
棉花HSP70基因的克隆与原核表达
2015-02-28 01:58曲延英陈全家倪志勇
西北植物学报 2015年4期
刘 娜,曲延英,陈全家,倪志勇
(新疆农业大学 农学院/新疆农业大学农业生物技术重点实验室,乌鲁木齐 830052)
棉花HSP70基因的克隆与原核表达
刘 娜,曲延英,陈全家,倪志勇*
(新疆农业大学 农学院/新疆农业大学农业生物技术重点实验室,乌鲁木齐 830052)
以抗旱性较强的棉花品种‘KK1543’为材料,采用RT-PCR技术克隆了1个棉花HSP70基因,命名为GhHSP70。研究结果表明:GhHSP70基因开放阅读框长1 997 bp,编码648个氨基酸,GhHSP70蛋白相对分子量为 70.94 kD,等电点为4.83。氨基酸序列比对和系统进化树分析发现,GhHSP70与欧洲大叶杨HSP70的亲缘关系最近,氨基酸序列一致性达到96.4%。为进一步分析基因的功能,构建原核表达载体pGEX-4T-1-GhHSP70,并在大肠杆菌中异源表达。SDS-PAGE分析表明所表达蛋白与预期蛋白大小一致,重组蛋白在37 ℃,0.2 mmol/L IPTG诱导2 h时表达量最大。研究为进一步研究棉花HSP70基因功能奠定基础。
棉花;热激蛋白70;基因克隆;原核表达
干旱是世界上危害最严重的一种自然灾害,是一个长期存在的世界性难题。目前世界上有三分之一以上的土地处于干旱和半干旱地带,其他地区在植物生长季节也常发生不同程度的干旱[1],干旱影响植物各个阶段的生长发育和生理代谢过程。棉花是关系国计民生的重要物资,是仅次于粮食作物的第二大农作物,其产值占中国经济作物的50%以上,在国民经济发展中具有重要地位[2]。干旱严重影响棉花的产量和品质,已成为中国棉花生产发展的重要限制因素。因此提高棉花的抗旱能力已经成为现代棉花研究工作中急需解决的关键问题之一。
热激蛋白(HSPs,heat shock proteins)是细胞受到不利环境刺激时被激活并表达增强的一类蛋白[3-4]。该类蛋白根据分子量的大小主要分为4类:Hsp90家族(83~90 kD)、Hsp70家族(66~78 kD)、Hsp60家族及小Hsp家族。其中HSP70是HSP家族中最重要、研究最深入的一种,是重要的分子伴侣[5-8],它能防止蛋白变性,并辅助蛋白质重折叠为正常构象,或者将受到不可恢复伤害的蛋白通过蛋白水解途径排除到细胞外,从而有利于生物体抵抗胁迫压力[9]。HSP70对植物抵抗干旱、高温、寒冷、氧化等胁迫乃至对植物的生长发育均起到十分重要的作用[10-13],是育种工作中不可多得的优良基因。本研究前期采用蛋白质组双向电泳技术和生物信息学方法,对抗旱性较强的棉花品种‘KK1543’进行研究,获得了113个在干旱胁迫下差异表达的蛋白。质谱和生物信息学分析发现蛋白质点1312为棉花HSP70蛋白,这说明棉花HSP70蛋白可能在干旱胁迫应答中具有功能[14]。
本研究根据获得的HSP70蛋白的氨基酸序列设计引物,以棉花品种‘KK1543’为材料,克隆棉花HSP70基因,对其进行序列分析,同时构建了基因的原核表达载体并在大肠杆菌中原核表达,为进一步研究棉花HSP70蛋白的功能奠定基础。
1 材料和方法
1.1 研究材料
棉花品种‘KK1543’由新疆农业大学农学院作物遗传育种实验室提供。在实验室已有的60份抗旱材料中,进行抗旱性筛选,发现‘KK1543’抗旱性较强。该品种不但具有抗旱性较好的特点、而且籽大,适用于水培条件下研究棉花的抗旱性。
1.2 试验方法
1.2.1 RNA提取及cDNA第一链合成 利用Plus植物总RNA提取试剂盒(天根试剂公司)提取棉花叶片总RNA。按照Thermo反转录试剂盒(Thermo试剂公司)说明书操作步骤,以获得的棉花叶片总RNA为模板,合成cDNA第一条链。
