饲料磷脂水平对巴丁鱼肝脏脂肪沉积及脂肪代谢酶活性的影响
2015-01-18 12:46麻艳群戴晓玲
饲料工业 2015年6期
■麻艳群 黄 凯 戴晓玲 于 丹 陈 涛
(广西大学动物科技学院,广西南宁 530005)
巴丁鱼(Pangasius sutchi)是我国新兴的一种名贵淡水养殖品种,隶属于鲶形目,科,圆腹属,又称暹罗河鲇、虎头鲨、八珍鱼或巴沙鱼,主要分布在东南亚一带,原产于泰国、马来西亚等国[1],我国近年来开始引种驯养。该鱼无小骨刺,肉质细嫩,口味鲜美。巴丁鱼在生长过程中,腹腔内会逐渐积累三块较大的油脂肪,可达到体重的5%~8%,这一特征在其它鱼类极为罕见。巴丁鱼在越南等地也因此而被称呼为“卡巴沙”——“三块脂肪鱼”。目前国内外对巴丁鱼营养生理方面的研究尚属空白[2-4]。脂肪肝是一种在养殖鱼类中较为常见的营养性疾病。营养不平衡的配合饲料,养殖密度过大,养殖模式不当等因素都有可能导致养殖鱼类营养代谢紊乱,造成肝脏脂肪代谢失调、沉积,脂肪含量升高,肝脏病变,脂肪肝发生。有研究表明,高等动物的肝脏和肌肉均能作为脂肪的贮存位点,体内脂肪合成酶的活性也能很快适应食物的变化,使脂肪代谢得以有效进行[5];但在鱼类,脂肪的贮存位点主要在肝脏,体内脂肪合成酶的活性需要2~3周方可适应食物的变化[6],因此,鱼类比高等动物更容易发生脂肪肝问题。磷脂是指分子中含有磷酸的复合脂,在自然界里广泛存在。有报道表明磷脂对脂肪的转运具有重要的作用,它参与脂肪酸代谢,有助于脂肪的溶解和吸收,降低血浆和肝脏中的脂肪含量[7-8]。本试验以不同水平磷脂饲料饲喂巴丁鱼,研究各组巴丁鱼肝脏脂肪沉积、脂肪合成相关酶活性及组织学变化情况,探讨其脂肪代谢规律,以期通过营养调控方式预防鱼类营养性脂肪肝发生,为磷脂在巴丁鱼人工饲料中的实际生产应用,推广养殖巴丁鱼提供一定参考。
1 材料与方法
1.1 材料
1.1.1 试验用鱼
巴丁鱼(Pangasius sutchi)由广西水产引育种中心提供。驯养7 d后选择规格基本一致、健康活力好的鱼苗900尾[初始体重约(1.45±0.08)g/尾],随机分成5组,每组设3个重复,每重复60尾鱼,每重复的鱼放养于一个水泥池(规格为1.52 m×1 m×1 m)里。
1.1.2 饲料
以豆粕、生物E蛋白、白鱼粉、α-淀粉、米糠等为基础饲料,在该基础上添加不同比例的大豆磷脂(磷脂含量65%),配制成5组饲料(见表1)。各组的磷脂水平分别为0%(PL0组)、1%(PL1组)、2%(PL2组)、3%(PL3组)和4%(PL4组)。
1.2 饲养管理
