
赤皮青冈ISSR-PCR反应体系的建立与优化
2014-12-28 06:38朱品红李志辉杨模华
中南林业科技大学学报 2014年6期
朱品红,李志辉,杨模华
(中南林业科技大学 林学院, 湖南 长沙 410004)
赤皮青冈ISSR-PCR反应体系的建立与优化
朱品红,李志辉,杨模华
(中南林业科技大学 林学院, 湖南 长沙 410004)
为建立适宜于赤皮青冈ISSR分析的扩增体系,以赤皮青冈叶片基因组DNA为材料,采用单因素和正交设计相结合的方法系统地测试模板DNA、Mg2+、引物、磷酸碱基脱氧核苷(dNTPs)浓度,Taq DNA聚合酶用量和退火温度这6个因素对ISSR-PCR反应结果的影响。综合分析表明:优化后的最佳反应体系为20 µL反应总体系中,含20 ng模板DNA,2.5 mmoL/L Mg2+,0.2 mmoL/L dNTPs,1.0 U Taq DNA聚合酶,0.2 µmoL/L引物;UBC 808号作引物最佳退火温度为59.3℃。这一优化体系的建立为进一步开展赤皮青冈遗传多样性的ISSR分析奠定了基础。
赤皮青冈;ISSR-PCR;正交设计;单因素试验;优化
简单重复序列区间扩增多态( inter-simple sequence repeat,ISSR)分子标记技术是近几年来发展起来的一种新型分子标记法,由Zietkiew ica等[1]于1994年在SSR (Simple Sequence Repeat)基础上创建的,综合了SSR和RAPD技术的优点,具有稳定性高、重复性好、操作方便、成本低廉等特点。因此近年来已广泛应用于遗传作图、遗传多样性分析、基因定位和种质资源鉴定[2-8]等方面。
赤皮青冈Cyclobalanopsis gilva (Bl.) Oerst.属壳斗科青冈属,常绿乔木,树皮暗赤褐色,又名红椆、赤皮稠,是珍贵的硬木树种。主要产自我国福建、湖南、浙江、贵州、台湾等省。适应性强,生于海拔300~1 500 m的山地,较耐干旱脊薄,能在丘陵酸性红壤与石灰岩发育而成的钙质土壤生长[9]。但近年来由于人类的影响,赤皮青冈天然种群逐渐枯竭,仅保存下少量的古木大树[10]。目前国内外对赤皮青冈的研究主要集中在地理分布[11],造林技术[9],理化特性[12-14]等方面。而关于ISSR标记技术的研究尚未见报道。本研究采用单因素试验与正交设计相结合的方法,建立了适合赤皮青冈的ISSR-PCR最佳反应体系,为ISSR分子标记技术应用于赤皮青冈的种质资源鉴定、遗传变异分析、系统进化等研究奠定了基础。
1 材料与方法
1.1 材 料
