
植物叶片氮分配及其影响因子研究进展
2015-01-19 02:57史作民唐敬超程瑞梅刘世荣
生态学报 2015年18期
史作民, 唐敬超, 程瑞梅, 罗 达, 刘世荣
中国林业科学研究院森林生态环境与保护研究所, 国家林业局森林生态环境重点实验室, 北京 100091
植物叶片氮分配及其影响因子研究进展
史作民*, 唐敬超, 程瑞梅, 罗 达, 刘世荣
中国林业科学研究院森林生态环境与保护研究所, 国家林业局森林生态环境重点实验室, 北京 100091
氮是植物生长的基本限制性因子,它的有效利用可以增加植物的适应性。叶片氮分配是指氮在植物叶片细胞各细胞结构以及游离化合物中所分配的比例。叶片氮的分配方式决定了叶片光合作用的强弱,影响叶片的坚韧程度以及化学防御强度,因此研究氮在植物叶片内的分配方式具有重要意义。阐述了叶片氮分配的方式,分析了影响叶片氮分配的生物和非生物因子(CO2,光,土壤养分),介绍了常用的叶片氮分配的研究方法,并对未来的研究进行了展望。
氮分配; 叶片; 影响因子; 研究进展
作为植物生长的必需元素之一,氮是很多生态系统的限制因子[1-3]。与其他元素相比,氮的有效性对植物生长能力的限制更强[4-5]。原因在于氮能通过调节植物体内一百多种基因来影响其生理过程,包括编码水通道蛋白、磷酸根和钾离子转运蛋白、硝酸盐转运蛋白和硝酸亚硝酸还原酶的基因和一些代谢酶,如天冬酰胺合成酶和组氨酸脱羧酶等[6]的基因。此外,氮也参与合成细胞里的其他的一些物质,如游离氨基酸[7],生氰糖苷[8]等。植物吸收到叶片中的氮存在于叶片细胞的不同细胞结构以及一些游离的化合物当中,这种分配方式决定了叶片光合作用的强弱[9-10],影响叶片的坚韧程度[11]以及化学防御强度[8],因此研究氮在植物叶片内的分配具有重要意义。
目前,关于叶片氮分配的研究国外已有大量报道,有些研究甚至已经深入到分子水平[1-5,7-11]。国内的研究也逐步开展,有张亚杰等对斜叶榕(Ficustinctoria)和假斜叶榕(F.subulata)[12]、Feng[13-16]对紫茎泽兰(Ageratinaadenophora)、Lei等[17]对互花米草(Spartinaalterniflora)和张绪成等[18-20]对春小麦(Triticumaestivum)的研究等。本文对叶片氮分配的研究进行综合评述,阐述了叶片氮的分配方式,分析了生物因子和非生物因子(光,CO2,土壤养分)对叶片氮分配的影响,并介绍了常用的研究方法,以期促进国内相关研究的深入开展。
1 叶片氮分配
叶片氮分配是指氮在植物叶片细胞各细胞结构以及游离化合物中的分配比例。叶片中的氮一般分配到细胞壁、细胞膜[11,21]、细胞核[22-23]以及细胞质中的叶绿体和线粒体等细胞器中,也有一些以游离化合物的形式存在。通常超过半数(最高到75%)的叶片氮会分配到叶绿体中参与光合作用[24-26]。参与光合作用的氮越多,植物生长越快[21]。光合作用强度与单位叶面积含氮量之比称为光合氮利用效率(PNUE)[10]。分配到叶绿体的氮缺乏可诱导光合抑制物的增加,阻碍光合进程和光合效率的提高[27]。
叶片细胞壁的氮分配量可以间接表示叶片的坚韧程度,这是植物基本的自卫手段[28-29]。有10%—30%的氮会分配到其中[30]。比叶重(LMA)与细胞壁氮含量呈显著相关[1],并且与分配到叶绿体的氮含量有关联[14]。生存环境较为恶劣的情况下,更多的氮会分配到细胞壁和一些游离化合物中[24]。如果在细胞壁里氮分配变多,那么分配到叶绿体的氮就会减少[1],这其中存在一个取舍关系[1,9,31]。但也有研究证明这种取舍关系是不可靠的[32]。一般认为,当生长环境较好,植物生长迅速,分配到叶绿体中的氮就会增加,而分配到细胞壁和游离化合物中的氮就会减少,比叶重减小;当环境恶劣时,植物会分配更多氮到细胞壁和游离化合物中。
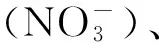
2 影响叶片氮分配的生物因子
不同生物学特性的植物叶片氮分配具有差异。光合氮利用效率越高的植物分配到Rubisco中的氮越多[37-38]。常绿植物[34,39]、耐胁迫植物[40]、演替后期植物[41]、高纬度植物[37,42]以及叶片寿命较长的植物[43]PNUE通常较低,也就意味着分配到Rubisco中的氮较少。Wright等总结了2500种植物的叶片功能性状后也得出叶片寿命较长的植物分配到Rubisco的氮较少的结论[44]。
大多数C3植物中,叶片水溶蛋白的含量约是叶片蛋白总量的一半以上,C4植物中则为1/4[45]。草本植物将更多的氮分配到Rubisco中[22,46-47]。例如Galmes等在巴利阿里群岛(Balearic Islands)对24个双子叶植物的研究表明,草本中Rubisco中的氮占叶片氮的13.2%—33.1%,高于木本植物[47]。有研究表明,高LMA植物分配到Rubisco中的氮较少[1],然而相似研究证实二者没有必然联系[37-38,48]。常绿植物分配到不活跃Rubisco中的氮较多,且分配到细胞壁中的氮的比例要高于落叶植物[1,34],这被Hikosaka和Shigeno对青叶山(Aobayama-hill)26种植物的研究结果所证明[49]。Zhu等研究发现,非固氮植物花花柴(Kareliniacaspica)和骆驼蓬(Peganumharmala)分配到叶绿体中的氮要少于固氮植物骆驼刺(Alhagisparsifolia)[50]。
入侵种为了迅速占领入侵区域,较少的将氮分配到细胞壁中,而将大量氮分配到叶绿体中以迅速生长[13-16,21]。Feng等对紫茎泽兰的研究发现,入侵的紫茎泽兰分配到叶绿体中的氮要比原生的紫茎泽兰多30%,最大净光合速率高24.4%,光合氮利用效率高20.2%,分配到细胞壁的氮低46.5%,而单位叶面积氮含量无显著差异[21]。Qing等对互花米草的研究表明,入侵的互花米草分配到Rubisco的氮含量以及比例都高于原生的互花米草,而单位叶面积氮含量差异不大[51]。
有些化学防御物质也有氮参与合成,氮分配到这些物质中的比例也比较可观。以生氰糖苷为例,不同树种分配到这种物质中的氮比例有差异,糖桉(Eucalyptuscladocalyx)为9.1%—15%;多花桉(E.polyanthemos)为0.2%;亚拉桉(E.yarraensis)为1.6%[8,31]。
3 影响叶片氮分配的非生物因子
由于大部分叶片氮会分配叶绿体中参与光合作用,因此影响光合作用的因子,如光、CO2和土壤养分等也会对氮在叶片中的分配产生影响。
3.1 光