各组使用相应的饲粮。每天9∶00和17∶00投喂,日投喂量一般为鱼体重的3%,以每次投食在1 h内吃完为宜。饲养周期为56 d。饲养期间于每日清晨投喂前用虹吸法吸出饲料残渣和粪便,换水1/3,水源为曝气后的自来水。试验期间平均水温(27.5±1.0)℃,pH值6.9±0.5,溶解氧(7.10±0.34)mg/l,氨氮(0.03±0.01)mg/l。每日观察记录各池鱼体活动及采食情况。
1.3 测定指标及方法
1.3.1 鱼体成分分析
饲养试验结束后,每重复取10尾鱼,解剖取肝脏,按照AOAC(1984)法测定其粗脂肪含量。计算公式如下:
肝脏脂肪含量(%)=100×肝脏脂肪量/肝重。
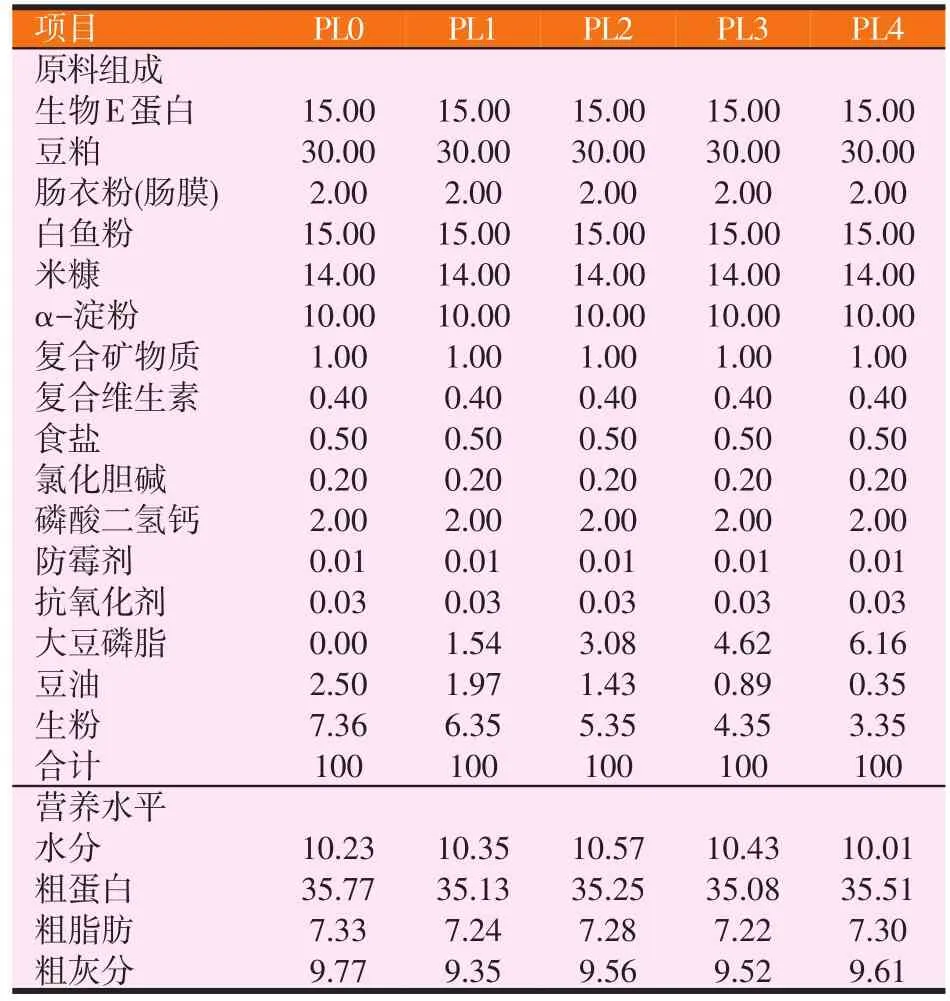
表1 各组饲料组成及营养水平(风干基础)(%)
1.3.2 肝脏组织切片的制备
饲养试验结束后,每重复取3尾鱼解剖,取出肝脏组织固定在Bouin's缓冲液中,做组织切片,HE染色,用以观察肝脏组织的形态学变化。
1.3.3 脂肪合成相关酶酶活测定
比色法测定与脂肪合成相关酶的苹果酸脱氢酶(NADP-malate dehydrogenase,NADP-MDH)和葡萄糖-6-磷酸脱氢酶(Glucose-6-phosphate dehydroge⁃nase,G-6-PDH)的活性。
1.3.3.1 酶液的调制
参照周长海的方法[9]进行部分修改。每重复取鱼10尾,用软布擦干后迅速解剖取出肝脏,用电子天平称取0.5 g左右肝脏样品,倒进匀浆器,加入两倍体积(约1.0 ml)的预冷1 mmol/l EDTA-2Na溶液和0.25 mol/l蔗糖溶液(1∶1),匀浆10 min。匀浆液于4℃9 800 r/min离心10 min,取上清液,4℃ 1 2 000 r/min再离心30 min。取上清液,将其分成两份,一份以1 mmol/l EDTA-2Na溶液和0.25 mol/l蔗糖溶液稀释3倍,用于测定NADP-MDH活性;另一份稀释6倍,用于测定G-6-PDH活性。
酶液蛋白质浓度采用南京建成生物工程研究所的总蛋白定量试剂盒(考马斯亮蓝法)进行测定。
1.3.3.2 NADP-MDH活性测定
依据周长海的方法[9]进行。试管中加入于25℃预温过的0.25 M双甘氨二肽1.2 ml(以2 mol/l NaOH调pH值为7.4)。依次加入0.05 M MnCl20.24 ml;675 μM β-NADP 0.8 ml;0.03 M L-malate(pH值为7.4)0.2 ml;0.25 M 蔗糖 0.51ml;1 mM EDTA-1Na 0.51 ml并混匀,随后加入前期准备好的0.05 ml酶液。震动混匀,测定吸光度(波长340 nm)。