1.2.2GhHSP70基因的扩增 根据前期质谱分析获得的棉花HSP70基因的氨基酸序列设计特异性引物GhHSP70-F(5′-AGAGTGATGGCCGGAAA-AGGAG-3′)和GhHSP70-R(5′-AGCTGTTAAACATCGGCAAAATACTATCT-3′),以棉花叶片cDNA第一链为模板,扩增基因。扩增体系:10×缓冲液(含Mg2+)5 μL,2.5 mmol/L dNTP 4 μL,10 μmol/L引物各2 μL,cDNA 1 μL,Taq酶(5 U/μL) 0.5 μL,ddH2O 35.5 μL 。反应程序:预变性94 ℃ 4 min,变性94 ℃ 30 s,退火60 ℃ 30 s,延伸72 ℃ 1 min 30 s,35个循环,再延伸72 ℃ 10 min,结束反应。经1%琼脂糖凝胶电泳检测扩增产物,采用普通DNA产物纯化试剂盒(天根试剂公司)纯化PCR产物,通过T-A克隆法连接到pMD19-T(Takara试剂公司),转化大肠杆菌DH5α感受态细胞(宝信试剂公司),涂布于含有氨苄青霉素(50 mg/mL)的LB固体培养基上,37 ℃过夜培养后,挑取白色单菌落,经含有氨苄青霉素的液体培养基震荡培养12 h后,菌液PCR检测是否有插入片段,将阳性克隆送往北京鼎国昌盛生物技术有限公司测序。
1.2.3 生物信息学分析 通过检索GenBank和利用DNAMAN等软件进行同源性和多重序列比较分析。用 MEG 4.0软件构建系统进化树,利用DNAStar的protean软件预测二级结构,利用在线软件(http://www.cbs.dtu.dk/services/TMHMM/)分析棉花热激蛋白70基因编码的氨基酸序列的跨膜结构。
1.2.4GhHSP70基因原核表达载体的构建 根据原核表达载体pGEX-4T-1的多克隆位点,设计带有酶切位点引物70-28-F(5′-TTAGGATCCATGGCCGGAAAAGGAGAAG-3′)和70-28-R(5′-TAAG-CGGCCGCTTAGTCGACTTCTTCTATCTTAGG-T-3′),并以pMD19-T-GhHSP70质粒为模板,按上述体系和程序进行PCR扩增,经1%琼脂糖凝胶电泳检测扩增产物。利用NotⅠ和BamHⅠ(Thermo试剂公司)分别双酶切PCR纯化产物与空质粒pGEX-4T-1,酶切产物纯化后,用T4DNA连接酶22 ℃定向连接20 min,将连接产物转化大肠杆菌DH5α,通过菌液PCR初步筛选阳性重组子,阳性克隆送上海美季测序公司测序。
1.2.5GhHSP70基因的原核表达 使用热激法将重组质粒转化大肠杆菌Rosetta(DE3)感受态细胞(北京全式金),涂布于含有氨苄青霉素(50 mg/mL)的LB固体培养基上,37 ℃过夜培养后挑取单菌落,经含有氨苄青霉素的液体培养基震荡培养12 h后,通过菌液PCR筛选阳性克隆。
将重组菌株接种于2 mL含50 mg/L氨苄青霉素的LB液体培养基中,37 ℃、200 r/min过夜培养。然后按1∶50比例转接到10 mL新鲜LB液体培养基中(含50 mg/L氨苄青霉素),225 r/min培养3 h(OD600≈0.6),加入IPTG至终浓度分别为0、0.2、0.5、1.0、1.5和2.0 mmol/L,37 ℃诱导4 h后收集菌液。以0.5 mmol/L IPTG诱导4 h的pGEX-4T-1空载体作为对照。8 000 r/min,离心10 min,弃上清,菌体样品加入30 μL 2×上样缓冲液和50 μL ddH2O,混匀,在100 ℃的沸水中煮沸10 min,冰上冷却后,取10 μL进行 SDS-PAGE(5%浓缩胶,12%分离胶)电泳检测。电泳后经考马斯亮兰染色、拍照,分析蛋白表达结果,确定最适IPTG诱导浓度。