光照的变化影响叶片氮的分配主要是因为它影响了叶片光合作用的能量供应,导致氮分配到光反应与暗反应的比例发生变化。高光强条件下暗反应能力成为制约光合作用的主要因素,因此叶片分配更多的氮到Rubisco中,而较少分配到光反应相关蛋白中[16,22,52-55]。Le[54]等对桃子(Prunuspersica)的研究、Feng[16]对紫茎泽兰以及Osada等[56]对库页蓼(Polygonumsachalinense)的研究支持这种观点。但也有研究发现高光强条件下叶片分配氮到Rubisco中比例下降,如张亚杰与冯玉龙的研究表明,生长光强低于自然光强的12%时,斜叶榕和假斜叶榕分配到Rubisco的氮比例(PC)随光强升高而升高;当生长光强高于自然光强的12%后,两种榕树的PC则随光强升高呈降低趋势[12]。
强光下叶片叶绿素降解较快[57-58],如果同时具有高大气CO2浓度则加剧了叶绿素降解[57-59]。Takashima等研究表明,低光条件下(30%自然光)的栎属植物比生长在高光条件下(90%自然光)的具有更高的叶绿素含量[1]。但张绪成对甘肃春小麦的研究则表明,与全光照下相比,遮阴后小麦叶绿素含量无明显差异[19]。
遮阴后植物叶片氮会增加[60-61]。植物将更多的氮分配到叶绿体中,减少次级含氮代谢物的产量[8,13,31]。Burns等对糖桉的研究表明,遮阴(40%全光照)会减少生氰糖苷的产生量[8]。Miller等对P.turneriana的研究也支持上述观点[62]。
3.2 二氧化碳
二氧化碳是光合作用的原料,环境中CO2浓度的变化会对光合作用以及光合氮利用效率产生影响[26,63-64],进而对叶片氮分配产生影响。外界CO2浓度增加,会间接提高羧化位CO2浓度,使羧化反应的原料增加。相对的,当CO2浓度减小到一定程度时,暗反应成为限制因子,分配到Rubisco的氮会相对增加[65-66]。这其中存在一个最优的比例关系[67],因此前人的研究得出了很多不同的结论。如Akita等的研究表明,在营养充足情况下,大气CO2浓度下培养的库页蓼比高CO2浓度下(700 μmol/mol)培养的分配到Rubisco中的氮比例略有增加[68]。Seneweera等对水稻(Orazysativa)的研究表明,大气CO2浓度(390 μmol/mol)与高CO2浓度下(1000 μmol/mol)培养的水稻相比,分配到Rubisco的氮比例在展叶期下降18.8%—16.2%,成熟期却提高14.9%—17.1%[69]。Lei等研究表明,在高CO2浓度条件下,紫茎泽兰分配到叶绿体的氮比例变化不大,但分配到细胞壁的氮比例增加[17]。这些研究结果的差异很可能与不同植物种类和不同发育时期对CO2需求量及对高CO2的反应差异有关。
长时间过高大气CO2浓度处理会促进叶片氮含量下降和叶绿素降解,导致光合机构活性下降[56,70-71]。Osada等对库页蓼的研究表明,随着CO2浓度增加,分配到叶绿素的氮含量有所下降[56]。王建林等的研究证明,经过60 d高CO2(750 μmol/mol)处理后会造成植物最大羧化效率降低,Rubisco的活性降低、含量减少[71]。
3.3 土壤养分
土壤中氮含量的变化会直接影响植物体内氮含量,进而对叶片氮分配造成影响,土壤中其他元素含量的变化也会对叶片氮分配产生影响,而叶片氮分配又会影响到叶片的光合作用及其化学防御强度等,进而影响植物的生长发育和环境适应能力,因此植物叶片氮分配对植物营养学的研究也具有一定的参考价值。
3.3.1 氮
随着土壤氮含量的增加,植物叶片内氮含量增加,单位叶面积的Rubisco含量也随之增加,这种现象在欧洲赤松(Pinussylvestris)[72]、火炬松(P.taeda)[73]中发现。随着土壤氮含量的增加,分配到Rubisco的氮的比例也有所提高,这被Akita等对库页蓼的研究所证明[68]。植物叶片叶绿素含量同样随着土壤氮含量的增加而增加。张绪成与上官周平的研究表明,土壤适量施氮后小麦叶绿素a含量显著提高[74]。Boussadia等发现,土壤高氮处理的两个油橄榄品种“Meski”和“Koroneiki”叶绿素a浓度分别为76、80 μg/cm2,经过减少氮供应处理之后,两种橄榄的叶绿素a含量都显著减少[75]。随着土壤氮含量的增加,叶片内分配到游离氨基酸的氮也有所增加。Ruan等研究表明,提高土壤氮供应可以提高分配到茶叶(Camelliasinensis)幼叶游离氨基酸的氮比例[7]。
3.3.2 磷
磷是组成转录蛋白质所必须的核糖核酸(Ribonucleic acid, RNA)的原料[76-77],会直接影响蛋白质的合成。同时磷也是合成ATP和Rubisco的原料[78],因此会影响氮分配到光合作用中的比例[79-80]。土壤增施磷肥可以增加叶片Rubisco含量和叶绿素含量。Warren研究表明,海岸松(P.pinaster)Rubisco中的氮含量与增施氮肥相关性不明显,而与增施磷肥相关性较高[77]。Tak等对鹰嘴豆(Cicerarietinum)进行4个梯度土壤磷施肥研究表明,施磷可以提高叶片叶绿素的含量,但分配到叶绿素的氮比例有所下降[79]。
3.3.3 钾
在正常的土壤钾供应水平下,叶片的叶绿素合成、运转会保持动态平衡,而钾的缺乏或过量会导致叶绿素含量、ATP含量以及Rubisco含量降低[81-82],并使光合电子传递及光合磷酸化受阻[83]。杨军等对奈李(P.salicina)的研究表明,土壤适度增施钾肥可以提高叶片叶绿素的含量[83]。郑炳松等对水稻的研究表明,土壤适度增施钾肥可以提高叶片的Rubisco含量[81]。
4 叶片氮分配的研究方法
4.1 化学分离法
化学分离法是利用不同含氮物质的化学特性,使用化学试剂将其从叶片中分离的方法。其优点是结果较为准确,能够直接获取氮含量数据;缺点是操作过程较为繁琐,容易出现人为提取误差,且费用较高。这种方法适合样本较少时叶片氮含量的测定。
蛋白质的分离与提取在氮分配研究中应用较多[1,11,21,24]。最早的一些研究没有将水溶蛋白与化学可溶蛋白分开,而是统称为可溶蛋白[84-85]。Takashima等[1]将细胞内蛋白质细分为水溶蛋白、化学可溶蛋白以及化学不溶蛋白,然后利用化学试剂将其分别提取。叶绿素也可以通过化学方法提取,在实验中应用较多的为丙酮提取法。Funk等不仅用化学方法分离了化学可溶、化学不溶与水溶蛋白,还提取了氨基酸与核酸,测定了叶片内的硝态氮与氨态氮含量[35]。
4.2 公式法
公式法是利用容易获得的叶片生理生态参数,利用经验公式来估算叶绿体中各含氮部分的氮含量的方法。其优点是操作简便,节省费用,只需获取部分比较容易获得的叶片生理生态参数就能估算;缺点是误差较大,计算结果不精确,无法获得细胞壁氮含量。这种方法适合样本较多时叶片氮含量的估算。
光合作用消耗氮的比例是研究的热点。早先的研究将参与光合的含氮物质按参与的阶段不同分为羧化系统(主要是Rubisco,参与暗反应)与类囊体(参与光反应)两部分[24,85],Evans和Seemann又将类囊体细分为生物力能学组分(Cytochrome f, Cyt f、耦合因子以及铁氧还蛋白-NADP+还原酶)与捕光系统(① 光系统Ⅰ(Photosystem Ⅰ, PSⅠ);② 光系统Ⅱ (Photosystem Ⅱ, PSⅡ);③ 光系统II捕光复合体(Light-harvesting complex Ⅱ, LHCⅡ))[22],但后人有时仍将类囊体作为一个整体进行研究。
