处理。采用阶段递增方式施肥时,对于试验前期,低剂量施肥处理苗木的直径低于高剂量处理;随着施肥试验的继续,各处理苗木地径增长速度均有所减慢,低剂量施肥处理苗木的直径逐渐超过高剂量处理,但50A由于前期施肥不足后期生长依然较弱。修正指数施肥时,50 ME和100 ME幼苗地径几乎呈直线式生长,即在整个观察期,其幼苗地径均保持高速生长状态。此外,在3种施肥方式下,处理为600 mg·株-1的幼苗生长曲线均与对照一致。
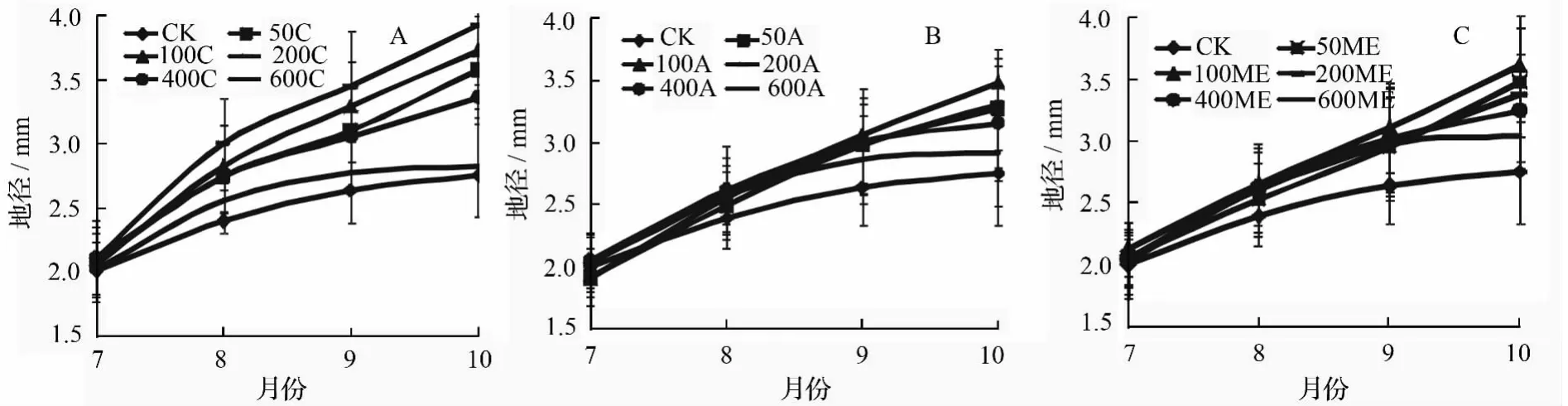
图2 不同施肥方式及施肥量下米老排地径生长曲线
试验结束时,除600 C和600 A外,其他施肥处理苗木地径均与对照形成显著差异(表2),比对照苗木地径增长了10.5%以上。200 C苗木直径达到最大值(3.92 mm),比对照增加了42.5%,与其他处理均差异显著。同等剂量下,施肥量为50 mg·株-1时,等量施肥和修正指数施肥方式均分别是阶段递增施肥方式的109.3%和106.5%,并形成显著差异。施肥量为100 mg·株-1的苗木在不同施肥方式间差异不显著。当施肥量增至400 mg·株-1时,等量施肥和阶段递增施肥间差异显著。除600 C外,等量施肥方式下不同施用量处理苗木的地径均大于其他方式,说明等量施肥方式更有利于植株地径的生长。
2.3 不同的施肥方式及施肥量对米老排苗期总生物量的影响
等量施肥时,200 C苗木各个时期总生物量均大于其他剂量处理,说明该施肥方式下中等剂量施肥有利于植株生物量积累;400 C苗木在前期增长较多,末期下降,说明该剂量能较好地满足米老排前期生长,但后期生长所需肥料不足;而600 C幼苗生物量增加在8月份后就趋于停滞。采用阶段递增方式施肥时,100 A苗木前期生物量低于其他处理,8月份进入生物量迅速积累期,至试验结束时达到最大;600 A苗木生物量积累呈现先升后降的趋势。修正指数施肥时,中低剂量处理苗木后期生物量积累大于前期,最大积累期均出现在9月份后,这是修正指数模型本身的特点决定的;600 ME苗木生物量积累模式类似于600 A苗木。
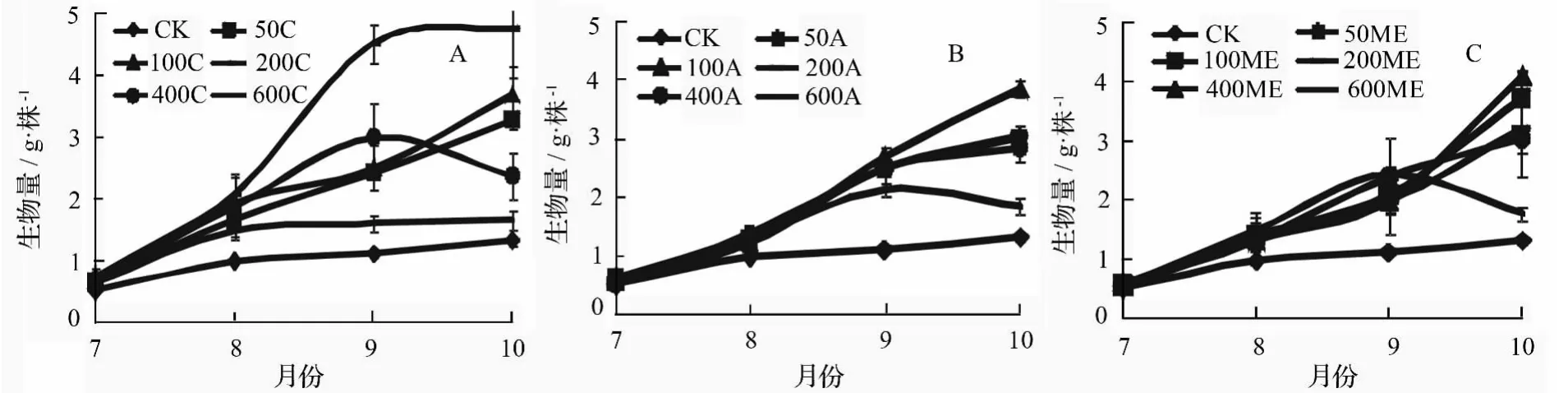
图3 不同施肥方式及施肥量下米老排生物量积累曲线
试验结束时,除3种方式最高剂量施肥苗木对米老排生物量积累促进效果不显著外,其他施肥处理苗木的生物量均与对照形成显著差异,比对照苗木生物量增长了约79.7%(表2)。200 C苗木生物量达到最大值(4.75 g·株-1),是对照苗木的360.6%,与其他处理均差异显著。阶段递增和修正指数施肥时,均以施氮量100 mg·株-1在促进生物量积累上最佳。在氮素使用量上,剂量为50、100、400 mg·株-1时均以修正指数方法施用时对生物量积累效果最好,分别至少比采用其他方式提高了32.0%、16.3%、49.0%。
2.4 不同施肥处理的米老排苗木质量指数
根据Dickson et al.[27]提出的苗木质量指数(QI)评价不同施肥方法及施用量对苗期米老排质量的影响。QI值越高则苗木质量越好。经方差分析和多重比较(表2)发现,不同施肥方式及施用量对苗期米老排质量指数的影响很大。经400 C、600 C、200 A、400 A、600 A、600 ME处理的米老排苗木质量指数均低于对照,质量指数均高于对照的施肥处理中,只有经100 C、200 C、50 ME和100 ME处理的与对照苗木形成显著差异,以经200 C处理的质量最好,其次是经100 ME和50 ME处理的,最后是经100 C处理的。