赤皮青冈种子来源于湖南省张家界市永定区,于2012年在汨罗科技示范园育苗,2013年4月将新鲜叶片置于低温储藏箱中带回实验室,洗净、晾干,-70℃冰箱中保存备用。
1.2 主要试剂及仪器
参照加拿大哥伦比亚大学UBC公司提供的ISSR引物序列,由上海英骏生物技术有限公司合成。经初步筛选,确定引物808号即(AG)8C作为本次反应体系优化试验的固定引物。扩增反应在GeneAmp PCR System 9700扩增仪上进行,试验所用Taq DNA聚合酶、Mg2+、dNTPs购自TINGEN公司,标准分子量( Marker)DL2000购自东盛公司。
1.3 基因组DNA的提取与检测
采用OMEGA公司生产的试剂盒对赤皮青冈DNA进行提取,方法参照说明书。λDNA测定提取液浓度,溴化乙锭(EB)染色,0.7%的琼脂糖凝胶电泳检测。
1.4 试验方法
1.4.1 单因素试验优化反应体系
根据预备实验结果,对影响ISSR-PCR反应体系的6个因素,分别设置了不同的梯度水平,见表1。通过查阅相关文献确定最初的反应体系为:40 ng的模板 DNA,1.4 µL 的 Mg2+,0.8 µL 引物,0.3 µL 的 dNTPs,0.3 µL 的 Taq DNA 聚 合 酶,10×Buffer缓冲液2.0 µL;PCR初步反应程序为94℃预变性5 min;94℃变性30 s,58℃退火45 s,72℃延伸2 min,共循环35次;最后72℃延伸7 min;4℃保温。
1.4.2 正交设计试验优化反应体系
对模板DNA、引物、Mg2+、dNTPs、Taq酶5个因素进行正交设计,因素水平参考单因素试验,依据PCR重复2次的扩增结果进行直观分析。
1.4.3 ISSR-PCR产物的检测
ISSR-PCR产物用EB染色,1.4%的琼脂糖凝胶电泳检测。5 V/cm恒定电压下电泳约1.5 h,然后用Bio-Rad紫外凝胶成像仪拍照保存。
2 结果与分析
2.1 DNA提取结果
赤皮青冈叶片基因组DNA的凝胶电泳检测结果如图1所示。图中主带明亮清晰,无明显拖带,点样空基本干净,无杂质,说明蛋白质、多糖、酚类等基本去除,满足ISSR分子标记实验的要求。

图1 赤皮青冈叶片基因组DNA电泳结果Fig.1 Electrophoresis pattern of genomic DNA of the leaves of C. gilva
2.2 单因素试验结果分析
2.2.1 模板DNA浓度对ISSR-PCR反应的影响
