植酸酶对黄金鲫生长、饲料利用和体成分的影响
2015-01-18 12:46王桂芹
饲料工业 2015年6期
■李 萌 王桂芹
(吉林农业大学动物科学技术学院,吉林长春 130118)
磷是动物营养中的一种必需矿物元素,并且发挥着重要的作用。由于植物性蛋白源替代鱼粉,使得水产动物饲料中植物性原料增多,磷多数以植酸盐的形式存在,难以被水产动物利用。因此,需要在饲料中额外添加磷源,这样不仅提高了饲料的成本,又使未被利用的植酸磷排出体外,造成了养殖水体的负担。植酸酶可以促进植酸盐的水解,磷酸根可以被动物吸收利用,所以饲料中添加植酸酶对提高植物磷的利用率,促进动物生长具有较好的效果[1]。目前,植酸酶在水产动物营养中的研究主要集中在改善鱼类的生产性能、骨磷沉积、消化酶活性及磷代谢等方面的作用[2-3]。研究的品种主要集中在异育银鲫[4]、青鱼[5]和黄河鲤[6]等水产动物上,有关植酸酶对黄金鲫生长性能、饲料利用和体成分的系统研究未见报道。因此,本试验在全植物蛋白源的黄金鲫饲料中添加植酸酶研究对其生长性能、饲料利用和体成分的影响,以期为植酸酶在以植物蛋白源为主的鱼饲料中的广泛应用提供理论依据和实践指导。
1 材料与方法
1.1 材料
试验用黄金鲫幼鱼购自二八一四渔场。试验用植酸酶购自中国上海生物工程有限公司(酶活力为5 000 U/kg)。对照组添加磷酸二氢钙,试验组添加植酸酶水平分别为0、300、600、900、1 200 U/kg。植酸酶以次粉为载体进行稀释[7]。饲料原料粉碎过60目筛,根据配方称重、混匀,挤压为直径1.0 mm颗粒,晒干并置于4℃冰箱保存、备用。
试验饲料的配方及营养组成见表1和表2。
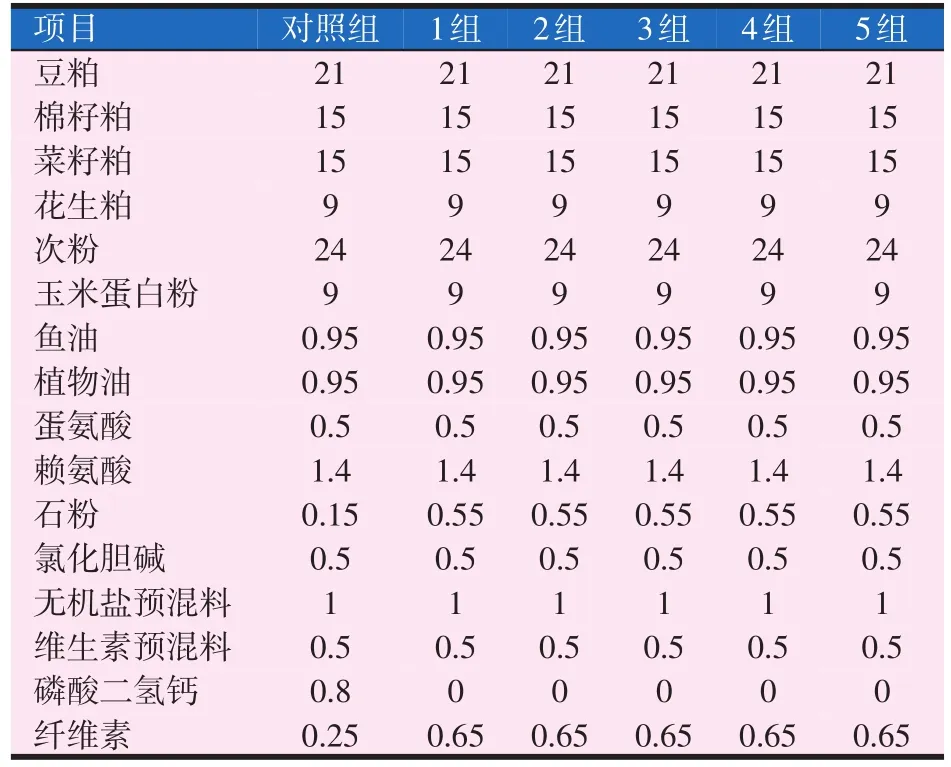
表1 黄金鲫试验饲料配方(%)
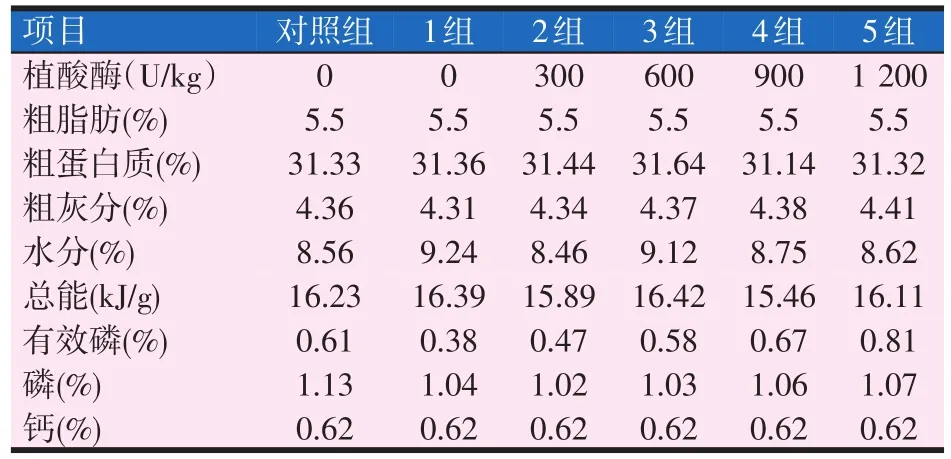
表2 黄金鲫试验饲料营养组成
1.2 饲养管理和样品收集
1.2.1 饲养管理
试验鱼先暂养15 d,期间投喂相同基础饲料,待试验鱼适应饲养环境后开始正式试验。试验共6组,每组3个重复,分别饲养在240 L水族箱中,每个水族箱放养30尾初始体重为(10.71±0.16)g的黄金鲫幼鱼。每天8:00和16:00投喂两次,24 h充气,试验饲养时间60 d,期间水温为20~24 ℃,溶解氧>6 mg/l。
1.2.2 样品的收集
1.3 测定指标
1.3.1 生长指标