3 结论与讨论
苗木形态特征是自身遗传特性与生长环境相互作用的结果及生理状况的外在表现[28]1-2,在预测苗木大田表现上具有重要指导意义。苗木高度是体现植株地上生长状况的指标,通常与地径结合使用[29]。一些研究认为,地径能反映苗木的综合质量,最能预测造林效果[30-31]。Iverson[32]认为生物量大的苗木在困难立地条件下竞争能力更强。苗木质量指数是多个形态指标的综合评价指数,比单一形态指标更能全面地衡量苗木各部分的协调性和平衡状况[28]1-2。
了解不同施肥方式及施肥量下植株营养生长动态,有利于掌握其不同时期的需肥特性,从而为不同苗木培育目的下合理施肥技术的选择做指导。本研究发现,除低剂量修正指数施肥(50 ME和100 ME)在整个试验期间保持“J”型高速增长状态外,各处理苗木株高生长均在进入8月份后减缓。在整个观察期内,3种方式下最高剂量施肥苗木地径生长曲线均与对照一致,低剂量修正指数施肥(50 ME和100 ME)幼苗地径几乎呈直线式生长,其幼苗地径均保持高速生长状态,其他处理苗木的地径均因后期施肥不足或肥料积累而较前期生长有不同程度减缓。此外,3种方式下中低剂量施肥有利于米老排后期生物量积累,高剂量施肥导致米老排后期生物量积累停滞或下降。
试验结束时,各施肥处理米老排在苗高、地径和总生物量等指标上均大于对照苗木,分别比对照提高了73.6%、2.4%和25.6%以上,再次验证了施肥对米老排形态指标的促进作用。所有处理中,100 C的苗高最大(44.4 cm),是对照的259.0%;200 C苗木地径最大(3.92 mm),是对照的142.5%;200 C苗木生物量最大,是对照的360.6%;200 C的质量指数最大,其次是100 ME和50 ME,分别比对照提高了101.4%、59.3%和22.4%。处理600 C的苗高、地径和总生物量等均为最低,可能是因为等量施肥方式常常引起苗木早期养分毒害,且后期养分供应不足导致缺素症[33],从而影响了苗木营养生长。
苗木质量很难用单一的指标或一项独立技术评价和说明[34-35],只能反映其某个方面。文中仅从形态指标上评价了不同施肥方式及施用量对苗期米老排的影响,今后还可以从生理指标上、苗木活力及养分等方面进行综合分析评价,使之更加科学、系统。
[1]Davis A S,Jaeobs D F,Kevyn E W,et al.Organic matter added to bareroot nursery beds influences soil properties and morphology of Fraxinus pennsyIvanica and Quercus rubra seedlings[J].New Forests,2006,26(31):293-303.
[2]张金浩,周再知,杨晓清,等.氮素营养对肯氏南洋杉幼苗生长、根系活力及氮含量的影响[J].林业科学,2014,50(2):31-36.
