
棉花纤维发育相关基因GhPRP10的克隆及其功能分析
2015-02-28 02:12李艳军刘永昌张新宇
西北植物学报 2015年4期
石 淼,李艳军,刘永昌,张新宇,薛 飞*,孙 杰
(1 石河子大学 生命科学学院,新疆石河子 832003;2 石河子大学 农学院/新疆兵团绿洲生态农业重点实验室,新疆石河子 832003)
棉花纤维发育相关基因GhPRP10的克隆及其功能分析
石 淼1,李艳军2,刘永昌2,张新宇2,薛 飞2*,孙 杰2
(1 石河子大学 生命科学学院,新疆石河子 832003;2 石河子大学 农学院/新疆兵团绿洲生态农业重点实验室,新疆石河子 832003)
利用电子序列拼接结合RT-PCR技术,从12DPA(开花后天数)棉纤维中克隆到1个编码富含脯氨酸蛋白(PRPs)基因,命名为GhPRP10(登录号KP036633)。GhPRP10基因开放阅读框为684 bp,编码228个氨基酸,其中脯氨酸(Pro)含量为34.6%。序列分析发现GhPRP10蛋白具有N端信号肽和富含脯氨酸区域,属于第一类PRPs。实时荧光定量PCR(RT-PCR)结果显示,GhPRP10在棉纤维组织中优势表达,在纤维发育过程中的表达量呈现先升高后降低的趋势,在18DPA纤维中表达量最高。利用Gateway技术构建植物过量表达载体,转入烟草BY-2悬浮细胞,表型观察和细胞长度测量结果显示,转GhPRP10基因细胞比野生型细胞显著增长。根据该基因的组织表达特征和转基因细胞表型分析,推测GhPRP10基因在纤维伸长和次生壁合成过程中发挥作用。
棉花;纤维;富含脯氨酸蛋白;BY-2烟草悬浮细胞
棉花是世界上最重要的经济作物之一,中国是世界上最大的棉花生产和消费国。棉纤维是棉花的主产品,其品质优劣直接决定纺织品的质量和效益。棉纤维是由胚珠表皮细胞发育而成的单细胞,其伸长率和最终长度远超过其它植物细胞,是研究植物细胞伸长和细胞壁生物合成的理想模型[1]。克隆棉纤维发育相关基因并解析其发育的分子调控机制,对改良棉纤维品质具有十分重要的理论意义[2]。
细胞壁是植物细胞特有的结构,它可以保护细胞并决定细胞的大小和形状,对细胞的生长发育具有重要作用[3]。植物细胞壁中除含有多糖外,还含有大量的蛋白成分。细胞壁结构蛋白主要分为4类[4]:富含脯氨酸蛋白(proline-rich Proteins,PRPs)、富含甘氨酸蛋白(glycine-rich proteins,GRPs)、伸展蛋白和阿拉伯半乳聚糖蛋白(arabinogalactan proteins,AGPs)。PRPs是一类富含脯氨酸和羟脯氨酸的细胞壁结构蛋白,在植物中广泛存在,前人已从大豆、小麦、玉米、水稻等植物中分离到多个编码该蛋白的基因[5-8],这些基因在逆境胁迫[9-12]和激素[13-14]响应的过程中发挥重要作用。此外,PRPs不仅能够决定细胞形态,而且能够调控植物的生长发育,AtPRP3[15]主要在拟南芥的根毛基部和正在伸长的根尖中表达,该基因突变后导致根毛形态异常、分枝发生变化,突变体植株的根毛显著减少,表明该基因参与根毛的形态建成。AtPRP4[16]在拟南芥的叶、茎、花、长角果和根形成的早期阶段表达,该基因突变体植株的侧根减少,叶片变小,种子生长受到抑制。AtPRPL1[17]仅在拟南芥的根和毛状体细胞中表达,将AtPRPL1基因敲除后使拟南芥根毛明显变短,该基因过量表达后能够促进根毛的伸长。