特定生长率(%/d)=(lnWt-lnW0)×100/t;
平均增重率(%)=100×(Wt-W0)/W0;
肥满度(g/cm3)=(W/L3)×100。
式中:W0——鱼初始重(g);
Wt——末重(g);
《李仲文女》《冯孝将子》两篇小说,一以悲剧结束,一以喜剧告终。推其原因,都不在当事人,而实取决于双方的家长,客观上反映了当时家长对男女婚姻的支配决定权。汤显祖则汲取这二个故事的精神,推陈出新,创作了《牡丹亭》传奇,塑造出杜丽娘这个带有悲剧与喜剧两重性的形象。汤显祖所谓“予稍为更而演之”,显然是自谦之词。他所说的“仿佛”,正表明不是机械地照搬原来的传说故事。
t——试验天数(d);
W、L——分别为每尾鱼的体质量(g)、体长(cm)。
1.3.2 饲料利用指标
饲料效率(%)=100×(Wt-W0)/I;
饲料系数=I/(Wt-W0);
蛋白效率(%)=(Wt-W0)/(I×CP);
蛋白质生产率(%)=100×(Nt×Wt-N0×W0)/(I×N)。
式中:I——摄入饲料重(g);
W0——鱼初始重(g);
Wt——鱼末重(g);
CP——饲料蛋白质含量(%);
N、N0和Nt——饲料、初、末鱼体氮含量(%)。
1.3.3 体成分指标
采用105℃恒温烘干法(GB/T6435—1994)测定水分含量;采用凯氏定氮法(GB/T6432—1994)测定粗蛋白质含量;采用550℃灼烧法(GB/T6438—1992)测定粗灰分含量;采用索氏抽提法(GB/T6433—1994)测定粗脂肪含量。
1.4 数据处理
采用SPSS 17.0软件进行单因素方差分析,用Duncan's多重比较法分析组间差异显著性程度。试验数据均以“平均值±标准差(Mean±SD)”表示。
2 结果
2.1 植酸酶对黄金鲫幼鱼生产性能的影响(见表3)
由表3可知,随着饲料中植酸酶的不断增加,黄金鲫末重、平均增重率有相同的趋势,即先升高后趋于平缓,当植酸酶添加到900 U/kg时,末重和平均增重率比1组显著增加(P<0.05),与对照组相比差异不显著(P>0.05);随着植酸酶添加量的继续增加,末重和平均增重率不再显著增加;随着饲料中植酸酶的不断增加,特定生长率和肥满度有相似的趋势,即先升高后趋于平缓,当植酸酶添加到900 U/kg时,特定生长率和肥满度显著高于1组(P<0.05),与对照组相比差异不显著(P>0.05);随着植酸酶的继续添加,特定生长率和肥满度不再显著增加。

