
甘草查尔酮A抑制小鼠黑色素瘤B16F10细胞糖酵解表型和诱导凋亡
2015-01-02 13:18王艳明刘瑛阎新燕司玲玲高彩霞于丽娜郑秋生
石河子大学学报(自然科学版) 2015年4期
王艳明,刘瑛,阎新燕,司玲玲,高彩霞,于丽娜,郑秋生,3
(1石河子大学药学院/新疆特种植物药资源教育部重点实验室,石河子 832000;2滨州医学院中西医结合学院,烟台 264003;3烟台大学生命科学学院,烟台264005)
恶性肿瘤的发病率和病死率逐年增加,它是导致人类死亡的主要疾病之一。恶性肿瘤具有快速增殖、抵抗凋亡、无限复制潜能、促进血管生成、组织侵袭转移等特征[1],在能量代谢方面,肿瘤细胞主要依赖无氧酵解(糖酵解)代谢,主要表现为葡萄糖摄取量增加、乳酸堆积,这些现象已经被认为是肿瘤细胞典型糖酵解特征[2]。富氧条件下,正常细胞通过有氧酵解的线粒体氧化磷酸化途径产生ATP,满足正常细胞代谢生长的需求;肿瘤细胞则依赖糖酵解以高速率的方式产生更多ATP,满足肿瘤细胞快速生长、增殖和代谢的需求。研究表明:糖酵解不仅能够为肿瘤细胞提供足够的能量和生长所必须的大分子物质(蛋白质、核苷酸、氨基酸等)[3],还能为肿瘤细胞进一步扩大和转移提供有利环境[4]。目前,肿瘤能量代谢已经成为热点研究之一。糖酵解是恶性肿瘤细胞生长、增殖、代谢的能量保障,通过抑制恶性肿瘤细胞特异的糖酵解代谢,是治疗恶性肿瘤的策略之一。
甘草查尔酮A是一种酚类查尔酮化合物,大量分布于甘草根茎中,具有抗寄生物[5]、抗菌[6]、抗肿瘤[7]等作用。现阶段甘草查尔酮A对肿瘤细胞能量代谢方面的研究较少,因此,本研究通过探讨甘草查尔酮A抑制B16F10细胞糖酵解表型和诱导细胞凋亡的作用,为今后靶向抑制黑色素瘤能量代谢研究提供实验依据。
1 材料与方法
1.1 材料
1.1.1 细胞株
小鼠黑色素瘤细胞 B16F10购买于中国典型培养物保藏中心,本实验室液氮冷冻保存。
1.1.2 药品及试剂
甘草查尔酮A(成都普瑞法生物科技有限公司,纯度为 98%,批号:14022606);DMEM培养基(GIBCO公司,批号:1342967);胎牛血清(杭州四季青生物工程材料有限公司,批号:140130);磺基罗丹明B(Sulforhodamine B,SRB,Sigma,批号:3520421);二甲基亚砜(DMSO,Sigma,批号:51596BPV)Giemsa干粉(北京拜尔迪生物技术有限公司,批号:R246403);葡萄糖测定试剂盒(上海荣盛生物药业有限公司,批号:20141105147);乳酸测定试剂盒(南京建成生物工程研究所,批号:20141226);ATP含量测定试剂盒(南京建成生物工程研究所,批号:20141222);AnnexinV/FITC凋亡检测试剂盒(南京凯基生物科技发展有限公司,批号:20150104)。
1.1.3 仪器
CO2细胞培养箱(美国 Thermo公司,Thermo 3131);多功能酶标仪(美国 Thermo公司,Thermo 3001);倒置荧光显微镜(德国 ZEISS公,MIC00266,);流式细胞仪(美国BD公司,FACSCalibur);MH-2微量振荡器(海门市其林贝尔仪器制造有限公司)。
1.2 方法
1.2.1 细胞培养
取生长状态良好细胞B16F10,用含10%胎牛血清、链霉素(100μg/mL)、庆大霉素(100μg/mL)的DMEM培养液(pH=7.5),在37℃、5%CO2饱合湿度的培养箱中培养,取对数生长期细胞进行各项实验。
1.2.2 甘草查尔酮A母液配制