适宜的模板DNA浓度是保证ISSR-PCR良好扩增效果的一个重要因素,DNA模板浓度过低,扩增不稳定甚至无法扩增;而浓度过高,又相应增加了非特异性扩增产物[15]。此次单因素试验设置了7个模板DNA浓度梯度,见图2,从图可以看出:赤皮青冈ISSR-PCR体系中,对DNA模板浓度的许可范围较大。在20 µL的反应总体系中所加的DNA模板体积(浓度50 ng/ µL)大于1.4 µL时扩增条带较弱,不清晰。当加入的体积在0.4~1.2 µL之间时扩增条带稳定、清晰。
2.2.2 引物浓度对ISSR-PCR反应的影响
引物与模板DNA的互补程度决定了PCR产物的特异性,是PCR反应的关键,浓度过高可能引起模板与引物的错配,PCR反应特异性下降,还会增大形成引物二聚体的几率;浓度过低则不能进行有效扩增。从图3可以看出:随着引物浓度的增高,扩增条带数由多到少,反应的稳定性也逐渐降低。当引物(浓度10 µmoL/L)加入量为0.4 µL时扩增条带清晰,数量较多,特异性强。加入量为0.6、0.8、1.0 µL时条带清晰度下降,特异性差。因此选用引物体积0.4 µL作为ISSR-PCR反应体系中最适加入量。
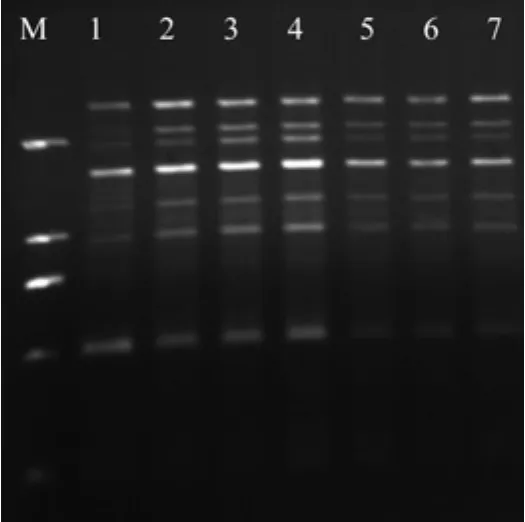
图2 不同水平模板DNA对ISSR扩增的影响Fig.2 Effect of different DNA template levels on ISSR amplif i cation
2.2.3 Mg2+浓度对ISSR-PCR反应的影响
Mg2+浓度影响了引物与模板双链的解链。退火温度、Taq DNA聚合酶的活性和产物的特异性也都受Mg2+浓度的影响[16-17]。在一定浓度范围内,随着Mg2+浓度升高,PCR扩增的特异性降低,然而浓度过低则影响PCR扩增产量,甚至不能扩增出条带。从图4可以看出20 μL的反应总体系中Mg2+(浓度50 mmol/L)加入量为0.6 μL时无扩增条带。加入量为1.8 μL时条带背景模糊,主次带
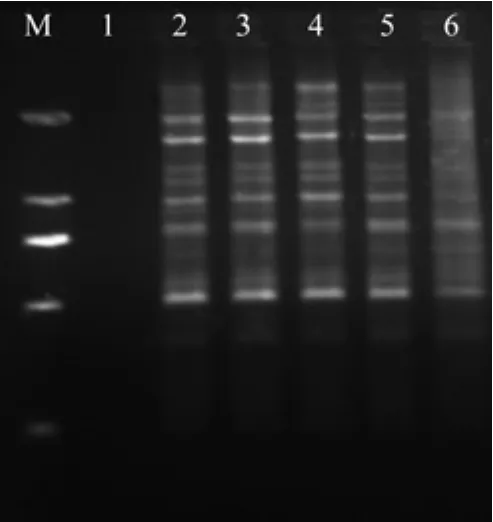
图4 不同水平Mg2+对ISSR扩增的影响Fig.4 Effect of different Mg2+ levels on ISSR amplif i cation
2.2.5 Taq DNA聚合酶用量对ISSR-PCR反应的影响
在ISSR-PCR反应中,Taq DNA聚合酶的活性与浓度对扩增反应有着重要的影响,浓度过低会导致扩增产物不足或者不能扩增,浓度过高会产生弥散现象,非特异性扩增产物增多。值得注意的是不同厂家生产的酶,或同一厂家不同批次的酶对不分明。在加入0.8~1.4 μL时扩增条带较多,其中加入1.0 μL时最清晰稳定,故确定1.0 μL作为ISSR-PCR反应体系中Mg2+最适加入量。