1.3.3.3 G-6-PDH活性测定
依据周长海等(2004)的方法[9]进行。试管中加入于25℃预温过的50 mM三乙醇2.65 ml(以0.1 M NaOH调pH值为7.5)。依次加入酶液0.25 ml;30 mM的β-NADP 0.05 ml;混合后在分光光度仪中培养2 min;加入40 mM G-6-磷酸二钠0.05 ml使反应开始,在340 nm下测定吸光度。空白对照试验都使用去离子水进行。
1.3.3.4 两种酶活计算公式
酶活=[V/(ε×D×P×v)]×[△e/△t]×1 000[nmol/(min×mg]。
式中:V——反应溶液最终总量;
ε——测定时NADP的分子吸光度系数(一般默认ε=6);
D——光路长(1 cm);
P——酶液蛋白质浓度(mg/ml);
v——酶液量(ml);
△e/△t——单位时间内的吸光度变化。
1.4 数据处理与分析
数据均以“平均值±标准误差(X±SE)”表示。用SPSS11.5软件对所有数据进行统计分析,结合Dun⁃can's多重比较法,显著水平采用<0.05。
2 结果
2.1 饲料磷脂对肝脏脂肪含量的影响(见表2)

表2 饲料磷脂对巴丁鱼肝脏脂肪含量的影响(%)
肝脏脂肪含量以PL0组的为最高,达(3.71±0.47)%,显著高于PL2组(P<0.05)(见表2)。
2.2 饲料磷脂对巴丁鱼肝脏组织学的影响
各组肝脏组织切片观察结果见图1。由肝脏组织切片图可以看出,肝脏中部分肝细胞内的细胞核发生偏移,细胞膜模糊,部分细胞核消失,肝细胞浆内含有程度不同的脂肪油滴,出现脂肪空泡,严重程度依次为PL0组>PL1组>PL4组。PL2组和PL3组肝脏细胞形状较规则,其整齐度、分布均匀度较之前述3组的情况要好,脂肪空泡也较少;PL2和PL3组间差异不明显。
2.3 饲料磷脂对巴丁鱼脂质合成相关酶活性的影响(见表3)
由表3可以看出,试验中NADP-MDH活性呈现“U”型变化:对照组的NADP-MDH活性最高(P<0.05),其次为PL1组,PL3组最低,之后随着饲料磷脂添加水平的提高,PL4组的NADP-MDH活性又升高,但仍显著低于对照组(P<0.05)。
肝脏中的G-6-PDH活性也体现出类似于NADPMDH的“U型”变化:对照组的G-6-PDH活性显著高于PL2、PL3、PL4组(P<0.05),PL1组、PL2组、PL3组随着磷脂添加水平的提高逐步降低,到了PL4组,活性又升高。
3 讨论