称取甘草查尔酮A 17 mg放入0.5 mL离心管中,加入DMSO 500μL将其溶解,即为100μmol/L甘草查尔酮A母液。临用前用DMEM培养基稀释到所需浓度。
1.2.3 SRB法测定细胞增殖抑制率
取对数生长期B16F10细胞,计数后用完全培养基制成细胞悬液,以细胞数0.8×104/mL接种于96孔板中(100μL/孔),培养过夜,然后加不同浓度的甘草查尔酮A完全培养基100μL,使终浓度分别为 0、15、30、45、60、75、90μmol/L,继续分别培养24 h和48 h,一定时间后,每孔加入50%(质量/体积)的三氯乙酸(TCA)50μL固定细胞,然后在4℃冰箱中放置1 h。培养板各孔用去离子水洗涤5遍,完全去除多余TCA,室温晾干96孔板,每孔加0.4% 的SRB 100μL,在室温下放置20 min,弃去各孔内液体后,用1% 乙酸洗涤5遍,去除未结合的染料,室温晾干,用DMSO(二甲基亚砜)150 μL溶解,微量振荡器上振荡8 min使结晶物充分溶解,酶标仪在515 nm波长下检测每孔吸光度(OD)值。并按公式计算药物对细胞增殖的抑制率:

1.2.4 细胞培养液中葡糖糖含量检测
取对数期细胞,消化,接种于6孔板,待细胞贴壁状态良好时,加甘草查尔酮A,使孔内浓度分别为 0、30、45和 60μmol/L,将其放入 37℃,5%CO2饱合湿度的培养箱中培养24 h后,取出6孔板,吸出培养液于离心管,3000 r/min离心5 min,留取上清培养液备用。按照普利莱基因技术试剂盒说明书操作后,在多功能酶标仪550 nm处测定吸光度值。
1.2.5 细胞培养液中乳酸含量测定
取对数期细胞,消化,接种于6孔板,待细胞贴壁状态良好时,加甘草查尔酮A,使孔内浓度分别为0、30、45和 60μmol/L,将其放入 37℃,5%CO2饱合湿度的培养箱中培养24 h后,取出6孔板,吸出上清培养液于离心管,3000 r/min离心5 min,留取上清培养液备用。然后按照南京建成生物工程研究所说明书操作后,在多功能酶标仪530 nm处测定吸光度值。
1.2.6 细胞内ATP含量测定
取对数期细胞,消化,接种于细胞培养瓶中,待细胞贴壁状态良好时,加甘草查尔酮A,使终浓度分别为 0、30、45、60μmol/L,将其放入 37℃,5%CO2饱合湿度的培养箱中培养24 h后,消化,离心,收集细胞,将收集好细胞加入300μL热双蒸水,放在热水浴(90-100℃)中匀浆破碎后,在沸水浴中加热10 min,取出混匀抽提1 min后,按照南京建成生物工程研究所说明书操作后,在多功能酶标仪636 nm处测定吸光度值。
1.2.7 Giemsa染色观察细胞形态
取对数生长期细胞,消化,离心后,以1.5×105铺6孔板(2 mL/孔),待细胞贴壁状态良好后,用终浓度为0,30,45和60μmol/L甘草查尔酮A处理B16F10细胞24 h,吸弃培养液,PBS洗2遍,每孔加固定液(甲醇∶冰醋酸=3∶1)1 mL,固定30 min后,吸弃固定液,PBS洗2遍,每孔加Giemsa染色液1.5 mL,染色 15 min,吸弃染液,PBS洗 2-3遍,吸弃PBS,晾干,镜下观察并拍照。
1.2.8 Hoechst33258染色观察细胞形态
取对数生长期细胞,消化,离心后,以1.5×105铺6孔板(2 mL/孔),待细胞贴壁状态良好,用终浓度为0、30、45、60μmol/L甘草查尔酮A处 理B16F10细胞24 h后,吸弃培养液,PBS洗2遍,用固定液(甲醇∶冰醋酸 =3∶1)固定10 min,用 PBS洗2遍,晾干,加适量Hoechst33258染色液染色10 min,PBS洗去未结合的荧光染料洗净,镜下观察并拍照。