目前人们已从棉纤维细胞中分离出多个优势或特异表达的PRPs基因[18-22],例如GhPRP1、GhPRP2、GhPRP3、GhPRP5等基因。过量表达GhPRP5的转基因拟南芥生长迟缓,叶表皮细胞变小,植株矮小[23]。虽然该类基因在棉纤维中大量存在,但功能研究仍较少,因此研究这类蛋白在纤维发育过程中的功能对明确纤维发育机制至关重要。本研究从棉纤维中克隆到1个新的PRP基因,命名为GhPRP10,通过实时荧光定量PCR分析该基因在棉花不同组织中的表达情况,构建植物过量表达载体转化野生烟草BY-2悬浮细胞,对转基因烟草悬浮细胞的表型变化进行观察,为进一步阐明该基因在棉纤维细胞发育过程中的功能奠定基础。
1 材料和方法
1.1 植物材料
棉花(GossypiumhirsutumL.)品种‘新陆早33号’由石河子大学棉花研究所提供。将该材料播种于石河子大学农学院试验田,按常规进行农事作业和田间管理,在盛花期挂牌标记棉铃,按3、6、9、12、15、18、21、24、27 DPA(开花后天数)采集棉铃,室内剥取纤维,液氮速冻后保存于-80 ℃冰箱。
室内取‘新陆早33号’种子剥去种皮,将种仁浸泡于0.1% HgCl2溶液中消毒10 min,用无菌水反复冲洗3~5次,播种于1/2 MS固体培养基,28 ℃暗培养4 d,然后进行水培。待长出真叶后,取其根、茎、真叶,花在田间植株上采集,上述材料液氮速冻后保存于-80 ℃冰箱,用于总RNA提取。
野生型烟草(Nicotianatabacum)BY-2悬浮细胞用LS固体培养基每15 d继代1次进行保存。
1.2 方 法
1.2.1 棉花总RNA提取和第一链cDNA合成 棉花根、茎、叶、花和不同发育时期纤维的总RNA提取采用改良CTAB法[24],使用Reverse Transcriptase M-MLV反转录酶和引物Oligo dT(T18)将RNA反转录合成cDNA,具体步骤按照说明书进行。
1.2.2GhPRP10基因克隆 以棉花PRP相关EST序列BQ410667为探针序列,使用NCBI BLASTn对棉花EST序列进行比对,对同源性高的EST序列利用DNASTAR软件进行序列拼接,得到一个新的PRP相关基因序列,长度为864 bp。通过ORF finder预测其开放阅读框(ORF),设计扩增该ORF特异性引物ORF1-F和ORF1-R(表1),以12DPA棉纤维cDNA为模板,使用Primer STAR高保真酶进行PCR扩增,程序为98 ℃预变性3 min;98 ℃变性10 s,54 ℃退火15 s,72 ℃延伸1 min,30个循环,72 ℃延伸10 min。扩增产物通过1%琼脂糖凝胶电泳进行检测,回收PCR产物连接到载体pGEM-T easy上,热激转化大肠杆菌TOP10感受态细胞,挑取阳性克隆经过PCR和酶切双重验证,将质粒送至北京六合华大基因有限公司进行测序。

表1 PCR扩增引物
1.2.3 生物信息学分析 使用在线软件InterProScan(http://www.ebi.ac.uk/interpro/scan.html)进行蛋白质一级结构分析,预测GhPRP10的跨膜信号肽。用DNAMAN和Clustal X进行多重序列比对及同源性比对,采用MEGA6软件构建系统进化树。