按上述方法,以终浓度为1 mmol/L IPTG进行重组菌株表达蛋白的诱导表达,以相同条件的pGEX-4T-1转化菌为对照。37 ℃培养0、2、4和6 h后分别收集菌液,进行SDS-PAGE(5%浓缩胶,12%分离胶)电泳检测,分析蛋白表达结果,确定最适培养时间。
2 结果与分析
2.1 GhHSP70基因克隆与序列分析
以棉花叶片cDNA为模板,利用引物GhHSP70-F和GhHSP70-R,通过PCR扩增获得一条大约2 kb左右的特异条带(图1)。通过T-A克隆将扩增产物克隆到pMD19-T载体上,获得重组质粒pMD19-T-HSP70,测序结果表明该基因ORF为1 997 bp,5′末端长为170 bp,3′末端长202 bp编码含648个氨基酸残基蛋白;利用DNAMAN软件预测的蛋白质的理论分子量为 70.94 kD,等电点为4.83。
同源性比对分析表明,棉花HSP70具有3个典型的HSP70签名基序序列(IDLGTTYS、IFDLGGGTFDVSLL、VVLVGGSTRIPKVQQ),其在氨基酸序列中所处的位置基本上相同,C-端特征基序均为EEVD,所得氨基酸序列在NCBI中进行Blast分析表明,第9个氨基酸到第618个氨基酸为该序列的结构域(图4)。GhHSP70编码的氨基酸
序列与已报道其他植物的HSP70序列都具有高度的同源性,其中与欧洲大叶杨HSP70的氨基酸水平一致性达到96.4%,表明HSP70进化的保守性。
将棉花HSP70的氨基酸序列与其他已报道植物的HSP70进行多序列比对,利用CLUSTALW和MEGA 4.0软件构建系统进化树(图2)。从图3可以看出,棉花(Gossypiumhirsutum)HSP70与欧洲大叶杨(Populustrichocarpa)HSP70为同一分支,说明棉花与欧洲大叶杨在进化上亲缘关系较近。
利用DNAStar软件的protean程序对棉花HSP70氨基酸序列进行二级结构预测(图3)。结果表明,该蛋白的二级结构由α-螺旋、 β-折叠、转角和无规则卷曲组成。利用在线软件http://www.cbs.dtu.dk/services/TMHMM/对棉花HSP70蛋白的跨膜区进行预测,表明棉花HSP70蛋白无跨膜区,预测结果如下(图5)。
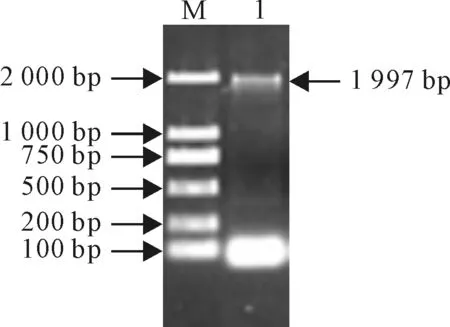
图1 RT-PCR产物电泳图1.RT-PCR产物;M.DL2000

图2 系统进化树

图3 GhHSP70二级结构预测标尺表示蛋白序列区段

图4 棉花与其他植物的HSP70的氨基酸序列比对结果划线部分为GhHSP70结构域,阴影部分表示同源氨基酸残基、签名序列和胞质HSP70 C-末端特征基序标出;CsHSP70.黄瓜(XP_004142749.1);JcHSP70.桐油树(KDP44981.1);MdHSP70.苹果(XP_008370710.1);MnHSP70.川桑(EXB58128.1);NtHSP70.烟草(AAR17080.1);ObHSP70.短花稻(XP_006663128.1);OSHSP70.水稻(NP_001068540.1);PtHSP70.欧洲大叶杨(XP_002311161.1);TcHSP70.可可(XP_007009362.1);VvHSP70.葡萄(XP_002284008.1);GhHSP70.棉花(ACJ11742)
2.2 GhHSP70基因原核表达载体构建与表达分析