Friend[86]在研究C3植物叶片时使用了最大羧化效率Vcmax和最大电子传递速率Jmax来估算氮分配到羧化系统和类囊体中的比例,分别以Rubisco和叶绿素作为代表:
Jmax=0.25×jmax×Nchl×N
式中,Vcmax为最大羧化效率(mol m-2s-1);Jmax最大电子传递速率(Eq m-2s-1,Farquhar[87]所使用的单位);kc为Rubisco周转次数,与温度相关,但温度对其值影响有限,在25℃时可取值2.5[87];Nrub为氮分配到Rubisco中的比例;Nchl为氮分配到叶绿素中的比例;N为叶片氮含量(mol/m2);jmax为单位叶绿素最大电子传递速率(Eq mol-1s-1),其计算公式为:
式中,E为相关活化能,取值为37000 (J/mol);R为通用气体常数8.314 (J K-1mol-1);T为叶片温度(K);H为去活化能,取值为220000 (J/mol);S为熵,取值为710 (J K-1mol-1)[87]。这种拟合方法以叶绿素含量代替类囊体的氮含量,忽略了类囊体上的其他含氮物质,对含氮物质的估算并不全面。
Evans[88-89]在研究紫花苜蓿(Medicagosativa)时,使用了最大电子传递速率与叶绿素含量的比值jmax来估算类囊体氮含量以及羧化系统氮含量:
T=0.068×jmax+34.4
S=0.33×jmax
式中,jmax最大电子传递速率(mmol 电子-mol-1Chl s-1),T为类囊体氮含量与叶绿素含量的比值(mol N/mol Chl)[88],S为羧化系统氮含量与叶绿素含量的比值(mol N/mol Chl)[89]。这种拟合方法比较简单,只使用了最大电子传递速率与叶绿素含量的比值进行公式拟合,但这就造成了其精确度不高;且公式的拟合是建立在单一植物研究结果之上的,在其他植物中使用效果较差[89]。
Niinemets和Tenhunen在对糖枫(Acersaccharum)的研究中,把参与光合作用的含氮物质分为了三部分:① 羧化系统;② 生物力能学组分;③ 捕光系统,并建立了一种能分别计算这三部分氮含量的模型[90]。其计算各部分氮含量的方法如下:
式中,PR为分配到羧化系统中的氮的比例;PB为分配到生物力能学组分的氮的比例;PL为分配到捕光系统的氮的比例;MA为单位面积叶干重(g/m2);Nm为单位干重叶片氮含量(g/g);Vcmax最大羧化效率(μmol m-2s-1);Jmax单位为(μmol m-2s-1);CC为叶片叶绿素浓度(mmol/g);Vcr是Rubisco比活,即单位Rubisco的CO2固定活性(μmol CO2g-1Rubisco s-1),与温度相关;Jmc为单位Cyt f的最大电子传递速率(μmol 电子 μmol-1cyt f s-1),与温度相关[90]。Vcr和Jmc的计算方法如下[91-92]:
式中,R为通用气体常数,取值为8.314 (J K-1mol-1);Tk是叶片温度(K),ΔHa、ΔHd、ΔS和c为活化能、去活化能、熵和比例常数,计算Vcr时的取值分别为74000 (J/mol)、203000 (J/mol)、645 (J K-1mol-1)和32.9;计算Jmc时的取值分别为24100 (J/mol)、564150 (J/mol)、1810 (J K-1mol-1)和14.77[93-94]。
CB为捕光系统中叶绿素与氮的比值(mmol Chl/g N),其取值可以利用拟合出的CB的值与MA的值的关系的线性公式来进行计算,其相关系数较高(r2=0.97),并且可以在多种植物类型中广泛应用。其公式为:
式中,[MA]为单位面积叶片干重(MA,g/m2)的值,[CB]为CB的值[90]。这样就可以通过MA的值来计算出CB的值,以便进一步计算PL。
这种氮分配的拟合方法不仅考虑了植物的羧化能力以及电子传递能力,还使用了单位面积叶片干重,叶绿素含量等作为公式中的变量,考虑比较全面,因此在最近的研究中得到广泛应用[1,12-15,35,95];且公式所拟合的数据可信度高,在很多研究中得到了验证;其叶片氮的分区也较为合理,是比较方便且准确的公式拟合方法。
5 叶片氮分配研究展望
5.1 叶片中其他含氮物质的氮分配
氮在叶片细胞内的分配比例能够反映出植物的生理特性及其在不同环境下所采取的对策。目前对叶片氮分配的研究主要集中在了叶绿体、细胞壁以及一些游离化合物中。然而植物叶片内ATP、不参与光合作用的酶、核酸[22-23]、游离氨基酸[7,23]中也会分配一部分的氮,这些含氮物质在植物生长发育和环境适应等方面也具有重要作用。例如,游离氨基酸是蛋白质等含氮化合物合成与分解过程的中介物质,可反映植物体内的氮代谢变化及植物对氮的吸收、运输、同化等状况[96-97],并且具有调节液泡渗透势、调节光呼吸氮代谢以及去除植物体内的NH3和SO2等物质的解毒作用[98]。未来应该加强氮在上述含氮物质中的分配及其与植物生长和环境适应性的关系研究。
另外,目前的研究多关注各种影响因子对单个含氮物质含量的影响,较少涉及这种物质所分配的氮占总氮的比例,而氮分配的比例变化或许比含氮物质的含量变化更能说明植物的叶片氮分配倾向。因此未来应该着重研究植物叶片在生物因子和非生物因子影响下氮分配比例的变化,以期更好的了解植物叶片氮分配的生理生态特征。
5.2 水分和温度的影响
影响叶片氮分配的因子很多,水也是其中之一。水分亏缺将导致参与光合作用的氮减少。在沙漠地区高水分利用效率的植物具有高光合氮利用效率[50],但也有研究得出水分利用效率与光合氮利用效率之间关系不明显的结论[14,99]。水分胁迫下叶绿体膨胀,排列紊乱,基质片层模糊,基粒间连接松弛,类囊体层肿胀或解体,光合器官的超微结构遭到破坏,叶绿素、核酸和蛋白质等大分子遭到破坏或损伤[100],且在水分胁迫下卡尔文循环中间物质含量减少,RuBP合成受到限制[101]。也有研究表明缺水可以提高叶片寿命,延长氮的利用时间,提高氮的利用效能[1]。因此了解植物叶片在水分变化下的氮分配具有重要的科学意义,值得继续深入研究。
酶活性存在最适合的温度范围,过高或过低的温度会使酶失活,进而影响光合作用以及氮的分配。有研究表明,植物在高温下通过叶绿素的降解来减少接收太阳辐射的光量子数,以防止体内温度过高或产生多余的自由基对植物造成伤害[102]。高温也可以抑制植物细胞器和细胞的功能,如降低光合器官的光能利用、降低PSⅡ活性[103-104]。植物在无法耐受的低温条件下,叶片蛋白质降解,游离氨基酸会增加;在可耐受的低温条件下脯氨酸含量增加,植物抗寒性增强[105],叶绿素含量下降[106]。关于温度对叶片氮分配影响的研究还较少见,在未来可以侧重对这方面的研究。
5.3 技术方法