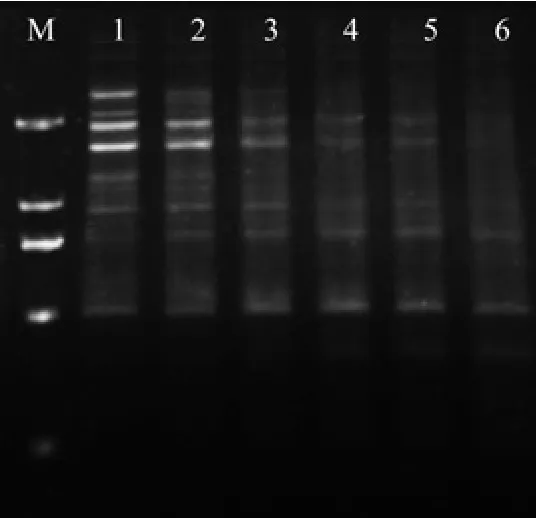
图3 不同水平引物对ISSR扩增的影响Fig.3 Effect of different primer levels on ISSR amplif i cation
2.2.4 dNTPs浓度对ISSR-PCR反应的影响
dNTPs是PCR反应中DNA序列扩增的原料,浓度过低会降低PCR产物的产量,过高即能产生错误掺入,又会与体系中游离的Mg2+结合,从而减少了Mg2+含量。如图5所示,当20 μL 的反应总体系中dNTPs(浓度10 mmol/L)加入量达到1.0 μL时已无扩增产物出现。加入0.8 μL时扩增条带较少,加入0.2,0.6 μL时条带清晰可见,而加入0.4 μL时主次带分明,清晰度最高。因此,本实验确定0.4 μL作为ISSR-PCR反应体系中dNTPs最适加入量。反应效果也存在差异[18]。如图6所示在20 µL 的反应总体系中Taq DNA聚合酶(浓度5 U/ µL)加入量为0.1 µL时,扩增条带弱,不能辨析,加入0.2µL时条带较强,背景清晰。当加入量逐渐增加时拖尾现象开始严重,达到0.5 µL时无扩增产物出现。因此选用0.2 μL作为ISSR-PCR反应体系中Taq酶最适加入量。
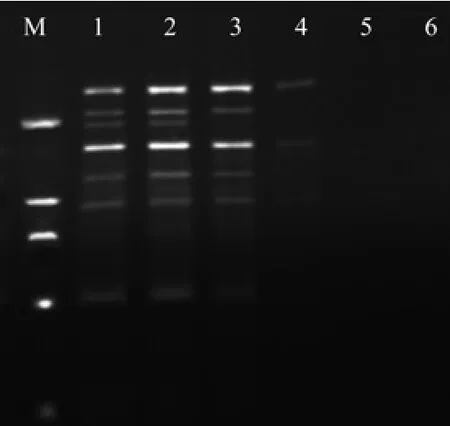
图5 不同水平dNTPs对ISSR扩增的影响Fig. 5 Effect of different dNTPs levels on ISSR amplif i cation
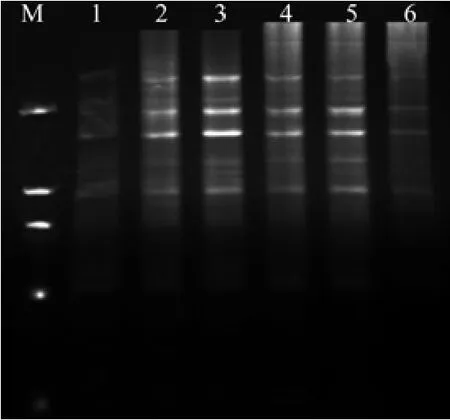
图6 不同水平Taq DNA聚合酶对ISSR扩增的影响Fig. 6 Effect of different Taq DNA polymerase levels on ISSR amplif i cation
2.2.6 最优ISSR-PCR退火温度的筛选
由于作为ISSR-PCR标记的引物长度、碱基组成、GC碱基含量及浓度等各不相同,各个引物的Tm值也有所不同。一般的温度范围是45~65℃[19]。退火温度偏高则引物与模板难结合,条带较少,背景弱;退火温度偏低则弱带较多,特异性差。若扩增结果相似则选择较高退火温度,以提高引物与模板结合的特异性。本试验在PCR梯度扩增仪上设置退火温度从48℃到62℃,自动生成8个温度梯度,见图7。由图可以看出退火温度为48~56.8℃时,由于温度偏低,导致背景模糊,杂带较多,特异性差;退火温度高于61.0℃时扩增受到抑制,电泳条带少。当温度为59.3℃时扩增产物最多,条带清晰。因此,本实验确定引物808的最佳退火温度为59.3℃。