1.2.4 实时荧光定量PCR 根据GhPRP10基因的cDNA序列设计引物PRPS-F和PRPS-R(表1),分别以棉花根、茎、叶、花和不同时期棉纤维为模板进行实时荧光定量PCR扩增,反应过程在罗氏Light Cycler 480仪器上进行。内参基因为UBI(GenBank序列号为AY117057),根据其序列设计引物UBI-F和UBI-R(表1),产物大小为210 bp。反应体系(10 μL)包含上、下游引物各0.2 μL,SYBR Premix ExTaq5 μL,cDNA模板1 μL,ddH2O 3.6 μL。实时荧光定量PCR反应参数:94 ℃预变性1 min,94 ℃变性15 s,55 ℃退火20 s,72 ℃延伸30 s,40个循环。实验设3次重复,结果按照2-△Ct法处理数据,并制作柱状图显示该基因在棉花不同组织中的表达情况。
1.2.5 植物过量表达载体构建及烟草BY-2悬浮细胞的转化 利用Gateway同源重组原理[25]构建GhPRP10植物过量表达载体pGWB17-GhPRP10,构建过程按照Invitrogen公司的pENTRTMDirectional TOPO®Cloning Kits和Gateway®LR ClonaseTMⅡ Enzyme Mix试剂盒进行。采用农杆菌电击转化法将质粒转入农杆菌LBA4404的菌株中。根据吴科瀛的方法将植物过表达载体转化烟草BY-2悬浮细胞[26],获得转基因抗性细胞团。挑取长势良好的抗性细胞团继代到新鲜筛选培养基中,每15 d继代1次,去除嵌合的野生细胞,继代4次后得到黄色纯合体转基因系。使用Trizol试剂盒提取总RNA以ORF1-F和ORF1-R为引物进行RT-PCR验证。取适量转基因细胞用含有抗生素的LS液体培养基摇培3~4 d后,用荧光素(FDA)进行染色后在荧光显微镜下连续6 d进行细胞形态观察拍照。使用Image J软件对野生型和3个转基因系细胞中分别选取的40个细胞的长度进行测量,数据统计分析使用SPSS 17.0软件。
2 结果与分析
2.1 GhPRP10基因克隆与序列分析
以12DPA棉纤维细胞cDNA为模板,利用引物ORF1-F和ORF1-R进行RT-PCR扩增拼接后的PRP基因ORF,获得该基因序列大小为687 bp(图1),将扩增序列与拼接序列进行比对,一致性达到100%。序列分析发现该基因共编码228个氨基酸,其中脯氨酸占34.6%,苏氨酸占13.2%,丝氨酸占13.2%,分子量为22 209.5 kD,等电点9.75。蛋白结构域分析发现(图2),该基因编码的蛋白属于第一类PRPs,第1~23个氨基酸残基为信号肽区域,有一个富含脯氨酸重复基序的区域,该区域含有15个分散的A(or S)T(or S)PPP基序,7个XPP基序和2个无规律的基序TQPPP和TSPPP,将该基因命名为GhPRP10。

图1 GhPRP10基因PCR扩增电泳结果M.Marker Ⅲ;1.阴性对照;2~6.PCR扩增产物