目前在利用化学方法提取不同性质蛋白质的时候存在提取不彻底的问题,从而使蛋白质含量出现偏差:化学不溶蛋白有可能提取不彻底;不能提取可溶于柠檬酸和乙醇的蛋白质[1];含有树脂的树种的水溶性蛋白的含量也可能被低估[35],因此总蛋白质含量可能被低估。有研究表明,在高粱(Sorghumbicolor)[107]、冬小麦(T.aestivuml)[108]、北海道黄杨(Euonymusjaponicus)[109]和枣树(Zizyphusjujuba)[110]等的叶片蛋白质提取中,采用新方法或者添加新配方裂解液可以显著提高蛋白质的获得量。其它含氮化合物的含量也可能被低估,如叶绿素的提取存在提取不彻底的问题。研究表明,采用最新的超声波混合提取剂法比以往的研磨法提取的菠菜叶绿素含量要高45.1%[111]。同时,利用公式拟合各种含氮物质的氮含量也会出现偏差[91]。植物叶片含氮物质提取技术和方法的改进以及植物叶片各部分氮含量拟合公式精度的提高也是本领域未来的研究重点之一。
[1] Takashima T, Hikosaka K, Hirose T. Photosynthesis or persistence: nitrogen allocation in leaves of evergreen and deciduousQuercusspecies. Plant, Cell & Environment, 2004, 27(8): 1047-1054.
[2] Vitousek P M, Porder S, Houlton B Z, Chadwick O A. Terrestrial phosphorus limitation: mechanisms, implications, and nitrogen-phosphorus interactions. Ecological Applications, 2010, 20(1): 5-15.
[3] Reich P B, Oleksyn J. Global patterns of plant leaf N and P in relation to temperature and latitude. Proceedings of the National Academy of Sciences of the United States of America, 2004, 101(30): 11001-11006.
[4] Raun W R, Johnson G V. Improving nitrogen use efficiency for cereal production. Agronomy Journal, 1999, 91(3): 357-363.
[5] Vitousek P M, Howarth R W. Nitrogen limitation on land and in the sea: how can it occur? Biogeochemistry, 1991, 13(2): 87-115.
[6] Wang Y H, Garvin D F, Kochia L V. Nitrate-induced genes in tomato roots. Array analysis reveals novel genes that may play a role in nitrogen nutrition. Plant Physiology, 2001, 127(1): 345-359.
[7] Ruan J, Haerdter R, Gerendás J. Impact of nitrogen supply on carbon/nitrogen allocation: a case study on amino acids and catechins in green tea [Camelliasinensis(L.) O. Kuntze] plants. Plant Biology, 2010, 12(5): 724-734.
[8] Burns A E, Gleadow R M, Woodrow I E. Light alters the allocation of nitrogen to cyanogenic glycosides inEucalyptuscladocalyx. Oecologia, 2002, 133(3): 288-294.
[9] Hikosaka K, Osone Y. A paradox of leaf-trait convergence: why is leaf nitrogen concentration higher in species with higher photosynthetic capacity? Journal of Plant Research, 2009, 122(3): 245-251.
[10] Hikosaka K. Interspecific difference in the photosynthesis-nitrogen relationship: patterns, physiological causes, and ecological importance. Journal of Plant Research, 2004, 117(6): 481-494.
[11] Onoda Y, Hikosaka K, Hirose T. Allocation of nitrogen to cell walls decreases photosynthetic nitrogen-use efficiency. Functional Ecology, 2004, 18(3): 419-425.
[12] 张亚杰, 冯玉龙. 不同光强下生长的两种榕树叶片光合能力与比叶重、氮含量及分配的关系. 植物生理与分子生物学学报, 2004, 30(3): 269-276.
[13] Feng Y L, Fu G L. Nitrogen allocation, partitioning and use efficiency in three invasive plant species in comparison with their native congeners. Biological Invasions, 2008, 10(6): 891-902.
[14] Feng Y L, Fu G L, Zheng Y L. Specific leaf area relates to the differences in leaf construction cost, photosynthesis, nitrogen allocation, and use efficiencies between invasive and noninvasive alien congeners. Planta, 2008, 228(3): 383-390.