图7 不同水平退火温度对ISSR扩增的影响Fig. 7 Effect of different annealing temperature levels on ISSR amplif i cation
2.3 正交试验结果分析
根据单因素试验结果,对模板DNA、引物、Mg2+、dNTPs及Taq DNA聚合酶5个因素在4个水平上进行正交试验设计,建立L16(45)正交表(表2)。各因素水平分别为DNA模板:0.4、0.6、0.8、1.2 µL;引物:0.4、0.6、0.8、1.0 µL;Mg2+:0.8、1.0、1.2、1.4 µL;dNTPs:0.2、0.4、0.6、0.8 µL;Taq 酶:0.15、0.2、0.3、0.4 µL。
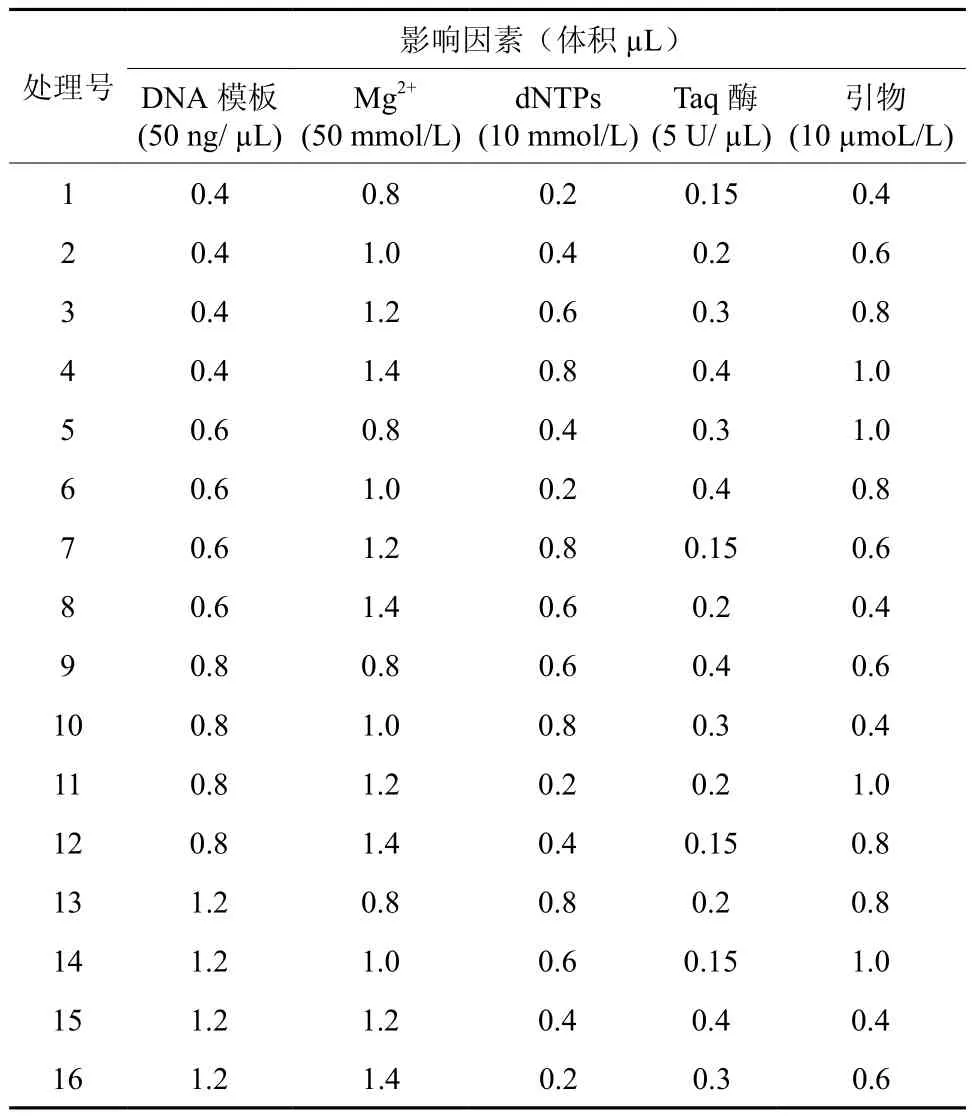
表2 赤皮青冈ISSR反应正交试验设计表L16(45)Table 2 ISSR reaction orthogonal design L16(45) of C.gilva
参考何正文等[20]的方法,根据L16(45)正交试验PCR扩增产物电泳结果(图8),分析谱带的强弱、数量、背景清晰度及杂带多少,对其进行直观分析,打分。最优的记为16分,最差的记为1分。则本次试验16个处理的分数依次为:8、16、15、11、13、6、1、14、4、12、2、3、9、5、7、10。根据分数进行直观分析(表3)得出各因素对赤皮青冈ISSR-PCR反应体系的影响程度即极差值R,R值越大说明该因素对反应的影响越显著。而Ki值反映了各个因素同一水平下的试验值之和,ki值则反映了各个因素水平对反应的影响效果,ki值越大说明该水平对扩增反应的影响越明显。由极差值R反映出各因素对赤皮青冈ISSR-PCR扩增体系的影响程度由高到低依次是:Taq DNA聚合酶、模板DNA、Mg2+、dNTPs、引物。由ki值得出理论最佳反应体系是:模板DNA(50 ng/ µL)0.4 µL、Mg2+(50 mmol/L)1.0 µL、dNTPs(10 mmol/L)0.4 µL、Taq酶(5 U/µL)0.3 µL、引物(10µmoL/L)0.4 µL。各因素的最佳水平组合并没在16个处理中出现,但是与分值最高的2号处理接近。所以,在利用正交试验设计优化反应体系时,可以不经过直观分析,直接根据电泳结果也能获得较优的反应体系[21]。
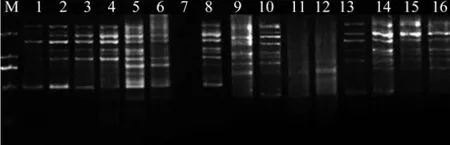
图8 正交试验PCR产物电泳结果Fig. 8 Electrophoresis of orthogonal design PCR products