对重组质粒pGEX-4T-1-GhHSP70进行菌液PCR检测,电泳结果见图6,在1 997 bp位置处有1条目的条带与预期结果一致,同时,该重组载体插入DNA片段的测序结果与棉花HSP70基因序列完全相同,表明棉花HSP70基因的原核表达载体构建成功。

图5 TMHMM工具预测结果图
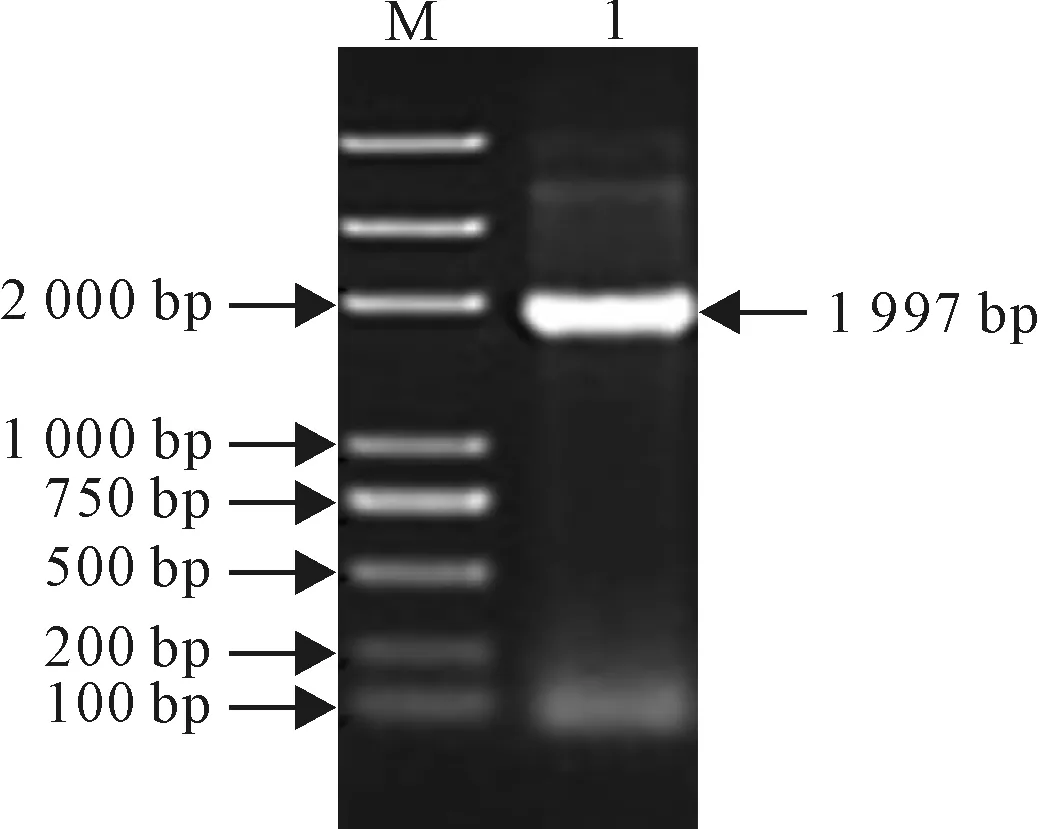
图6 pGEX-4T-1-GhHSP70菌液PCR

图7 IPTG终浓度对 HSP70蛋白表达的影响M.蛋白质标准分子量;1.pGEX-4T-1空载体未诱导;2.pGEX-4T-1空载体经0.5 mmol/L IPTG诱导4 h;3.pGEX-4T-1-GhHSP70未诱导;4~8.pGEX-4T-1-GhHSP70分别经终浓度为0.2、0.5、1.0、1.5和2.0 mmol/L的IPTG诱导4 h
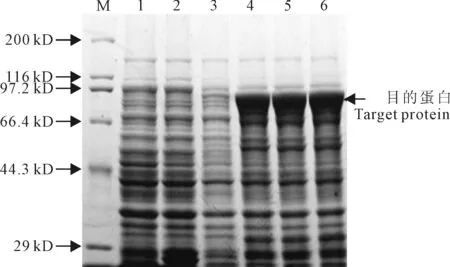
图8 IPTG诱导培养时间对HSP70蛋白表达的影响M.蛋白质标准分子量;1.pGEX-4T-1空载体未诱导;2.pGEX-4T-1空载体经0.5 mmol/L IPTG诱导4 h;3.pGEX-4T-1-GhHSP70未诱导;4~6.pGEX-4T-1-GhHSP70经0.5 mmol/L IPTG分别诱导2、4和6 h
在37 ℃下,在终浓度为 0.2、0.5、1.0、1.5和2.0 mmol/L的IPTG与不加IPTG下,分别对含重组质粒pGEX-4T-1-GhHSP70的大肠杆菌进行表达诱导4 h,对pGEX-4T-1空载体在0.5 mmol/L IPTG诱导表达4 h,其表达产物的SDS-PAGE电泳检测结果(图7)显示,在5个不同浓度的IPTG诱导下,均能诱导出融合蛋白,且所表达蛋白与预期蛋白大小一致,而pGEX-4T-1空载体则没有融合蛋白表达,表明0.2 mmol/L IPTG诱导就获得最佳的蛋白诱导效果。
在37 ℃下,用IPTG(0.5 mmol/L)诱导棉花HSP70重组蛋白表达,分别诱导0、2、4和6 h。发现在诱导时间为2、4和6 h,均能诱导出融合蛋白,其中诱导时间为2 h时重组蛋白表达量就已达到最大值。而经0.5 mmol/L IPTG诱导4 h的pGEX-4T-1空载体则没有融合蛋白表达(图8)。
3 讨 论
HSP70是研究最多、在生物体内分布最广、进化上最保守的一类热激蛋白,在植物受到高温、干旱、低温、高盐等非生物胁迫时可迅速并短时表达,以减少植物细胞受到的伤害。长期以来一直受到人们重视。而在各种非生物胁迫中,干旱对棉花产量的影响占首位,可以导致棉花细胞脱水、膜系统和酶系统遭到破坏、细胞功能丧失等[15]。因此,运用基因工程手段发掘和克隆抗旱基因,分离出与棉花抗旱品质紧密相关的基因显得至关重要,对于今后培育抗旱棉花新品系具有重大意义。
本研究则根据前期采用蛋白质组双向电泳技术和生物信息学方法,对抗旱性较强的棉花品种‘KK1543’进行研究,获得了113个在干旱胁迫下差异表达的蛋白。发现蛋白质点1312为棉花HSP70蛋白,推测该蛋白可能在干旱胁迫应答中具有功能。因此以获得的差异蛋白HSP70的氨基酸序列为基础,利用RT-PCR技术成功克隆出棉花HSP70基因,系统进化树分析,发现棉花HSP70与已报道其他植物的HSP70均有较高的同源性,其中与欧洲大叶杨HSP70具有的氨基酸水平一致性最高,蛋白亲缘关系最近。棉花HSP70氨基酸序列存在真核细胞HSP70家族的三个特征标签以及真核细胞特征基序,含有四肽重复序列GGMP和ATP结合位点的特征性基序DLGTT-S-V(13-21)。 具有细胞质定位的HSP70 C-末端特征基序EEVD。