[15] Feng Y L, Auge H, Ebeling S K. InvasiveBuddlejadavidiiallocates more nitrogen to its photosynthetic machinery than five native woody species. Oecologia, 2007, 153(3): 501-510.
[16] Feng Y L. Photosynthesis, nitrogen allocation and specific leaf area in invasiveEupatoriumadenophorumand nativeEupatoriumjaponicumgrown at different irradiances. Physiologia Plantarum, 2008, 133(2): 318-326.
[17] Lei Y B, Feng Y L, Zheng Y L, Wang R. F, De G H, Zhang Y P. Innate and evolutionarily increased advantages of invasiveEupatoriumadenophorumover nativeE.japonicumunder ambient and doubled atmospheric CO2concentrations. Biological Invasions, 2011, 13(12): 2703-2714.
[18] 张绪成, 于显枫, 高世铭. 高大气 CO2浓度下氮素对小麦叶片光能利用的影响. 植物生态学报, 2010, 34(10): 1196-1203.
[19] 张绪成, 于显枫, 马一凡, 上官周平. 高大气 CO2浓度下小麦旗叶光合能量利用对氮素和光强的响应. 生态学报, 2011, 31(4):1046-1057
[20] 张绪成, 于显枫, 马一凡. 施氮和大气 CO2浓度升高对小麦旗叶光合电子传递和分配的影响. 应用生态学报, 2011, 22(3): 673-680.
[21] Feng Y L, Lei Y B, Wang R F, Callaway R. M, Valiente B A, Li Y P, Zheng Y L. Evolutionary tradeoffs for nitrogen allocation to photosynthesis versus cell walls in an invasive plant. Proceedings of the National Academy of Sciences of the United States of America, 2009, 106(6): 1853-1856.
[22] Evans J R, Seemann J R. The allocation of protein nitrogen in the photosynthetic apparatus: costs, consequences and control. Plant biology, 1989, 8(1):183-205.
[23] Chapin III F S. The cost of tundra plant structures: evaluation of concepts and currencies. American Naturalist, 1989, 133(1):1-19.
[24] Evans J R. Photosynthesis and nitrogen relationships in leaves of C3plants. Oecologia, 1989, 78(1): 9-19.
[25] Warren C R, Adams M A. Phosphorus affects growth and partitioning of nitrogen to Rubisco inPinuspinaster. Tree Physiology, 2002, 22(1): 11-19.
[26] Warren C R, Adams M A. Internal conductance does not scale with photosynthetic capacity: implications for carbon isotope discrimination and the economics of water and nitrogen use in photosynthesis. Plant, Cell & Environment, 2006, 29(2): 192-201.
[27] Henley W J, Levavasseur G, Franklin L A, Osmond C B, Ramus J. Photoacclimation and photoinhibition inUlvarotundataas influenced by nitrogen availability. Planta, 1991, 184(2):235-243.
[28] Ridenour W M, Vivanco J M, Feng Y, Horiuchi J I, Callaway R M. No evidence for trade-offs:Centaureaplants from America are better competitors and defenders. Ecological Monographs, 2008, 78(3): 369-386.
[29] Coley P D. Herbivory and defensive characteristics of tree species in a lowland tropical forest. Ecological Monographs, 1983, 53(2): 209-234.
[30] Stitt M, Schulze D. Does RuBPCase control the rate of photosynthesis and plant growth? An exercise in molecular ecophysiology. Plant, Cell & Environment, 1994, 17(5): 465-487.
[31] Goodger J Q D, Gleadow R M, Woodrow I E. Growth cost and ontogenetic expression patterns of defence in cyanogenicEucalyptusspp. Trees, 2006, 20(6): 757-765.
[32] Harrison M T, Edwards E J, Farquhar G D, Nicotra A B, Evans J R. Nitrogen in cell walls of sclerophyllous leaves accounts for little of the variation in photosynthetic nitrogen-use efficiency. Plant, Cell & Environment, 2009, 32(3): 259-270.
[33] Murray M B, Smith R I, Friend A, Jarvis P G. Effect of elevated [CO2] and varying nutrient application rates on physiology and biomass accumulation of Sitka spruce (Piceasitchensis). Tree Physiology, 2000, 20(7): 421-434.
[34] Warren C R, Adams M A. Evergreen trees do not maximize instantaneous photosynthesis. Trends in Plant Science, 2004, 9(6): 270-274.
[35] Funk J L, Glenwinkel L A, Sack L. Differential allocation to Photosynthetic and Non-Photosynthetic nitrogen fractions among native and invasive species. PloS One, 2013, 8(5): e64502.
[36] 许振柱, 周广胜. 植物氮代谢及其环境调节研究进展. 应用生态学报, 2004, 15.(3): 511-516.
[37] Westbeek M H M, Pons T L, Cambridge M L, Atkin O K. Analysis of differences in photosynthetic nitrogen use efficiency of alpine and lowlandPoaspecies. Oecologia, 1999, 120(1): 19-26.
[38] Ripullone F, Grassi G, Lauteri M, Borghetti M. Photosynthesis-nitrogen relationships: interpretation of different patterns betweenPseudotsugamenziesiiandPopulus×euroamericanain a mini-stand experiment. Tree Physiology, 2003, 23(2): 137-144.
[39] Field C, Mooney H A. Leaf age and seasonal effects on light, water, and nitrogen use efficiency in a California shrub. Oecologia, 1983, 56(2/3): 348-355.
[40] Chazdon R L, Field C B. Determinants of photosynthetic capacity in six rainforestPiperspecies. Oecologia, 1987, 73(2): 222-230.
[41] Ellsworth D S, Reich P B. Photosynthesis and leaf nitrogen in five Amazonian tree species during early secondary succession. Ecology, 1996, 77(2): 581-594.
[42] Hikosaka K, Nagamatsu D, Ishii H S, Hirose T. Photosynthesis-nitrogen relationships in species at different altitudes on Mount Kinabalu, Malaysia. Ecological Research, 2002, 17(3): 305-313.
[43] Reich P B, Uhl C, Walters M B, Ellsworth D S. Leaf lifespan as a determinant of leaf structure and function among 23 Amazonian tree species. Oecologia, 1991, 86(1): 16-24.
[44] Wright I J, Reich P B, Westoby M, Ackerly D D, Baruch Z, Bongers F, Cavender-Bares J, Chapin T, Cornelissen J H C, Diemer M, Flexas J, Garnier E, Groom P K, Gulias J, Hikosaka K, Lamont B B, Lee T, Lee W, Lusk C, Midgley J J, Navas M L, Niinemets ü, Oleksyn J, Osada N, Poorter H, Poot P, Prior L, Pyankov V I, Roumet C, Thomas S C, Tjoelker M G, Veneklaas E J, Villar R .The wordwide leaf economics spectrum. Nature, 2004, 428(6985): 821-827.