表3 饲料植酸酶水平对黄金鲫生产性能的影响
2.2 植酸酶对黄金鲫幼鱼饲料利用的影响(见表4)
由表4可知,随着饲料中植酸酶的不断增加,饲料效率和蛋白质效率有相同的趋势,即先升高后趋于平缓,当植酸酶添加到900 U/kg时,饲料效率和蛋白质效率显著高于1组,且与对照组相比差异不显著(P>0.05);随着饲料中植酸酶的不断增加,饲料系数有降低的趋势,当植酸酶添加到900 U/kg时,饲料系数与对照组相比差异不显著(P>0.05);随着饲料中植酸酶的不断增加,蛋白质生产率有先升高后趋于平缓的趋势,当植酸酶添加到600 U/kg时,蛋白质生产率显著高于1组(P<0.05),且与对照组相比差异不显著(P>0.05)。
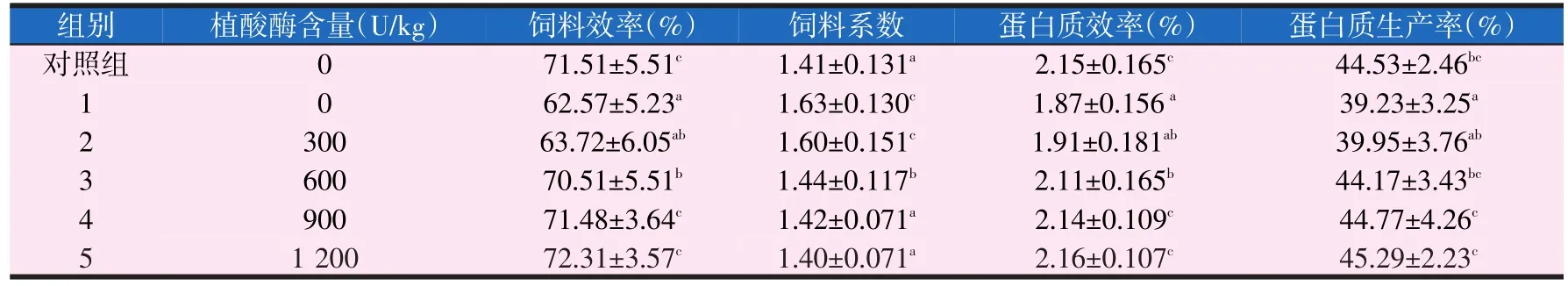
表4 饲料植酸酶水平对黄金鲫饲料利用的影响
2.3 植酸酶对黄金鲫幼鱼体成分的影响(见表5)
由表5可知,各试验组黄金鲫随着饲料中植酸酶的添加,肌肉蛋白质含量有逐渐增加的趋势,当植酸酶添加到900 U/kg时,肌肉蛋白质含量显著高于1组(P<0.05),且与对照组相比差异不显著(P>0.05);随着饲料中植酸酶的添加,肌肉脂肪含量有逐渐降低的趋势,当植酸酶添加到600 U/kg时,肌肉脂肪含量显著低于1组(P<0.05),且与对照组相比差异不显著(P>0.05);各试验组随着饲料中植酸酶的添加,肌肉水分和灰分无显著变化。

表5 饲料植酸酶水平对黄金鲫体成分的影响
3 讨论
3.1 植酸酶对黄金鲫幼鱼生长性能的影响
植酸磷是植物性饲料中磷的主要存在形式,但鱼类的消化系统中缺乏植酸酶来水解植酸磷。因此,若饲料中有效磷的添加较少,会降低鱼类的生长性能和对饲料的利用率[8]。刘行彪等[9]在斑点叉尾鮰基础饲料中添加植酸酶,斑点叉尾鮰的增重率、特定生长率显著提高。张瑞等[10]在含1.1%磷酸二氢钙的建鲤饲料中添加500 U/kg的包被型植酸酶,与只添加1.5%磷酸二氢钙的基础饲料相比,植酸酶组的建鲤特定生长率和增重率高于后者。本试验在黄金鲫饲料中添加600~900 U/kg植酸酶与未添加植酸酶1组相比,提高了试验鱼的增重率和特定生长率。这与孟祥科等[11]对红鳍东方鲀、刘晓侠等[12]对异育银鲫的研究结果相一致。原因为全植物性饲料中添加植酸酶可提高饲料中有效磷的含量,去除了饲料中植酸磷的抗营养作用,使饲料中钙磷比适宜,有利于鱼体的生长。
3.2 植酸酶对黄金鲫幼鱼饲料利用的影响