图2 GhPRP10基因的cDNA及氨基酸序列方框部分表示N端信号肽;双下划线表示A(S)T(S)PPP基序;虚线表示TQPPP和TSPPP;粗下划线表示XPP基序;星号表示终止密码子
2.2 GhPRP10系统进化分析
利用Clustal X2.1将GhPRP10氨基酸序列与其它植物中PRPs氨基酸序列进行同源比对(图3),图中显示GhPRP10与陆地棉GhPRP(AAA79360),亚洲棉Cotton_A_20973、Cotton_A_21704,雷蒙德氏棉Gorai_013G048900、Gorai_005G233500,海岛棉GbPRP(AAA79364),可可树TcUP(EOX98411)的一致性均较高。为进一步明确该蛋白与其它植物中富含脯氨酸蛋白之间的进化关系,在GenBank中选取已登录的不同植物的富含脯氨酸蛋白构建进化树(图4),结果显示GhPRP10是雷蒙德氏棉Go-rai013G048900、亚洲棉Cotton_A_20973基因的同源基因,GhPRP10与陆地棉GhPRP(AAA79360)亲缘关系较近,与其他PRPs基因的遗传距离相对较远。

图3 植物中不同PRPs蛋白的氨基酸序列比对
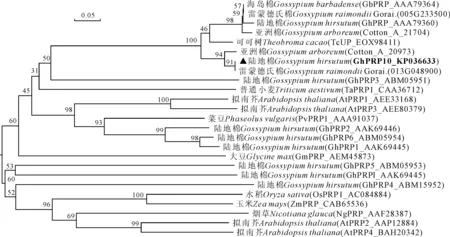
图4 棉花中GhPRP10与其它植物中PRPs的进化树分析图中分支点的数字表示Bootstrap验证中基于1 000次重复该节点可信度的百分比;标尺代表遗传距离;三角符号代表GhPRP10
2.3 GhPRP10基因的表达分析
利用Real time PCR技术分析GhPRP10基因在棉花不同组织中的表达情况,结果表明该基因在棉花根、叶和花中几乎不表达,在茎中微弱表达;在棉纤维中呈现优势表达,且受棉纤维发育调控,即在3~9DPA纤维细胞中几乎不表达,12~18DPA表达量持续增高,在18DPA达到最高,21~27DPA表达量有所降低,但仍维持较高的表达水平(图5)。由上述结果推测GhPRP10在棉纤维伸长及次生壁合成阶段发挥作用。
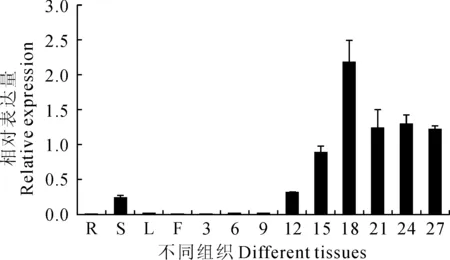
图5 GhPRP10在棉花不同组织中的表达情况R.根;S.茎;L.叶;F.花;3~27分别为3~27 DPA棉纤维细胞
2.4 转基因BY-2细胞的表型分析
将GhPRP10基因构建到植物表达载体上转化烟草BY-2细胞。RT-PCR结果显示8个BY-2细胞转基因系均扩增出约687 bp条带(图6),表明GhPRP10基因已经成功转入烟草BY-2细胞中。将摇培好的野生型细胞和转基因细胞分别进行染色后在荧光显微镜下连续观察6 d,发现野生型细胞在6 d中均表现为不规则圆形,细胞团由小迅速增大,增殖速度快(图7,W1~W6);而转基因细胞在6 d中均呈现不规则长形,分布较松散,细胞团增殖速度慢(图7,T1~T6)。对野生型和3个转基因细胞系的细胞长度进行测量,结果显示3个转基因细胞系(n=40)的平均细胞长度均显著长于野生型细胞,为野生型细胞的2倍以上(图8)。表明该基因的表达能够促进细胞伸长,推测其在棉纤维细胞伸长方面发挥作用。

图6 转基因BY-2细胞的RT-PCR验证M.Marker Ⅲ;1~8.8个转基因BY-2细胞系;9.野生型细胞;10.阳性对照