[45] Meinzer F C, Zhu J. Nitrogen stress reduces the efficiency of the C4CO2concentrating system, and therefore quantum yield, inSaccharum(sugarcane) species. Journal of Experimental Botany, 1998, 49(324): 1227-1234.
[46] Hikosaka K, Terashima I. Nitrogen partitioning among photosynthetic components and its consequence in sun and shade plants. Functional Ecology, 1996, 10(1): 335-343.
[47] Galmes J, Flexas J, Keys A J, Cifre J, Mitchell R. A, Madgwick P J, Parry M A. Rubisco specificity factor tends to be larger in plant species from drier habitats and in species with persistent leaves. Plant, Cell & Environment, 2005, 28(5): 571-579.
[48] Terashima I, Araya T, Miyazawa S I, Sone K, Yano S. Construction and maintenance of the optimal photosynthetic systems of the leaf, herbaceous plant and tree: an eco-developmental treatise. Annals of Botany, 2005, 95(3): 507-519.
[49] Hikosaka K, Shigeno A. The role of RuBPCase and cell walls in the interspecific variation in photosynthetic capacity. Oecologia, 2009, 160(3): 443-451.
[50] Zhu J T, Li X Y, Zhang X M, Yu Q, Lin L S. Leaf nitrogen allocation and partitioning in three groundwater-dependent herbaceous species in a hyper-arid desert region of north-western China. Australian Journal of Botany, 2012, 60(1): 61-67.
[51] Qing H, Cai Y, Xiao Y, Yao Y, An S. Leaf nitrogen partition between photosynthesis and structural defense in invasive and native tall formSpartinaalterniflorapopulations: effects of nitrogen treatments. Biological Invasions, 2012, 14(10): 2039-2048.
[52] Boardman N K. Comparative photosynthesis of sun and shade plants. Annual Review of Plant Physiology, 1977, 28(1): 355-377.
[53] Seemann J R, Sharkey T D, Wang J, Osmond C B. Environmental effects on photosynthesis, nitrogen-use efficiency, and metabolite pools in leaves of sun and shade plants. Plant Physiology, 1987, 84(3): 796-802.
[54] Le Roux X, Walcroft, A S, Daudet F A, Sinoquet H, Chaves M M, Rodrigues A, Osorio L. Photosynthetic light acclimation in peach leaves: importance of changes in mass: area ratio, nitrogen concentration, and leaf nitrogen partitioning. Tree Physiology, 2001, 21(6): 377-386.
[55] Niinemets Ü, Kull O, Tenhunen J D. An analysis of light effects on foliar morphology, physiology, and light interception in temperate deciduous woody species of contrasting shade tolerance. Tree Physiology, 1998, 18(10): 681-696.
[56] Osada N, Onoda Y, Hikosaka K. Effects of atmospheric CO2concentration, irradiance, and soil nitrogen availability on leaf photosynthetic traits ofPolygonumsachalinensearound natural CO2springs in northern Japan. Oecologia, 2010, 164(1): 41-52.
[57] Liberloo M, Tulva I, Raïm O, Kull O, Ceulemans R. Photosynthetic stimulation under long-term CO2enrichment and fertilization is sustained across a closedPopuluscanopy profile (EUROFACE). New Phytologist, 2007, 173(3): 537-549.
[58] Zhou Y, Lam H M, Zhang J. Inhibition of photosynthesis and energy dissipation induced by water and high light stresses in rice. Journal of Experimental Botany, 2007, 58(5): 1207-1217.
[59] Haake V, Geiger M, Walch L P, Engels C, Zrenner R, Stitt M. Changes in aldolase activity in wild-type potato plants are important for acclimation to growth irradiance and carbon dioxide concentration, because plastid aldolase exerts control over the ambient rate of photosynthesis across a range of growth conditions. The Plant Journal, 1999, 17(5): 479-489.
[60] Waterman P G, Ross J A M, Mckey D B. Factors affecting levels of some phenolic compounds, digestibility, and nitrogen content of the mature leaves ofBarteriafistulosa(Passifloraceae). Journal of Chemical Ecology, 1984, 10(3): 387-401.
[61] Höft M, Verpoorte R, Beck E. Growth and alkaloid contents in leaves ofTabernaemontanapachysiphonStapf (Apocynaceae) as influenced by light intensity, water and nutrient supply. Oecologia, 1996, 107(2): 160-169.
[62] Miller R E, Gleadow R M, Woodrow I E. Cyanogenesis in tropicalPrunusturneriana: characterisation, variation and response to low light. Functional Plant Biology, 2004, 31(5): 491-503.
[63] Flexas J, Ribas-Carbó M, Diaz-Espejo A, Antonio, Galmes J, Medrano H. Mesophyll conductance to CO2: current knowledge and future prospects. Plant Cell and Environment, 2008, 31(5):602-621.
[64] Terashima I, Hanba Y T, Tazoe Y, Vyas P, Yano S. Irradiance and phenotype: comparative eco-development of sun and shade leaves in relation to photosynthetic CO2diffusion. Journal of Experimental Botany, 2006, 57(2): 343-354.
[65] Wullschleger S D, Norby R J, Hendrix D L. Carbon exchange rates, chlorophyll content, and carbohydrate status of two forest tree species exposed to carbon dioxide enrichment. Tree Physiology, 1992, 10(1): 21-31.
[66] Hikosaka K, Hirose T. Leaf and canopy photosynthesis of C3plants at elevated CO2in relation to optimal partitioning of nitrogen among photosynthetic components: theoretical prediction. Ecological Modelling, 1998, 106(2): 247-259.
[67] Wullschleger S D. Biochemical limitations to carbon assimilation in C3plants-a retrospective analysis of the A/Ci curves from 109 species. Journal of Experimental Botany, 1993, 44(5): 907-920.
[68] Akita R, Kamiyama C, Hikosaka K.Polygonumsachalinensealters the balance between capacities of regeneration and carboxylation of ribulose-1, 5-bisphosphate in response to growth CO2increment but not the nitrogen allocation within the photosynthetic apparatus. Physiologia Plantarum, 2012, 146(4): 404-412.
[69] Seneweera S. Reduced nitrogen allocation to expanding leaf blades suppresses ribulose-1, 5-bisphosphate carboxylase/oxygenase synthesis and leads to photosynthetic acclimation to elevated CO2in rice. Photosynthetica, 2011, 49(1): 145-148.