1.2.9 流式细胞术检测细胞凋亡率
细胞经药物作用24 h后,3000 r/min离心5 min,收集细胞,预冷PBS洗涤细胞1次,调整细胞数为1×105/mL,加入Binding buffer悬浮细胞液500μL,加入Annexin V-FITC 5μL,混匀后加入碘化丙啶(propidium iodide,PI)5μL,再次混匀。室温,避光,反应 10 min,在 1 h内进行流式细胞仪检测。激发波长 Ex=488 nm;发射波长Em=530 nm。
1.3.0 统计学分析
所有实验均设3个平行组或重复3次,结果以±S表示,以t检验(Origin7.5统计软件)进行组间比较。以P<0.05为差异具有统计学意义,P<0.01为差异具有显著统计学意义。
2 结果与分析
2.1 甘草查尔酮A明显抑制B16F10细胞生长
甘草查尔酮A以不同浓度分别作用于B16F10细胞24或48 h,随药物浓度升高,细胞抑制率明显增高,呈浓度依赖性和时间依赖性(图1)。24和48 h的 IC50分别为(47.31±2.06)μmol/L和(36.88±1.2)μmol/L。
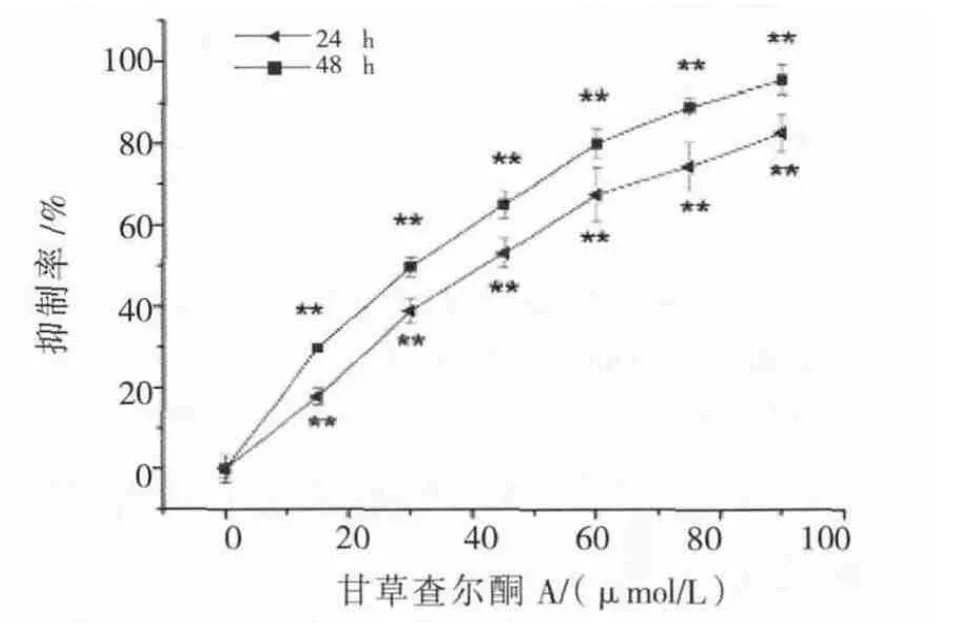
图1 甘草查尔酮A对B16F10细胞增殖影响Fig.1 Effect of Licochalcone A on the proliferation of B16F10 cells
2.2 甘草查尔酮A抑制B16F10细胞摄取葡萄糖
肿瘤细胞通过大量摄取葡萄糖获取足够的能量和生物大分子物质,以满足自身快速增殖的需求。甘草查尔酮A能浓度依赖性的抑制B16F10细胞摄取葡萄糖(图2)。
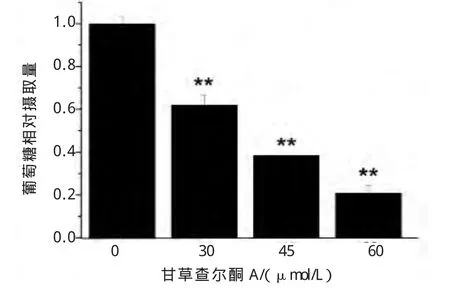
图2 甘草查尔酮A对B16F10细胞葡萄糖摄取量的影响Fig.2 Effect of Licochalcone A on the glucose uptake of B16F10 cells
2.3 甘草查尔酮A抑制B16F10细胞乳酸生成
葡萄糖被摄入肿瘤细胞后,经过多种糖酵解激酶的催化反应转化为最终产物乳酸,乳酸的大量堆积有利于肿瘤细胞的浸润和转移[4]。不同浓度甘草查尔酮 A(0、30、45、60μmol/L)作用于 B16F10细胞24 h后,明显减少B16F10细胞培养液中乳酸含量,并呈浓度依赖性(图3)。
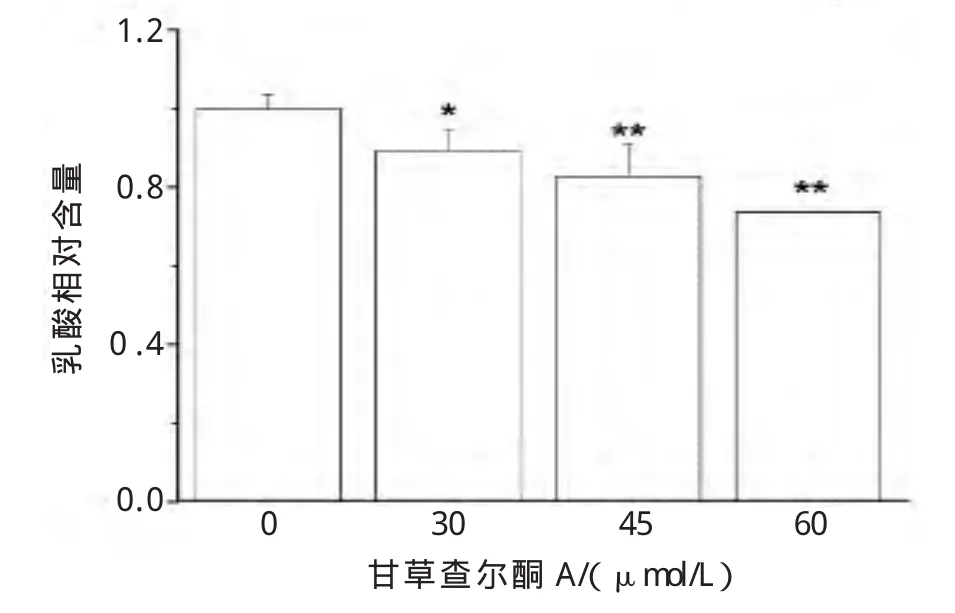
图3 甘草查尔酮A对B16F10细胞培养液中乳酸生成的影响Fig.3 Effect of Licochalcone A on the extracellular lactic acid production of B16F10 cells
2.4 甘草查尔酮A抑制B16F10细胞内ATP的生成
正常细胞通过有氧酵解的线粒体氧化磷酸化途径产生ATP,而肿瘤细胞依赖糖酵解高速率产生ATP,维持细胞活跃的能量代谢。从图4可知,甘草查尔酮A能浓度依赖性抑制B16F10细胞内ATP的生成。
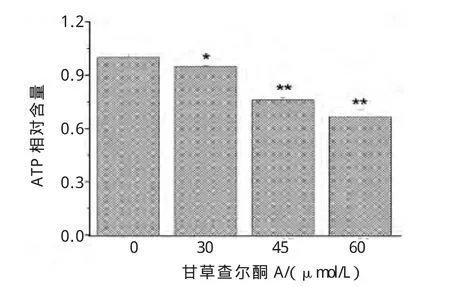
图4 甘草查尔酮A对B16F10细胞内ATP生成的影响Fig.4 Effect of Licochalcone A on the intracellular ATP production of B16F10 cells