3.1 饲料磷脂对鱼体脂肪含量的影响
鱼类没有像哺乳类的皮下脂肪层,其脂肪多数蓄积于肝脏、肌肉和腹脂等部位[10],肌肉和腹脂是长期储存部位,而肝脏却是短期储存部位,因此它比肌肉和腹脂更容易受到外源物质的影响[11]。
Meyers[12]认为,在饲料中添加磷脂,能增加载脂蛋白的脂肪运输能力,降低脂肪沉积,从而防治脂肪肝,保护肝脏。Fontagne等[13]通过观察锦鲤(Cyprinus car⁃pio)仔鱼组织发现饲料中若缺乏磷脂则会使仔鱼肠部细胞的脂肪小滴聚积。曹俊明等[14]报道,饲料中添加5%的大豆磷脂,52 d后草鱼(Ctenopharyngodon idellus)肝胰脏脂肪含量会大幅度降低。本试验结果与前人报道相一致,添加了磷脂的4个处理组鱼体肝脏脂肪含量均比对照组有不同程度的降低,这表明饲料中适宜的磷脂水平会促进脂肪代谢,能在一定程度上降低鱼体肝脏中的脂肪沉积。
3.2 饲料磷脂对肝脏组织学的影响
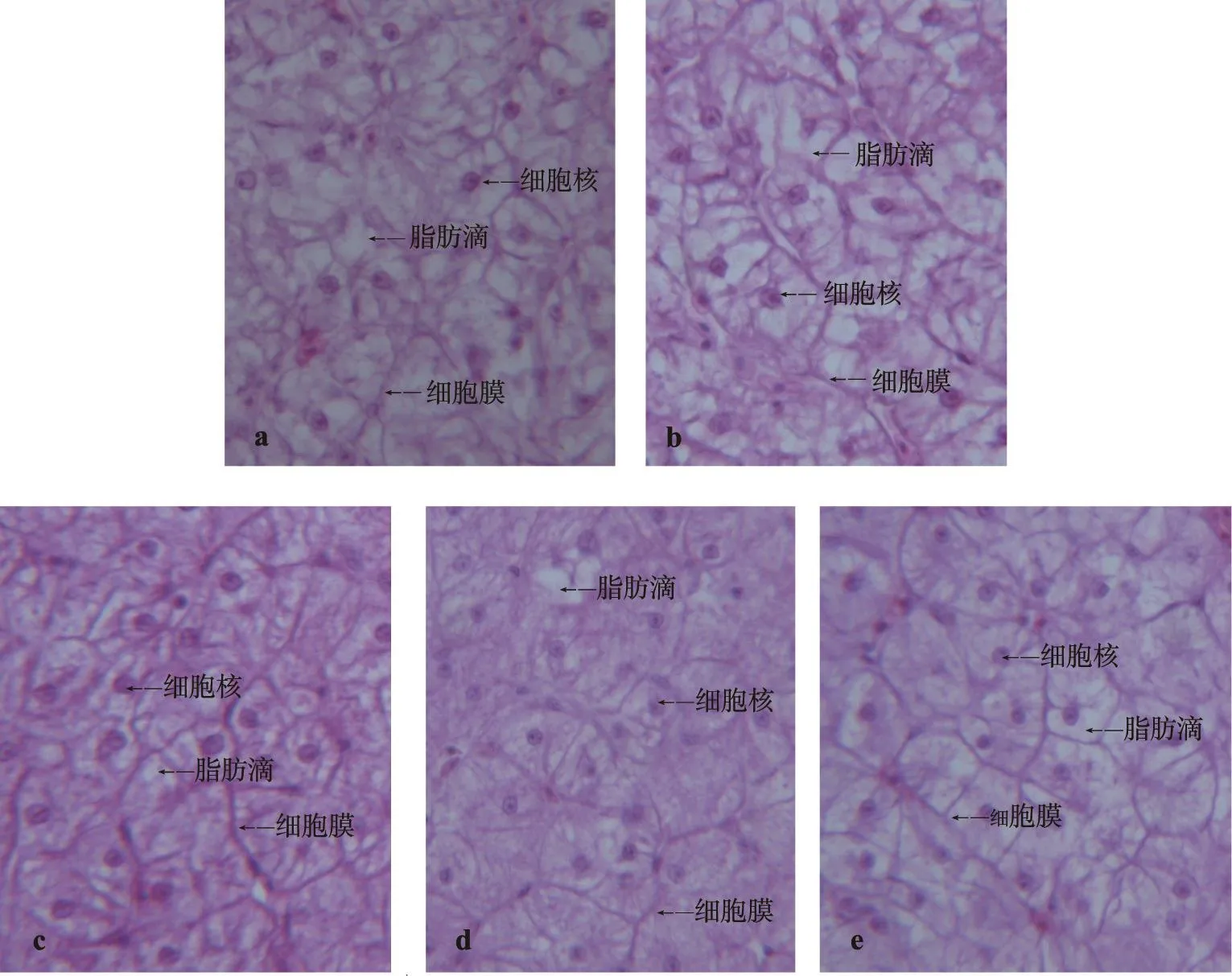
图1 巴丁鱼肝脏组织学观察

表3 饲料磷脂对巴丁鱼脂质合成相关酶活性的影响[nmol/(min·mg)]
有研究表明,磷脂对脂肪的转运具有重要的作用,有助于脂肪的溶解和吸收,降低肝脏脂肪含量。Izquierdo等(2000)发现,给金头鲷(Sparus aurata)仔鱼投喂低水平磷脂饲料,鱼肠道黏膜和肝组织内会有大量脂肪空泡存在,而在饲料中添加大豆磷脂则能显著增加仔鱼肠道脂蛋白的含量,增强脂肪运输能力[15]。Olsen等(2003)报道[16],在饲料中补充大豆磷脂能使虹鳟(Oncorhynchus mykiss)鱼肠上皮细胞形态正常,脂滴积累减少。本试验对肝脏组织学的观察结果与前人的研究报道基本一致,未添加磷脂的对照组出现脂肪空泡的程度最严重,显现轻度脂肪肝症状;其次较严重的是1%磷脂水平组,这可能是因为磷脂能通过直接或间接的作用影响到乳糜微粒脂蛋白的合成,增强脂类的运输能力,而当饲料中缺乏磷脂或磷脂水平不足时,脂肪运输能力下降,脂肪难以从肝脏向肝外组织转运,从而脂肪沉积加重,肝脏组织切片图显示大量脂肪空泡积累。和2%、3%磷脂水平组相比,4%磷脂水平的肝脏也积累了较多的脂肪空泡,这或许是因为饲料中添加4%的磷脂已超过了该阶段巴丁鱼机体的磷脂需要量,因而非但未能有效地提高脂肪运输能力,反而在实质上增加了肝脏脂肪代谢的负担,脂肪也逐渐堆积在肝细胞内。
3.3 饲料磷脂对脂肪代谢酶活性的影响