外源基因在大肠杆菌中能否高效表达受很多因素的影响,如大肠杆菌菌株、诱导剂浓度、温度和诱导时间等[16]。有报道指出,0.1~1 mmol/L范围内的IPTG均适于重组蛋白诱导表达[17],而低浓度的IPTG可以减少化学物质对细胞的损伤[18]。在预实验基础上本实验将IPTG浓度确定为0.2 mmol/L,在37 ℃下诱导2 h时重组蛋白表达量就已达到最大值,实验结果证明该浓度诱导棉花HSP70重组蛋白表达是可行的。并为下一步纯化GhHSP70蛋白以及研究其功能提供实验基础。GhHSP70基因在大肠杆菌中表达的实验结果,为以后进一步将HSP基因应用到棉花抗旱的实际育种工作中提供了有利的理论基础。
[1] LI H CH(黎祜琛),YIN ZH J(印治军).A review of studies of drought resistance in tree species and drought resistant forestation technology[J].WorldForestryResearch(世界林业研究),2003,16(4):17-22(in Chinese).
[2] YANG ZH N(杨忠娜),TANG J J(唐继军),YU X L(喻晓玲).Xinjiang cotton industry present situation and countermeasure research[J].ResearchofAgriculturalModernization(农业现代化研究),2013,34(3):298-302(in Chinese).
[3] RITOSSA F.A new puffing pattern induced by temperature shock and DNP inDrosophila[J].Experientia(Basel),1962,18:571-573.
[4] TISSIERES A,MITCHELL H K,TRACY U M.Protein synthesis in salivary glands ofDrosophilamelanogaster-relation to chromosome puffs[J].J.Mol.Biol.,1974,84:389-398.
[5] FRIZT S,RALF P,ANDREAS R.Regulation of the heat-shock response[J].PlantPhysiol.,1998,117(4):1 135-1 141.
[6] PHILIP J,CHRISTINE P,etal.Functional specificity among Hsp70 molecular chaperones[J].Science,1997,275(17):387-389.
[7] WANG X W,BASIA V,ODED S,etal.Role of plant heat-shock proteins and molecular chaperones in the abiotic stress response[J].TrendsinPlantScience,2004,19(5):244 -252.
[8] JAKOB U,GAESTAL M,etal.Small heat shock protein are molecular chaperones[J].BiologicalChemistry,1993,268(3):1 517-1 520.
[9] KIANG JG,TSOKOS G C.Heat shock protein 70 kDa:molecular biology,biochemistry and physiology[J].Pharmacol.Ther.,1998,80(2):183-201.