[3]Moscatelli M C,Lagomarsino A,De Angelis P,et al.Short-and medium-term contrasting effects of nitrogen fertilization on Cand N cycling in a poplar plantation soil[J].Forest Ecology and Management,2008,255(3):447-454.
[4]De Pinheiro H A R,Marcelis L F M.Regulation of growth at steady-state nitrogen nutrition in lettuce(Lactuca sativa L.):interactive effects of nitrogen and irradiance[J].Annals of Botany,2000,86(6):1073-1080.
[5]丁晓纲,刘喻娟,张应中,等.不同氮素浓度指数施肥对银桦、蓝花楹幼苗生长及其根系和叶片的影响[J].中国农学通报,2013,29(19):39-45.
[6]李素艳,孙向阳,刘凯英.指数施肥技术在草坪培育中的应用[J].北京林业大学学报,2003,25(4):44-48.
[7]王冉,李吉跃,张方秋,等.不同施肥方法对马来沉香和土沉香苗期根系生长的影响[J].生态学报,2011,31(1):98-106.
[8]Timmer V R,Miller B D.Effects of contrasting fertilization and moisture regimes on biomass,nutrients,and water relations of container grown red pine seedlings[J].New Forests,1991,5(4):335-348.
[9]Birge Z D K,Salifu K F,Jacobs D F,et al.Modified exponential nitrogen loading to promote morphological quality and nutrient storage of bareroot-cultured Quercus rubra and Quercus alba seedlings[J].Scandinavian Journal of Forest Research,2006,21(4):306-316.
[10]McAlister JA,Timmer V R.Nutrient enrichment of white spruce seedlings during nursery culture and initial plantation establishment[J].Tree Physiology,1998,18(3):195-202.
[11]朱存福,李福秀,丁长秀.不同施肥方法对台湾相思苗木生长的影响[J].山东林业科技,2010(1):28-30.
[12]Chen Lin,Zeng Jie,Jia Hongyan,et al.Growth and nutrient uptake dynamics of Mytilaria laosensis seedlings under exponential and conventional fertilizations[J].Soil Science and Plant Nutrition,2012,58(5):618-626.