[70] Chaiklahan R, Khonsarn N, Chirasuwan N, Ruengjitchatchawalya M, Bunnag B, Tanticharoen M. Response ofSpirulinaplatensisC1 to high temperature and high light intensity. Kasetsart Journal (Natural Sciences), 2007, 41(1): 123-129.
[71] 王建林, 温学发, 赵风华, 房全孝, 杨新民. CO2浓度倍增对 8 种作物叶片光合作用、蒸腾作用和水分利用效率的影响. 植物生态学报, 2012, 36(5): 438-446.
[72] Warren C R, Dreyer E, Adams M A. Photosynthesis-Rubisco relationships in foliage ofPinussylvestrisin response to nitrogen supply and the proposed role of Rubisco and amino acids as nitrogen stores. Trees, 2003, 17(4): 359-366.
[73] Tissue D T, Thomas R B, Strain B R. Long-term effects of elevated CO2and nutrients on photosynthesis and Rubisco in loblolly pine seedlings. Plant, Cell & Environment, 1993, 16(7): 859-865.
[74] 张绪成, 上官周平. 施氮对不同抗旱性冬小麦叶片光合与呼吸的调控. 应用生态学报, 2006, 17(11): 2064-2069.
[75] Boussadia O, Steppe K, Zgallai H, Ben El Hadj, S, Braham M, Lemeur R, Van Labeke M C. Effects of nitrogen deficiency on leaf photosynthesis, carbohydrate status and biomass production in two olive cultivars ‘Meski’and ‘Koroneiki’. Scientia Horticulturae, 2010, 123(3/4): 336-342.
[76] Pieters A J, Paul M J, Lawlor D W. Low sink demand limits photosynthesis under Pi deficiency. Journal of Experimental Botany, 2001, 52(358): 1083-1091.
[77] Warren C R, McGrath J F, Adams M A. Differential effects of N, P and K on photosynthesis and partitioning of N inPinuspinasterneedles. Annals of Forest Science, 2005, 62(1): 1-8.
[78] Reich P B, Oleksyn J, Wright I J. Leaf phosphorus influences the photosynthesis-nitrogen relation: a cross-biome analysis of 314 species. Oecologia, 2009, 160(2): 207-212.
[79] Tak H I, Ahmad F, Babalola O O, Inam A. Growth, photosynthesis and yield of chickpea as Influenced by urban wastewater and different levels of phosphorus. International Journal of Plant Research, 2012, 2(2): 6-13.
[80] Osaki M, Shinano T, Tadano T. Effect of nitrogen, phosphorus, or potassium deficiency on the accumulation of ribulose-1, 5-bisphosphate carboxylase/oxygenase and chlorophyll in several field crops. Soil Science and Plant Nutrition, 1993, 39(3): 417-425.
[81] 郑炳松, 蒋德安, 翁晓燕, 陆庆, 奚海福. 钾营养对水稻剑叶光合作用关键酶活性的影响. 浙江大学学报, 农业与生命科学版, 2001, 27(5): 489-494.
[82] 李玲, 王会肖, 张玉铭, 蔡燕, 刘丽芳. 施肥对小麦旗叶光合特性的影响研究. 中国生态农业学报, 2011,19(6): 1268-1271.
[83] 杨军, 马振峰, 刘桂华. 钾营养对柰李叶片光合作用及叶绿素荧光的影响. 中国农学通报, 2010, 26(20): 238-244.
[84] Evans J R. Nitrogen and photosynthesis in the flag leaf of wheat (TriticumaestivumL.). Plant physiology, 1983, 72(2): 297-302.
[85] Terashima I, Evans J R. Effects of light and nitrogen nutrition on the organization of the photosynthetic apparatus in spinach. Plant and Cell Physiology, 1988, 29(1): 143-155.
[86] Friend A D. Use of a model of photosynthesis and leaf microenvironment to predict optimal stomatal conductance and leaf nitrogen partitioning. Plant, Cell & Environment, 1991, 14(9): 895-905.
[87] Farquhar G, Caemmerer D S, Berry J A. A biochemical model of photosynthetic CO2assimilation in leaves of C3species. Planta 1980, 149(1): 78-90.
[88] Evans J R. Photosynthetic acclimation and nitrogen partitioning within a lucerne canopy. I. Canopy characteristics. Functional Plant Biology, 1993, 20(1): 55-67.
[89] Evans J R. Photosynthetic acclimation and nitrogen partitioning within a lucerne canopy. II. Stability through time and comparison with a theoretical optimum. Functional Plant Biology, 1993, 20(1): 69-82.
[90] Niinemets Ü, Tenhunen J D. A model separating leaf structural and physiological effects on carbon gain along light gradients for the shade-tolerant speciesAcersaccharum. Plant, Cell & Environment, 1997, 20(7): 845-866.
[91] Johnson F H, Henry E R. Williams W. The nature of enzyme inhibitions in bacterial luminescence: sulfanilamide, urethane, temperature and pressure. Journal of Cellular and Comparative Physiology 1942, 20(3): 247-268.
[92] Harley P C, Thomas R B, Reynolds J F, Strain B R. Modelling photosynthesis of cotton grown in elevated CO2. Plant, Cell & Environment, 1992, 15(3): 271-282.
[93] Jordan D B, William L O. The CO2/O2specificity of ribulose 1, 5-bisphosphate carboxylase/oxygenase. Planta, 1984, 161(4): 308-313.
[94] Nolan W G, Robert M S. Multi-temperature effects on Hill reaction activity of barley chloroplasts. Biochimica et Biophysica Acta (BBA)-Bioenergetics, 1976, 440(3): 461-475.
[95] 冯秋红, 程瑞梅, 史作民, 刘世荣, 王卫霞, 刘兴良, 何飞. 四川巴郎山齿果酸模叶片氮素及其分配的海拔响应. 植物生态学报, 2013, 37(7): 591-600.
[96] 陆若辉, 周焱, 董越勇, 娄烽, 何青芳. 核酸混配氨基酸在大棚草莓上应用的研究. 中国土壤与肥料, 2006,(5): 36-38.
[97] 李美云, 于明礼. 植物有机营养肥料研究进展. 安徽农业科学, 2007, 35(33): 10773-10775.
[98] 周青. 植物中氨基酸的生理作用. 生物学通报, 1986, (8): 7-9.
[99] Hill J P, Germino M J, Wraith J M, Olson B E, Swan M. B. Advantages in water relations contribute to greater photosynthesis inCentaureamaculosacompared with established grasses. International Journal of Plant Sciences, 2006, 167(2): 269-277.
[100] 孙梅霞, 祖朝龙, 徐经年. 干旱对植物影响的研究进展. 安徽农业科学, 2004, 32(2): 365-367, 384-384.