图7 野生型和转基因BY-2细胞荧光显微图W1~W6.1~6 d野生型细胞的观察图;T1~T6.1~6 d转基因细胞观察图
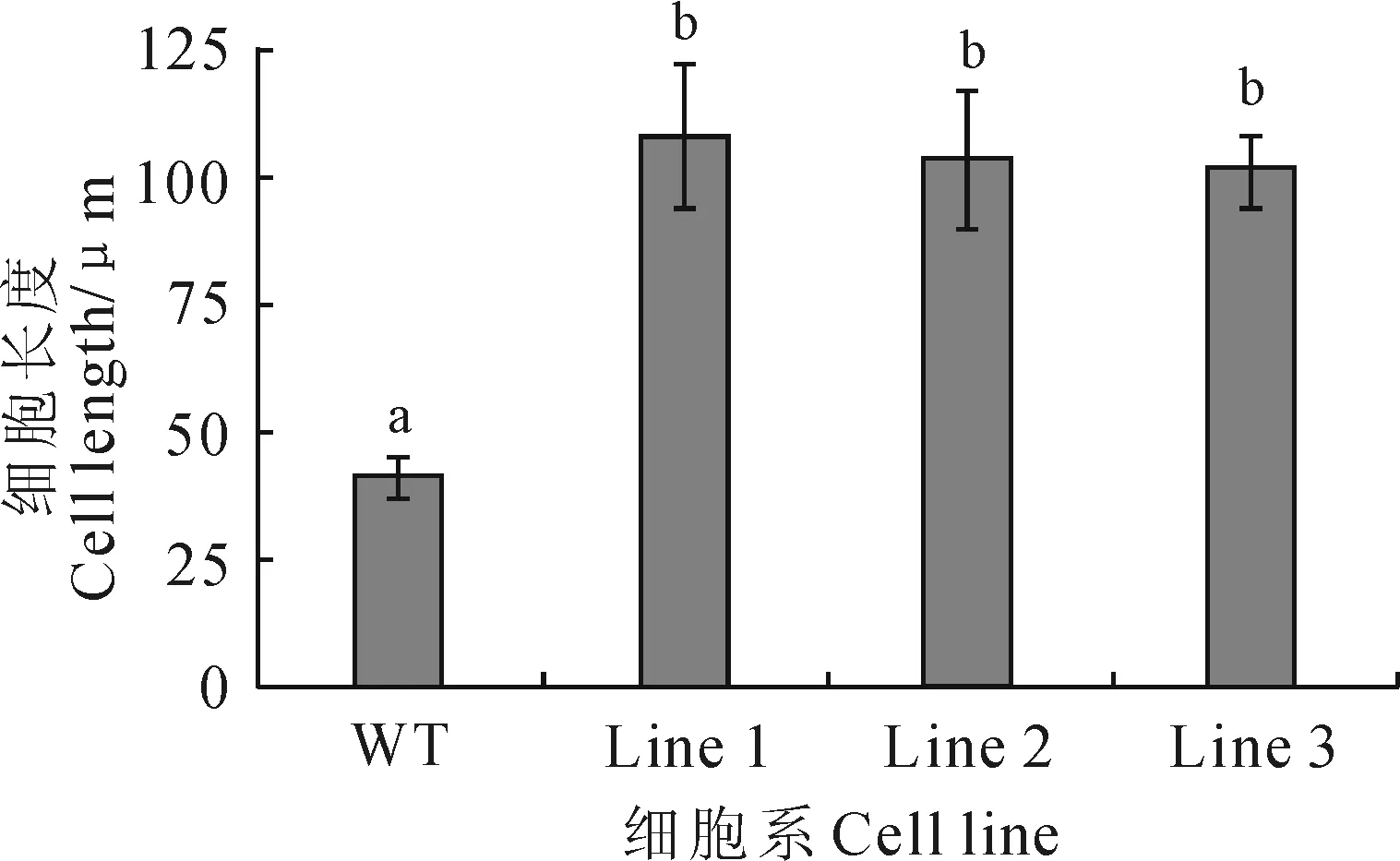
图8 野生型和转基因BY-2细胞长度比较WT.野生型细胞;1~3.转基因细胞系;标注字母表示0.05水平显著性
3 讨 论
PRPs是植物细胞壁结构蛋白[27],按结构可分为3类[11]:第一类由N端信号肽和富含脯氨酸重复基序区构成;第二类由N端信号肽、富含脯氨酸重复基序区和C端含有半胱氨酸区构成;第三类由N端亲水结构域和脯氨酸重复基序区构成。本研究从棉纤维中克隆了1个新的PRPs基因GhPRP10,蛋白质一级结构分析发现GhPRP10蛋白属于第一类PRPs,含有N端信号肽,该结构域可能起着将蛋白质锚定到细胞壁上的作用。进化树分析发现GhPRP10与其他已命名的陆地棉PRPs基因遗传距离相对较远,表明该基因是一个新的PRP基因。同源性分析发现GhPRP10在陆地棉供体祖先种雷蒙德氏棉、亚洲棉中的同源基因分别是Gorai013G-048900和Cotton_A_20973,说明GhPRP10在陆地棉中存在多个拷贝。
棉纤维发育分为4个时期,即起始期(0~5DPA)、伸长期(5~20DPA)、次生壁合成期(20~45DPA)和成熟期(45~50DPA)[28]。前人已在棉纤维中发现多个PRP基因[18-22],其中GhPRP3、GhPRP5在棉纤维细胞中特异表达,在伸长期表达量较高,且受纤维发育调控[19-20]。本研究中实时荧光定量PCR分析表明GhPRP10基因在棉花纤维中优势表达,其表达量呈先上升,至18DPA达到最高峰,之后降低的趋势,但在21~27DPA表达量仍维持在较高水平。GhPRP10高表达时期处于棉纤维细胞伸长与次生壁合成期,推测GhPRP10不仅参与棉纤维伸长,且在次生壁的建成过程中发挥重要作用。
已有研究表明,富含脯氨酸蛋白涉及细胞的生长发育以及应对外界环境刺激等多种功能[3-4]。烟草NtProRP1是一个渗透胁迫响应因子,对花粉管伸长及在种胚发育的早期有重要作用[29]。HyPRP1在辣椒和烟草中对细胞坏死具有正调控作用,对病原菌的防御具有负调控作用[30]。关于棉花PRPs基因的功能研究很少,近期许文亮等发现在棉花中干扰GhPRP5表达后能使棉纤维变长,说明该基因在棉纤维发育过程中为负调控因子。同时,GhPRP5还参与调控拟南芥胁迫相关基因的表达[23]。本研究通过将GhPRP10基因转化烟草BY-2细胞,发现该基因过量表达使BY-2细胞长度显著增长,且细胞的形态发生了明显的改变,由圆形细胞变为长形细胞,这与该基因在棉纤维的伸长期和次生壁增厚期优势表达相一致。综上分析,GhPRP10基因能够促进细胞伸长,推测它能调控棉纤维的发育。Tábata等[31]发现快速碱化多肽因子AtRALF1通过调控AtPRP1、AtPRP3(富含脯氨酸的蛋白)、AtHRPG2(富含羟脯氨酸的糖蛋白),以及TCH4(木葡聚糖)来阻止油菜素类固醇的合成,从而影响根细胞的伸长和侧根的形成。GhPRP10能够促进烟草悬浮细胞的伸长,但如何调控棉纤维细胞的发育还有待进一步研究。