NADP-MDH是生物体三羧酸循环中最后一个氧化还原酶,可以将氧化型辅酶Ⅱ(NADP)还原成还原型辅酶Ⅱ(NADPH)[17];G-6-PDH是戊糖磷酸代谢途径中的关键酶,能催化G-6-Pi脱氢产生5-磷酸戊糖和NADPH,不管是NADP-MDH还是G-6-PDH,它们催化反应的产物都有NADPH,而NADPH是动物体内脂肪酸合成以及碳链延长的重要辅酶,为脂肪酸合成提供氢原子[18],NADPH在体内的供应状况会直接影响到脂质的合成代谢,因此NADP-MDH与G-6-PDH在肝脏内的活性大小和NADPH的生成量直接相关,进而影响肝脏内脂质的合成代谢[19]。有研究发现,饲喂高胆固醇日粮的小鼠,其肝脏内脂肪酸合成有关的酶活性升高,肝脏因聚积了太多的脂肪而受损[20];在饮食诱导肥胖的小鼠脂肪组织里G-6-PDH活性显著升高[21]。猪背膘中的NADPH生成酶的活性与其胴体脂肪率、背膘厚呈正相关,与瘦肉率呈负相关,因此也有研究人员指出,猪背和膝外层脂肪中NADPMDH和G-6-PDH活性可作为肉用性状早期选种的遗传标记,早期选育瘦肉型猪时,以NADPH生成酶活性为标准强于以背膘厚度为标准[22-23]。
本试验结果中,NADP-MDH和G-6-PDH活性随着饲料磷脂水平的添加均呈现“U”型变化,这有可能是因为在饲料缺乏磷脂或磷脂水平不足时,脂肪在肝脏沉积加重,机体需要增强脂质代谢酶活性去适应脂肪代谢的需要;而当饲料中添加的磷脂含量超过了该阶段机体的需要量,肝脏脂肪代谢负担增加时,肝脏内脂质代谢酶活性也会相应增加。
结合肝脏组织切片图,我们发现,尽管对照组、PL1组的鱼体已增强了脂质合成酶的活性,但其肝脏组织内仍存有较多的脂肪油滴,肝脏脂肪含量大,这或许说明了鱼体对相应饲料日粮做出的主动代偿性增强并不足以真正满足机体脂质代谢的需要,因此在生产养殖中,要预防“营养性脂肪肝”的发生,合理配制人工饲料尤为重要。
4 结论
①在巴丁鱼幼鱼饲料中添加适宜的磷脂(3%),能有效降低鱼体肝脏中的脂肪沉积。
②当饲料缺乏磷脂或磷脂过量时,肝脏脂肪沉积增多,脂肪合成相关酶的活性补偿性升高。
猜你喜欢
中老年保健(2022年4期)2022-11-25
英语世界(2022年9期)2022-10-18
肝博士(2022年3期)2022-06-30
保健医苑(2022年4期)2022-05-05
肝博士(2021年1期)2021-03-29
小哥白尼(趣味科学)(2020年1期)2020-06-16
中成药(2019年12期)2020-01-04
当代水产(2019年9期)2019-10-08
中成药(2018年7期)2018-08-04
中成药(2017年12期)2018-01-19