[101] Tezara W, Mitchell V J, Driscoll S D, Lawior D W. Water stress inhibits plant photosynthesis by decreasing coupling factor and ATP. Nature, 1999, 401(6756): 914-917.
[102] Tzeng S, Hsu B D. Chlorophyll degradation in heat-treatedChlorellapyrenoidosa. A flow cytometric study. Functional Plant Biology, 2001, 28(1): 79-83.
[103] Schrader S M, Kleinbeck K R, Sharkey T D. Rapid heating of intact leaves reveals initial effects of stromal oxidation on photosynthesis. Plant, Cell & Environment, 2007, 30(6): 671-678.
[104] 孙谷畴, 曾小平, 刘晓静, 赵平. 适度高温胁迫对亚热带森林3种建群树种幼树光合作用的影响. 生态学报, 2007, 27(4):1283-1291.
[105] 龚明, 刘友良, 朱培仁. 低温下稻苗叶片中蛋白质及游离氨基酸的变化. 植物生理学通讯, 1989, 25(4): 18-22.
[106] 邵怡若, 许建新, 薛立, 张柔, 吴彩琼, 卢广超. 低温胁迫时间对 4 种幼苗生理生化及光合特性的影响. 生态学报, 2013, 33(14): 4237-4247.
[107] 许克静, 陈云, 雷珍珍, 程海丽, 陈磊, 乐超银. 适于 SDSPAGE 分析的高粱叶片蛋白质提取方法. 河南农业科学, 2013, 42(5): 38-40.
[108] 李红兵, 康振生. 适于小麦叶片蛋白质组分析的样品提取方法研究. 西北植物学报, 2011,31(3): 632-638.
[109] 郝强, 葛秀秀, 张睿鹂, 尉莉莉, 关雪莲. 北方常绿阔叶木本植物叶片蛋白质双向电泳技术体系优化. 西北植物学报, 2010,30(9): 1906-1912.
[110] 杜绍华, 卜志国. 枣树叶片蛋白质提取及 SDS-PAGE 单向电泳条件优化. 安徽农业科学, 2012, 40(13): 7631-7632, 7655.
[111] 张秀君, 孙钱钱, 乔双, 朱海, 孙丹, 江丕文. 菠菜叶绿素提取方法的比较研究. 作物杂志, 2011,(3): 57-59,60-60.
A review of nitrogen allocation in leaves and factors in its effects
SHI Zuomin*, TANG Jingchao, CHENG Ruimei, LUO Da, LIU Shirong
KeyLaboratoryonForestEcologyandEnvironmentalSciencesofStateForestryAdministration,InstituteofForestEcology,EnvironmentandProtection,ChineseAcademyofForestry,Beijing100091,China
Nitrogen is an essential limiting resource for plant growth, and its effective utilization can increase the adaptability of plants. This is because nitrogen is required for the regulation of more than a hundred genes in plants and influences their physiological processes. Leaf nitrogen allocation is defined as the proportion of nitrogen distributed in every plant leaf cell and free compounds. Leaf nitrogen allocation not only determines the photosynthetic capacity of leaves but also affects their toughness and chemical defenses. Therefore, research on nitrogen allocation in leaves is of great importance. In this study, we evaluated the mode of leaf nitrogen allocation. Leaf nitrogen is generally distributed in the cell wall, cell membrane, nucleus, and organelles such as chloroplasts and mitochondria in the cytoplasm; leaf nitrogen is also found in free compounds. Usually more than half (up to 75%) of the leaf nitrogen is assigned to chloroplasts, which are involved in photosynthesis. An increase in the amount of nitrogen involved in photosynthesis results in faster plant growth. The amount of leaf nitrogen allocated to cell walls can indirectly result in blade tenacity, which is a basic method of plant self-defense; 10%—30% of the nitrogen is assigned to the cell walls. Leaf nitrogen is mainly found in the form of plant proteins, and usually 71%—77% of the leaf nitrogen is found in the form of proteins, followed by free amino acids, alkaloids, cyanogenetic glycosides, phospholipids, nucleic acids, nitrate nitrogen, ammonia nitrogen, and adenosine triphosphate. We also analyzed biological and environmental factors (carbon dioxide, illumination, and soil nutrients) that affect the allocation of leaf nitrogen. With respect to biological factors, a higher amount of nitrogen is assigned to Rubisco, resulting in an increase in the photosynthetic nitrogen use efficiency of plants. In most C3plants, more than half of the total leaf protein is soluble; in C4plants, a quarter of the total leaf protein is soluble. In herbs, a higher amount of nitrogen is allocated to Rubisco. In order to rapidly occupy an area, an invasive plant species assigns less nitrogen to the cell walls and more to the chloroplasts. With respect to environmental factors, illumination affects the distribution of leaf nitrogen, mainly because it affects the energy supply for photosynthesis; this results in a change in the amount of nitrogen assigned for light and dark reactions. Carbon dioxide is required for photosynthesis, and changes in the carbon dioxide concentration in the environment will affect photosynthesis, photosynthetic nitrogen use efficiency, and the distribution of leaf nitrogen. Changes in the nitrogen content in the soil will directly affect the nitrogen content in plants and, in turn, affect leaf nitrogen allocation. Changes in the content of other elements in the soil will also affect leaf nitrogen allocation. We have reported on the methods frequently used for measuring leaf nitrogen allocation. There are two commonly used methods: the formula method and chemical separation method. We have also discussed different perspectives for future research on leaf nitrogen allocation.
nitrogen allocation; leaves; effect factors; research progress
国家自然科学基金重大项目课题(31290223); 中国科学院碳专项(XDA05060100)
2014-01-24;
日期:2014-11-19
10.5846/stxb201401240184
*通讯作者Corresponding author.E-mail: shizm@caf.ac.cn
史作民, 唐敬超, 程瑞梅, 罗达, 刘世荣.植物叶片氮分配及其影响因子研究进展.生态学报,2015,35(18):5909-5919.
Shi Z M, Tang J C, Cheng R M, Luo D, Liu S R.A review of nitrogen allocation in leaves and factors in its effects.Acta Ecologica Sinica,2015,35(18):5909-5919.
猜你喜欢
科学(2022年4期)2022-10-25
阅读(科学探秘)(2020年8期)2020-11-06
铁道通信信号(2020年9期)2020-02-06
数学大王·趣味逻辑(2019年5期)2019-06-13
小学科学(学生版)(2019年5期)2019-05-21
中国果业信息(2019年1期)2019-01-05
生物学教学(2017年9期)2017-08-20
Coco薇(2016年5期)2016-06-03
考试周刊(2016年6期)2016-03-11
少儿科学周刊·儿童版(2015年10期)2015-11-07