2.5 甘草查尔酮A对B16F10细胞形态影响
甘草查尔酮A作用B16F10细胞24 h后,Giemsa染色观察细胞形态,对照组细胞贴壁状态良好,呈不规则梭形,随着药物浓度的增加(30、45、60μmol/L),细胞数量逐渐减少,高浓度组细胞(60μmol/L)呈固缩圆形结构(图5)。Hoechst 33258染色进一步观察细胞形态,如图6所示,正常组细胞染色均匀,细胞大而饱满,随着药物浓度的增加(30、45、60μmol/L),细胞出现典型凋亡特征:细胞体积逐渐变小,胞核皱缩,碎裂,呈现致密颗粒状荧光。
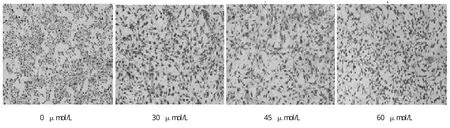
图5 Giem sa染色法观察甘草查尔酮A对B16F10细胞形态影响(×200)Fig.5 Effect of Licochalcone A on morphological changes in B16F10 cells observed by Giemsa staining(×200)

图6 Hoechst 33258染色法观察甘草查尔酮A对B16F10细胞凋亡形态影响(×400)Fig.6 Effect of Licochalcone A on apoptotic morphological changes in B16F10 cells observed by Hoechst 33258 staining(×400)
2.6 甘草查尔酮A诱导B16F10细胞凋亡
为进一步明确甘草查尔酮A对B16F10细胞凋亡的影响,流式细胞仪检测细胞凋亡率。图7细胞流式散点图中,A表示右上象限(细胞晚期凋亡),B表示右下象限(细胞早期凋亡)。甘草查尔酮A能诱导B16F10细胞发生早期凋亡和晚期凋亡(图7、图 8)。
与对照组相比,高浓度组(45和60μmol/L)早期凋亡(P<0.01)具有显著性差异;且高浓度组(45和60μmol/L)晚期凋亡也(P<0.05,P<0.01)具有差异性,其中高浓度组(60μmol/L)细胞晚期凋亡率达38.1%±1.90%(图 8)。
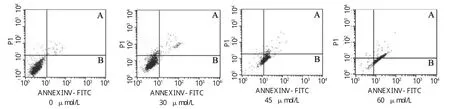
图7 细胞流式散点图Fig.7 Flow cytometry scatter diagram
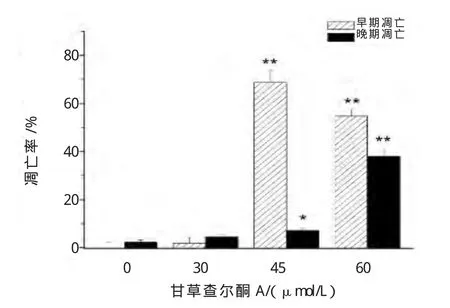
图8 甘草查尔酮A对B16F10细胞凋亡率影响Fig.8 Effect of Licochalcone A on apoptotic rates of B16F10 cells
3 讨论
恶性黑色素瘤是由皮肤和其他器官黑素细胞产生的肿瘤,其发病率与总体死亡率依旧呈现逐年上升的趋势[8],给人类健康带来巨大威胁。深入认识黑色素瘤发生和发展的分子机制对于开辟新的治疗黑色素瘤方法有重要意义。近年来,探索通过靶向糖酵解代谢途径治疗恶性肿瘤的策略正备受关注[9]。