表3 正交试验数据分析Table 3 Data analysis of orthogonal tests
通过正交试验确定模板DNA最佳含量为0.4µL。Mg2+、引物和dNTPs在单因素试验和正交试验结果中最佳含量相同,分别是1.0 µL、0.4 µL和0.4 µL。正交试验中Taq酶的最佳含量为0.3 µL比单因素试验的最佳值高出0.1 µL,考虑到经济原因,选择0.2 µL作为PCR反应的最佳含量。综合单因素试验与正交设计试验结果,可以得到赤皮青冈ISSR-PCR的最佳反应体系,见表4。
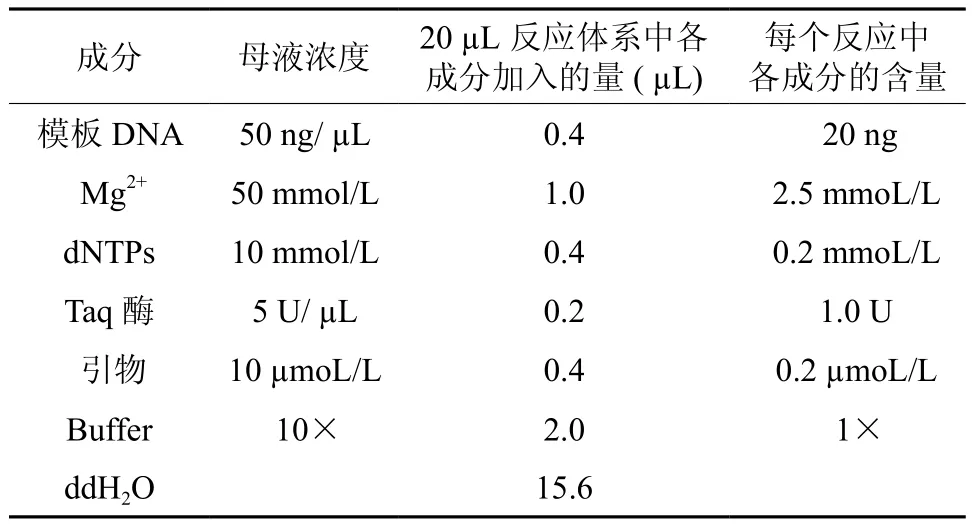
表4 赤皮青冈ISSR-PCR最佳反应体系Table 4 The most suitable ISSR-PCR system for C.gilva
2.4 ISSR-PCR反应体系的稳定性检测
图9是在本研究得出的最佳反应体系条件下以808号作引物,退火温度59.3℃,扩增18个样品的PCR电泳结果。从图可以看出,优化后的体系均能扩增出清晰度高、多态性好的条带,说明本研究确立的赤皮青冈ISSR-PCR反应体系是稳定可靠的。

图9 808号引物在优化后的体系下扩增18个样品的PCR结果Fig.9 The PCR results of 18 specimens with peimer 808 and optimized reaction system
3 小结与讨论
因为退火温度会随着物种和引物种类的不同而发生改变,因此在进行某一物种的ISSR-PCR体系优化时,对不同的引物退火温度应重新进行筛选。本研究确定在进行赤皮青冈ISSR-PCR扩增时808号作引物的最佳退火温度为59.3℃。在实验的整个过程中还应注意尽可能使用同一厂家的药品和试剂,以便保证分析结果的稳定性。
前人在进行ISSR-PCR反应体系优化时有的只考虑各因素不同水平对试验结果的影响,忽略了因素之间的交互作用,有的只采用正交设计试验方法,使结果的评价带有一定的主观性,降低了最佳反应水平的的可靠程度。本研究则综合运用两种试验方法,在一定程度上避免了各自的局限性。通过此次研究确立的赤皮青冈ISSR-PCR最佳反应体系,将为赤皮青冈及其近缘种的种质资源鉴定及遗传多样性的分析研究工作奠定基础。
[1] Zietkiew icz E, Rafalski A, Labuda D. Genome fingerprinting by simple sequence repeat (SSR) anchored polymerase chain reaction amplif i cation[J]. Genomics,1994,20:176-183.