[1] KIM H J,TRIPLETT B A.Cotton fiber growth in plant andinvitro:models for plant cell elongation and cell wall biogenesis[J].PlantPhysiology,2001,127(4):1 361-1 366.
[2] GUO W ZH(郭旺珍),SUN J(孙 敬),ZHANG T ZH(张天真).Cloning and molecular breeding of the fiber quality gene[J].ChineseScienceBulletin(科学通报).2003,48(5):410-417(in Chinese).
[3] KEEGSTRA K.Plant cell walls[J].PlantPhysiology,2010,154(2):483-486.
[4] SHOWALTER A M.Structure and function of plant cell wall proteins[J].ThePlantCell,1993,5(1):9-23.
[5] ZHAI Y(翟 莹),LEI T T(雷婷婷),LI J W(李景文),etal.Cloning and expression of a stress-inducedGmPRPgene in soybean (Glycinemax)[J].ActaAgronomicaSinica(作物学报),2011,37(12):2 152-2 157(in Chinese).
[6] 赵维萍.普通小麦中一类富含脯氨酸蛋白基因的克隆与功能分析[D].南京:南京农业大学,2011.
[7] VIGNOLS F,JOSE-ESTANYOL M,CAPARROS-RUIZ D,etal.Involvement of a maize proline-rich protein in secondary cell wall formation as deduced from its specific mRNA localization[J].PlantMolecularBiology,1999,39(5):945-952.
[8] WANG R,CHONG K,WANG T.Divergence in spatial expression patterns and in response to stimuli of tandem-repeat paralogues encoding a novel class of proline-rich proteins inOryzasativa[J].JournalofExperimentalBotany,2006,57(11):2 887-2 897.
[9] PRIYANKA B,SEKHAR K,REDDY V D,etal.Expression of pigeonpea hybrid-proline-rich protein encoding gene (CcHyPRP) in yeast andArabidopsisaffords multiple abiotic stress tolerance[J].PlantBiotechnologyJournal,2010,8(1):76-87.