[13]闫彩霞,杨锦昌,尹光天,等.米老排不同高度级苗木形态特征的分析[J].林业资源管理,2013(5):98-102.
[14]Endress P K A.Suprageneric taxonomic classification of the Hamamelidaceae[J].Taxon,1989,38(3):371-376.
[15]Bogle A L.Multilacunar nodal anatomy in Mytilaria(Hamamelidaceae)[J].Journal of the Arnold Arboretum Harvard University,1990,71(1):111-117.
[16]潘开玉,杨亲二.花木属和壳菜果属(金缕梅科)的核型研究[J].植物分类学报,1994,32(3):235-239.
[17]施苏华,黄椰林,章群,等.壳菜果亚科(金缕梅科)与相关亚科的核糖体DNAITS区序列分析及系统发育关系[J].中山大学学报:自然科学版,1999,38(11):36-40.
[18]梁善庆,罗建举.人工林米老排木材解剖性质及其变异性研究[J].北京林业大学学报,2007,29(3):142-148.
[19]梁善庆,罗建举.人工林米老排木材的物理力学性质[J].中南林业科技大学学报,2007,27(5):97-100,116.
[20]梁善庆,罗建举.人工林米老排木材化学成分及其在树干高度上的变异[J].中南林学院学报,2004,24(5):28-31,52.
[21]唐社云.米老排与南酸枣混交造林试验报告[J].西部林业科学,1992(2):35-40.
[22]刘济祥,何伟民.米老排引种试验及繁育技术研究[J].江西林业科技,2008(1):17-19.
[23]吕福如.防火生土带改建米老排防火林带的林木生长与地力维持效果[J].河北林果研究,2002,17(4):317-322.
[24]刘恩,刘世荣.南亚热带米老排人工林碳贮量及其分配特征[J].生态学报,2012,32(16):5103-5109.
[25]明安刚,贾宏炎,陶怡,等.米老排人工林碳素积累特征及其分配格局[J].生态学杂志,2012,31(11):2730-2735.
[26]Timmer V R,Armstrong G.Growth and nutrition of containerized Pinus resinosa seedlings at varying moisture regimes[J].New Forests,1989,3(2):171-180.
[27]Dickson A,Leaf A L,Hosner J F.Quality appraisal of white spruce and white pine seedling stock in nurseries[J].The Forestry Chronicle,1960,36(1):10-13.
[28]刘勇.苗木质量调控理论与技术[M].北京:中国林业出版社,1999:1-2.
[29]孙慧彦.长白落叶松苗木质量动态评价与施肥技术研究[D].北京:北京林业大学,2011.
[30]South D B,Menzies M I,Holden D G.Stock size affects outplanting survival and early growth of fascicle cuttings of Pinus radiata[J].New Forests,2005,29(3):273-288.
[31]Dey D C,Parker W.Morphological indicators of stock quality and field performance of red oak(Quercus rubra L.)seed lings underplanted in a central Ontario shelter wood[J].New Forests,1997,14(2):145-156.
[32]Iverson R D.Planting-stock selection:meeting biological needs and operational realities[M]//Duryea M L,Landis T D.Forest nursery manual:production of bareroot seedling.Corvallis:Kluwer Academic Publishers,1984:261-266.
[33]Timmer V R,Armstrong G,Miller B D.Steady-state nutrient preconditioning and early outplanting performance of containerized black spruce seedlings[J].Canadian Journal of Forest Research,1991,21(5):585-594.
[34]Ritchie G A.Assessing seedling quality[M]//Duryea M L,Landis T D.Forest nursery manual:Production of bareroot seedlings.Corvallis:Kluwer Academic Publishers,1984:243-259.
[35]Puttonen P.Looking for the“silver buIlet”-can one test do it all?[J].New Forests,1996,13:9-27.
猜你喜欢
现代农村科技(2022年9期)2022-08-16
Journal of Palaeogeography(2022年1期)2022-03-25
快乐语文(2021年35期)2022-01-18
林业科技(2020年3期)2021-01-21
今日农业(2020年19期)2020-12-14
农民致富之友(2019年4期)2019-03-13
摄影之友(影像视觉)(2017年1期)2017-07-18
制导与引信(2016年3期)2016-03-20
中国科技信息(2015年2期)2015-11-16
植物营养与肥料学报(2014年1期)2014-03-11