[2] Zhou J L, Jie Y C, Jiang Y B,et al.Development of simple sequence repeats(SSR) markers of ram ieand comparison of SSR and inter-ISSR marker systems[J].Progress in Natural Science,2005, 15(2):137-142.
[3] Vijiayan K,Srivastava P P, AwasthiA K. Analysis of phylogenetic relationship among five mulberry (Morus) species using molecular markers[J].Genome, 2004, 47: 439-448.
[4] 沈永宝, 施季森, 赵洪亮. 利用ISSR DNA标记鉴定主要银杏栽培品种[J].林业科学, 2005, 41(1): 202-204.
[5] 孙 洪, 程 静, 詹克慧, 等. ISSR标记技术及其在作物遗传育种中的应用[J].分子植物育种, 2005, 3(1): 123-127.
[6] 黄福平, 梁月荣, 陆建良, 等. 应用RAPD和ISSR分子标记构建茶树回交1代部分遗传图谱[J]. 茶叶科学, 2006, 26(3):171-176.
[7] 邢建宏, 陈存及, 张国防, 等. 樟树几种化学类型及近缘种的ISSR分析[J]. 福建林业科技, 2008, 35(4): 101-104.
[6] Wolfe A D, Xiang Q Y, Kephart S R. Assessing hybridization in natural population of Penstemon (Scrophulariaceae) using hypervariable inter-simple sequence repeat (ISSR) bands [J].Molecular Ecology, 1998, 7(9): 1107-1125.
[9] 罗仲春, 徐玉书. 赤皮青冈造林应用技术研究[J]. 中南林业调查规划, 1995, 14(3): 23-25.
[10] 祁承经, 汤庚国. 树木学(南方本)(第2版)[M]. 北京: 中国林业出版社, 2005.
[11] 谢 健. 赤皮青冈种群的生存分析[J]. 福建林学院学报, 2011,31(3): 254-256.