[10] YI Z,MICHAEL SCHLPPI.Cold responsiveEARLI1 typeHyPRPsimprove freezing survival of yeast cells and form higher order complexes in plants[J].Planta,2007,227(1):233-243.
[11] HE C Y,ZHANG J S,CHEN S Y.A soybean gene encoding a proline-rich protein is regulated by salicylic acid,an endogenous circadian rhythm and by various stresses[J].TheoreticalandAppliedGenetics,2002,104(6-7):1 125-1 131.
[12] QIN L X,ZHANG D J,LI X B,etal.CottonGhHyPRP3 encoding a hybrid proline-rich protein is stress inducible and its overexpression inArabidopsisenhances germination under cold temperature and high salinity stress conditions[J].ActaPhysiologiaePlantarum,2013,35(5):1 531-1 542.
[13] ZHANG D J(张德静),QIN L X(秦丽霞),LI L(李 龙),etal.Expression of cottonGhPRP5 gene inArabidopsisenhances susceptibility to ABA and salt stresses[J].ActaAgronomicaSinica(作物学报),2013,39(3):563-569(in Chinese).
[14] CHAI Q X(柴秋霞),LI B CH(李本昌),XU Z Q(徐子勤).Subcellular localization and resistance toGibberellafujikuroiof AtELHYPRP2 in transgenic tobacco[J].ChineseJournalofBiotechnology(生物工程学报),2014,30(3):472-484(in Chinese).
[15] BERNHARDT C,TIERNEY M L.Expression ofAtPRP3,a proline-rich structural cell wall protein fromArabidopsis,is regulated by cell-type-specific developmental pathways involved in root hair formation[J].PlantPhysiology,2000,122(3):705-714.
[16] RAAB S,HOTH S.A Mutation in theAtPRP4 splicing factor gene suppresses seed development inArabidopsis[J].PlantPhysiology,2007,9(3):447-452.
[17] BORON A K,VAN ORDEN J,NEKTARIOS MARKAKIS M,etal.Proline-rich protein-likePRPL1 controls elongation of root hairs inArabidopsisthaliana[J].JournalofExperimentalBotany,2014,65(18):5 485-5 495.
[18] JOHN M E,KELLER G.Characterization of mRNA for a proline-rich protein of cotton fiber[J].PlantPhysiology,1995,108(2):669-676.
[19] TAN H,CREECH R G,JENKINS J N,etal.Cloning and expression analysis of two cotton (GossypiumhirsutumL.) genes encoding cell wall proline-rich proteins[J].DNASequence,2001,12(5-6):367-380.
[20] XU W L(许文亮),HUANG G Q(黄耿青),LI X B(李学宝),etal.Molecular characterization and expression analysis of five novel genes encoding proline-rich proteins in cotton[J].ProgressinBiochemistryandBiophysics(生物化学与生物物理进展),2007,34(5):509-517(in Chinese).
[21] LI X B(李学宝),HUANG G Q(黄耿青),XU W L(许文亮),etal.Solation of the cotton genes that encoded cell wall proteins and their expression profile in cotton fibers[J].JournalofCentralChinaNormalUniversity(华中师范大学学报),2005,39(4):509-513(in Chinese).
[22] FENG J X,JI S J,SHI Y H,etal.Analysis of five differentially expressed gene families in fast elongating cotton fiber[J].ActaBiochimicaandBiophysica,2004,36(1):51-56.
[23] XU W L,ZHANG D J,LI X B,etal.Cotton PRP5 gene encoding a proline-rich protein is involved in fiber development[J].PlantMolecularBiology,2013,82:353-365.
[24] JIANG J X(蒋建雄),ZHANG T ZH(张天真).Extraction of total RNA in cotton tissues with CTAB-acidic phenolic method[J].CottonScience(棉花学报),2003,15(3):166-167(in Chinese).
[25] KARIMI M,INZÉ D,DEPICKER A.GATEWAY vectors for agrobacterium-mediated plant transformation[J].TrendsinPlantScience,2002,7(5):193-195.