糖酵解被认为是肿瘤细胞特征性表型之一[10]。肿瘤细胞依赖糖酵解获得足够的能量和必须生物大分子物质,以满足细胞快速增殖,无限复制的需求。葡萄糖摄取量快速增加和乳酸大量堆积是肿瘤细胞典型糖酵解特征。肿瘤细胞能通过细胞膜上的葡萄糖转录蛋白摄入葡萄糖,随后大部分的葡萄糖经过多种糖酵解激酶的催化,生成ATP和大分子物质,这些物质的生成有利于肿瘤细胞快速生长、增殖、转移和浸润。另外,乳酸的大量积累会使肿瘤细胞微环境酸化,这种酸性环境有利于肿瘤细胞的生存[4]。本实验室前期高通量测序结果显示:糖酵解与黑色素瘤细胞的发生和发展密切相关,且黄酮类化合物异甘草素能显著抑制黑色素瘤B16F0细胞糖代谢,本实验中甘草查尔酮A是与异甘草素分子结构极为相似的黄酮类化合物;B16F10细胞和B16F0细胞都是小鼠黑色素瘤细胞,两者相比,B16F10细胞恶性程度、转移和浸润能力更高,因此推测甘草查尔酮A可能对B16F10细胞糖代谢有抑制作用。结果显示,甘草查尔酮A能显著抑制B16F10细胞增殖和细胞糖酵解表型(细胞葡萄糖摄取量、细胞外乳酸含量和细胞内ATP含量明显减少)。
肿瘤细胞糖酵解代谢与凋亡有密切联系。研究表明:肿瘤细胞糖酵解代谢受到抑制时,还伴随细胞凋亡的发生[11-13]。为明确甘草查尔酮A抑制B16F10细胞糖酵解过程中是否伴随细胞凋亡发生,首先通过Giemsa和Hoechst 33258染色法观察细胞形态,其中Gimesa染色结果显示,随着药物浓度的增加,细胞数量逐渐减少,高浓度组细胞(60μmol/L)呈固缩圆形状;Hoechst 33258染色结果显示:加药组细胞出现典型凋亡特征:细胞核体积逐渐缩小,胞核皱缩,碎裂,呈现致密颗粒状荧光状;随后通过流式细胞仪检测细胞凋亡情况。结果显示,甘草查尔酮A能浓度依赖性的诱导B16F10细胞凋亡,与对照组相比,高浓度组细胞(45和60μmol/L)的早期凋亡率具有显著性差异(P<0.01)。
综上所述,甘草查尔酮A可能是通过抑制B16F10细胞摄取葡萄糖,减少乳酸和ATP生成,使细胞无法获得足够的能量和细胞增殖必须的生物大分子物质,削弱B16F10细胞糖酵解代谢,从而抑制B16F10细胞增殖。由于肿瘤细胞长期处于“饥饿”状态,就会引起应激性反应—凋亡的发生,肿瘤细胞可以通过诱导部分细胞凋亡,减少能量消耗,增加自身生存的几率。这些结果表明,甘草查尔酮A能通过抑制B16F10细胞糖酵解表型和诱导细胞凋亡实现抗黑色素瘤作用。
[1]Hanahan D,Weinberg R A.Hallmarks of cancer:the next generation[J].Cell,2000,100(1):57-70.
[2]Warburg O.On the origin of cancer cells[J].Science,1956,123(3191):309-314.
[3]Zhao Y,Liu H,Riker A I,et al.Emerging metabolic targets in cancer therapy[J].Front Biosci(Landmark Ed),2011,16:1844-1860.
[4]Gatenby R A,Gillies R J.Why do cancers have high aerobic glycolysis[J].NatRev Cancer,2004,4(11):891-899.
[5]Xiao X Y,Hao M,Yang X Y,et al.Licochalcone A inhibits growth of gastric cancer cells by arresting cell cycle progression and inducing apoptosis[J].Cancer Lett,2011,302(1):69-75.