添加适量的植酸酶能降解植酸磷,将不可利用磷水解为可利用磷,从而提高鱼类饲料的利用率。占海红等[6]在对黄河鲤饲料中添加植酸酶,黄河鲤的饵料系数降低,试验表明,添加适量植酸酶,能提高黄河鲤对饲料的利用率。本试验在黄金鲫饲料中添加900 U/kg植酸酶与1组相比,提高了饲料效率和蛋白质效率,降低了饲料系数。这与周萌等[13]对南美白对虾、舒秋艳等[5]对青鱼、郑涛等[14]对奥尼罗非鱼饲料添加植酸酶的研究结果相一致。其原因为全植物饲料中添加植酸酶,可释放饲料中的植酸磷变为有效磷,从而提高饲料中磷的利用率,降低植酸磷与蛋白质和某些矿物质的络合,间接提高饲料中蛋白质和矿物质的利用率,所以添加植酸酶有利于鱼类对饲料的利用。
3.3 植酸酶对黄金鲫幼鱼体成分的影响
有研究表明,鱼体的肌肉脂肪含量随植酸酶的添加而有所下降。Takeuchi等[15]发现投喂低磷饲料组的鲤体脂肪含量高于投喂高磷饲料的。当饲料中磷缺乏时,抑制了脂肪酸的β-氧化过程,从而使鱼体的脂肪含量维持在较高水平。本试验在黄金鲫饲料中添加900 U/kg植酸酶,与未添加植酸酶1组相比,降低了黄金鲫肌肉脂肪含量。这与Schafer等[16]对鲤鱼、余丰年等[4]对异育银鲫得出了一致的结果。本试验在黄金鲫饲料中添加900 U/kg植酸酶,与未添加植酸酶1组相比,提高了黄金鲫肌肉蛋白质含量。这与曾虹等[17]在鲤鱼基础饲料中添加植酸酶的试验结果相一致。表明鱼类对脂肪利用率的提高起到了节约蛋白质的作用,减少了蛋白质作为能量而消耗,使饲料蛋白质最大限度地转化为鱼体蛋白质,提高了蛋白质的利用率。
4 结论
在本试验条件下,黄金鲫全植物蛋白源饲料中添加植酸酶可达到对照组(适宜的钙磷比)的生长和饲料利用水平,即植酸酶在黄金鲫全植物蛋白源饲料中的适宜添加量为600~900 U/kg。
猜你喜欢
国外畜牧学(猪与禽)(2021年6期)2022-01-11
国外畜牧学(猪与禽)(2021年4期)2021-08-31
国外畜牧学(猪与禽)(2021年6期)2021-03-28
防护林科技(2020年9期)2020-11-09
中南林业调查规划(2018年3期)2018-11-24
防护林科技(2018年10期)2018-11-07
中国畜禽种业(2018年1期)2018-01-18
畜牧兽医科技信息(2015年5期)2015-12-27
中南林业调查规划(2015年3期)2015-12-20
兽医导刊(2015年8期)2015-03-25