[10] HUANG X F(黄祥富),HUANG SH ZH(黄上志),FU J R(傅家瑞).Functional and expression of plant heat shock protein[J].ChineseBulletinofBotany(植物学通报),1999,16(5):530-536(in Chinese).
[11] NATAALIE B,CHRISTIANE R,YVES D,etal.Accumulation of small heat shock proteins,including mitochondrial HSP22,induce by oxidative stress and adaptive response in tomato cells[J].ThePlantJournal,1998,13(4):519-527.
[12] LEE B H,WON S H,LEE H S.Expression of the chloroplast-localized small heat shock protein by oxidative stress in rice[J].Gene,2000,245(2):283-290.
[13] DEROCHER A,VIERLING E.CytoplasmicHSP70 homologues of pea:differential expression in vegetative and embryonic organs[J].PlantMolecularBiology,1995,27(3):441-456.
[14] 郭忠军.干旱胁迫下棉花蛋白质组双向电泳体系构建与差异表达蛋白功能分析[D].乌鲁木齐:新疆农业大学,2012.
[15] LIU J D(刘金定),YE W W(叶武威).Cotton resistant research and utilization in our country[J].ChinaCotton(中国棉花),1998,25(3):5-6(in Chinese).
[16] LI Y(李 琰),ZHANG Q(张 谦),LI H X(李海霞),etal.Prokaryotic ex-pression analysis of a NBS-typePtDRG01 gene isolated fromPopulustomentosaCarr[J].ActaBot.Boreal.-Occident.Sin.(西北植物学报),2008,28(5):882-888(in Chinese).
[17] WU R(吴 任),XIE SH T(谢数涛),SUN Y(孙 勇),etal.High level prokaryotic expression of heat shock protein 70 inLitopenaeusvannamei[J].JournalofFisherySciencesofChina(中国水产科学),2006,3(2):305-309(in Chinese).
[18] 里斯.基因与基因组分析[M].北京:北京大学医学出版社,2006:352.
(编辑:宋亚珍)
Cloning and Prokaryotic Expression of Heat ShockProtein 70 Gene inGossypiumhirsutumL.
LIU Na,QU Yanying,CHEN Quanjia,NI Zhiyong*
(College of Agronomy,Xinjiang Agricultural University,Key Laboratory of Agricultural Biological Technology,Xinjiang Agricultural University,Urumqi 830052,China)
In this study,the strong drought-resistant cotton variety ‘KK1543’ was used as the material.RT-PCR method was used to clone aHSP70 gene,a gene coding forGhHSP70 was isolated from cotton (GossypiumhirsutumL.).The open reading frame was 1 997 bp,encoding 648 amino acid residues with a predicted molecular mass of 70.94 kDa and a basic isoelectric point of 4.83.Homology analysis and phylogenetic tree analysis founded that GhHSP70 shared 96.4% amino acid identity with the HSP70 fromPopulustrichocarpa,and GhHSP70 protein is close to PtHSP70.An expression vector pGEX-4T-1-GhHSP70 was constructed.Then,the recombinant plasmid was transformed intoEscherichiacoli.The SDS-PAGE results displayed that the expressed protein was consistent with the anticipated size.The expressed protein quantity induced by 0.2 mmol/L IPTG treatment for 2 h at 37 ℃ is the highest.This study provide a fundamental condition supporting research on function ofGhHSP70 gene.
cotton;heat shock protein 70;gene cloning;prokaryotic expression
1000-4025(2015)04-0688-06
10.7606/j.issn.1000-4025.2015.04.0688
2014-12-17;修改稿收到日期:2015-01-30
新疆维吾尔自治区自然科学基金(2014211A025)
刘 娜(1990-),女,在读博士研究生,主要从事棉花分子育种研究。E-mail:416549170@qq.com
Q785;Q786
A
*通信作者:倪志勇,博士,副教授,主要从事植物逆境分子生物学研究。E-mai:nizhiyong@126.com
猜你喜欢
环球时报(2022-09-20)2022-09-20
中国动物保健(2022年2期)2022-05-05
今日农业(2020年24期)2020-12-15
中国洗涤用品工业(2019年4期)2019-05-11
中成药(2018年1期)2018-02-02
中成药(2017年3期)2017-05-17
中南民族大学学报(自然科学版)(2015年2期)2015-12-16
安徽医科大学学报(2015年9期)2015-12-16
小资CHIC!ELEGANCE(2015年15期)2015-09-01
现代检验医学杂志(2015年4期)2015-02-06