[26] WU K Y(吴科瀛),LIANG Y L(梁玉玲).Study on tobacco BY-2 cell culture and salt stress[J].AnimalHusbandryandFeedScience(畜牧与饲料科学),2009,30(4):8-9(in Chinese).
[27] FRANCISCO S M,TIERNEY M L.Isolation and characterization of a proline-rich cell wall protein from soybean seedlings[J].PlantPhysiology,1990,94(4):1 897-1 902.
[28] DU X M(杜雄明),PAN J J(潘家驹),WANG R H(汪若海).Differentiation and development of fiber cells on the ovules in cotton[J].CottonScience(棉花学报),2000,12(4):212-217(in Chinese).
[29] CHEN J,ZHAO J,NING J,etal.NtProRP1,a novel proline-rich protein,is an osmotic stress-responsive factor and specifically functions in pollen tube growth and early embryogenesis inNicotianatabacum[J].PlantCellandEnvironment,2014,37(2):499-511.
[30] YEOM S I,SEO E,OH S K,etal.A common plant cell-wall proteinHyPRP1 has dual roles as a positive regulator of cell death and a negative regulator of basal defense against pathogens[J].ThePlantJournal,2012,69(5):755-768.
(编辑:宋亚珍)
Cloning and Functional Analysis ofGhPRP10 Relatedto Fiber Development inGossypiumhirsutum
SHI Miao1,LI Yanjun2,LIU Yongchang2,ZHANG Xinyu2,XUE Fei2*,SUN Jie2
(1 College of Life Science,Shihezi University,Shihezi,Xinjiang 832003,China;2 College of Agronomy,Shihezi University/ Key Oasis Eco-agriculture Laboratory of Production and Construction Group,Shihezi,Xinjiang 832003,China)
In this study,a new gene encoding proline-rich protein was isolated from 12 days post anthesis(DPA) cotton fiber,and designated asGhPRP10.The ORF of the gene was 684bp encoding 228 amino acids,contained 34.6% proline.The bioinformatics analysis showed that the protein encoded by this gene belonged to the first class of PRPs,which has a signal peptide in N terminal and a proline rich conserved region.The quantitative RT-PCR results showed thatGhPRP10 is preferentially expressed in cotton fiber.The transcript level increased along with fiber development and reached the highest abundance at 18DPA,after which time it gradually decreased.The plant over-expression vector was constructed using Gateway technology and then transformed into tobacco suspension BY-2 cells.The phenotype observation and cell length measurement found that the transgenic cells were significantly longer than that of the wild type.Based on the expression profile ofGhPRP10 and phenotype analysis of transgenic cells,it is presumed that the gene plays an important role in the process of fiber elongation and secondary wall synthesis.
cotton;fiber;proline-rich protein;tobacco suspension BY-2 cells
1000-4025(2015)04-0639-07
10.7606/j.issn.1000-4025.2015.04.0639
2014-11-26;修改稿收到日期:2015-03-05
国家自然科学基金(U1128301);十二五科技支撑计划项目(2011BAD35B05);石河子大学育种专项(gxjs2012-yz01,gxjs2012-yz04)
石 淼(1989-),女,在读硕士研究生,主要从事棉花分子育种研究。E-mail:shim0501@163.com
*通信作者:薛 飞,副教授,博士,主要从事棉花分子育种研究。E-mail:xuefei2011@gmail.com
Q785;Q786
A
猜你喜欢
中老年保健(2022年3期)2022-11-21
中国纤检(2022年8期)2022-09-22
纺织标准与质量(2022年4期)2022-09-05
南方医科大学学报(2022年3期)2022-04-13
棉纺织技术(2021年4期)2021-07-14
浙江大学学报(农业与生命科学版)(2021年3期)2021-07-10
中国预防兽医学报(2020年8期)2020-11-05
三农资讯半月报(2020年15期)2020-08-25
棉花学报(2020年3期)2020-08-08
科学与财富(2019年15期)2019-10-21