[6]Yo Y T,Shieh G S,Hsu K F,et al.Licorice and licochalcone-a induce autophagy in LNCaP prostate cancer cells by suppression of Bcl-2 expression and the mTOR pathway[J].J Agric Food Chem,2009,57(18):8266-8273.
[7]Kim J K,Shin E K,Park J H,et al.Antitumor and antimetastatic effects of licochalcone A in mouse models[J].JMol Med(Berl),2010,88(8):829-838.
[8]Thompson J F,Scolyer R A,Kefford R F.Cutaneous melanoma[J].Lancet,2005,365(9460):687-701.
[9]Garber K.Energy deregulation:licensling tumors to grow[J].Science,2006,312(5777):1158-1159.
[10]Zheng J.Energy metabolism of cancer:Glycolysis versus oxidative phosphorylation(Review)[J].Oncol Lett,2012,4(6):1151-1157.
[11]徐高峰,白晓斌,王茂德,等.脑胶质瘤细胞糖酵解表型特征及对细胞增殖与凋亡的影响[J].南方医科大学学报,2013,33(3):406-411.Xu G F,Bai X B,Wang M D,et al.Characterization of glycolytic phenotype of SHG44 human glioma cells under hypoxic conditions and its association with cell proliferation and apoptosis[J].J South Med Univ,2013,33(3):406-411.
[12]Wang Z,Wang D,Han S,et al.Bioactivity-guided identification and cell signaling technology to delineate the lactate dehydrogenase A inhibition effects of spatholobus suberectus on breast cancer[J].PloS one,2013,8(2):56631.
[13]Zhao Y,Liu H,Riker A I,et al.Emerging metabolic targets in cancer therapy[J].Front Biosci:Landmark Ed,2011,16(3):1844-1866.
猜你喜欢
中南医学科学杂志(2022年6期)2022-12-04
中国循证心血管医学杂志(2020年11期)2020-01-08
天然产物研究与开发(2018年9期)2018-10-08
中华老年口腔医学杂志(2016年4期)2017-01-15
中国医学装备(2016年6期)2016-12-01
西南国防医药(2016年6期)2016-12-01
癌症进展(2016年8期)2016-08-22
癌变·畸变·突变(2016年5期)2016-08-22
小猕猴学习画刊(2016年9期)2016-05-14
癌症进展(2016年10期)2016-03-20