[12] 吴丽君, 李志辉, 赵嫦妮. 赤皮青冈光合作用的日变化研究[J]. 江西林业科技, 2011, (5):1-3.
[13] 景美清, 李志辉, 杨模华, 等. 赤皮青冈种子质量与萌发特性研究[J]. 中国农学通报, 2012, 28(34): 27-30.
[14] 赵嫦妮, 徐德禄, 李志辉. 配方施肥对赤皮青冈容器苗生长的影响[J]. 中南林业科技大学学报, 2013, 33(5): 22-25.
[15] 杨 华, 宋绪忠, 尹光天,等.黄藤ISSR反应体系的条件优化[J]. 福建林学院学报.2006,26(2):152-155.
[16] 邹喻苹, 葛 颂, 王晓东.系统与进化植物学中的分子标记[M]. 北京: 科学出版社, 2001.
[17] 张志红,谈凤笑,何航航,等.红树植物海漆ISSR条件的优化[J]. 中山大学学报: 自然科学版, 2004,43(2): 63-66.
[18] 朱柏芳,朱 笃,邓荣根,等.穗花杉ISSR引物反应条件的优化与筛选[J].植物研究,2006,26(3):318-322.
[19] Reddym P, SarilN, Sidd iq E A. Inter simple sequence repeat(ISSR) polymorphism and its application in plant breeding [J].Euphytica, 2002, 128(1): 9-17.
[20] 何正文,刘运生,陈立华,等.正交设计直观分析法优化PCR条件[J].湖南医科大学学报,1998,23(4):403-404.
[21] 胡尚力, 徐刚标, 梁 艳, 等. 伯乐树cpDNA-PCR反应体系的优化与引物筛选[J].中南林业科技大学学报,2013,33(7): 67-71.
Establishment and optimization of Cyclobalanopsis gilva ISSR-PCR reaction system
ZHU Pin-hong, LI Zhi-hui, YANG Mo-hua
(School of Forestry, Central South University of Forestry and Technology, Changsha 410004, Hunan, China)
∶ In order to establish the optimal system of ISSR-PCR for Cyclobalanopsis gilva, with the genomic DNA of C.gilva leaves as material, single factor test and orthogonal experiment design were used to systematically test six parameters of ISSR-PCR which include template DNA, Mg2+, primers, dNTPs concentration, dose of Taq DNA polymerase and annealing temperature.Integrated analysis implies that the optimal reaction system of ISSR (20 μL) is:20 ng template DNA, 2.5 mmoL/L Mg2+, 0.2 mmoL/L dNTPs, 0.2 µmoL/L primers and 1.0 U Taq DNA polymerase, if UBC808 as the primers in ISSR-PCR that the optimized annealing temperature is 59.3℃.The establishment of the optimal system lays an important foundation for using ISSR technology in research C.gilva genetic diversity in future.
∶ Cyclobalanopsis gilva; ISSR-PCR; orthogonal design; single factor tests; optimization
S723.7
A
1673-923X(2014)06-0061-05
2013-09-04
“十二五”国家科技支撑项目“楸树和赤皮青冈珍贵用材林定向培育技术研究与示范”(2012BAD21B03)
朱品红(1987-),女,河南焦作人,硕士研究生,从事林木定向培育研究
李志辉(1957-),男,湖南安化人,教授,博士生导师,主要从事森林培育教学和研究工作;
E-mail:lzh1957@126.com
[本文编校:吴 彬]
猜你喜欢
广东药科大学学报(2022年3期)2023-01-04
生物学通报(2022年1期)2022-11-22
计算机应用与软件(2022年6期)2022-07-12
自然灾害学报(2022年2期)2022-05-10
中国种业(2021年11期)2021-11-25
中国学校体育(2021年10期)2021-04-26
生物学教学(2019年3期)2019-03-22
报刊荟萃(上)(2017年7期)2017-07-16
魅力中国(2016年29期)2017-05-27
江苏农业科学(2016年6期)2016-07-25
