
海拉尔地区苜蓿生产性能及影响因子研究
2014-12-21 03:25徐丽君辛晓平杨桂霞
草原与草坪 2014年4期
徐丽君,辛晓平,杨桂霞
(呼伦贝尔国家野外站/中国农业科学院农业资源与农业区划研究所,北京 100081)
苜蓿作为牧草之王,其光合作用的研究一直被受国内外众多学者的高度关注。近年来,针对不同栽培条件、不同品种和不同生育期苜蓿单叶及整株净光合速率(Pn)、蒸腾速率(Tr)和植物水分利用率(WUE)之间的关系[1-4],及其对植物叶片的气孔导度(Gs)和水分亏缺(VPD)等自身因子及气温(Ta)、空气相对湿度(RH)和光合有效辐射(PAR)等环境因子间的响应[5,6],光合作用与产量间的关系[7,8]、农艺措施[9,10]对光合作用的影响等方面进行了大量的研究。随着研究的深入,研究领域已逐步延伸到羧化效率、量子产量、叶绿素荧光等光合能力(自然或特定CO2浓度水平和最适温、湿度条件下的光饱和光合速率)及某些生化指标的日变化规律[11-15]等方面。
呼伦贝尔草原是我国温带草甸草原分布最集中、最具代表性的地区,是我国目前原生植被保存最好、景观生态类型和生物多样性最丰富的草原生态系统类型[16]。随着人为、放牧的干扰,天然草地生产力日益下降,这对大力发展畜牧业的海拉尔市经济发展提出了严峻的考验,为保证畜牧业健康良好的发展,确保饲草料供应充足,人工草地扮演了重要角色。通过研究比较在海拉尔地区大面积种植的苜蓿品种的光合特性和生产性能,以期筛选出适宜在海拉尔地区种植的苜蓿品种,同时探讨影响苜蓿生产性能的因子,为该地区苜蓿栽培和利用提供依据。
1 研究区概况与研究方法
1.1 研究区概况
试验地选在呼伦贝尔生态实验站,地处N 49°06′~49°32′,E 119°32′~120°35′,研究区域内水热条件较好,属于温带大陆性季风气候,海拉尔河与伊敏河交汇于此,水资源丰富,无霜期110d,年平均气温-2℃ ,日照时数年均2 807h,太阳辐射年总量平均为5161 MJ/m2。土壤以黑钙土为主。
1.2 研究方法
1.2.1 供试材料 选择生长2年龄黄花苜蓿(Medicagosativacv.Yellow flower)、杂花苜蓿(M.varia)、肇东苜蓿(M.sativacv.Zhaodong)、龙牧801(M.sativacv.Longmu 801)的苜蓿人工草地为研究对象。种植采用条播,行距为40cm,播量均为15kg/hm。
试验地苜蓿5月初返青,9月中旬收获,每年收获两茬,分别在6月下旬和8月中旬,鲜草产量8t/hm2。在苜蓿处于开花期进行测定,植株平均高度55cm、日均气温20℃。
1.2.2 测定方法 2010年7月28~29日连续2d(晴天),选取苜蓿健康植株上部第3片完全伸展的健康叶片,利用Li-6400便携式光合仪测定其光合生理生态指标,包括叶片的光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间 CO2浓度(Ci)等生理因子以及大气温度(Ta)、相对湿度(RH)等气象指标。每项指标重复测量3次。测定时间全天为7∶00~19∶00,每隔2 h测定1次。叶片瞬时水分利用效率(WUE)由式(1)求算。

1.2.3 植株产量因子测定 株高由随机选择生长中等植株从地面测量至植株顶端,重复10次;植株距地2/3处的宽度为宽幅;随机选择生长健康植株,数根颈上的分枝数,重复10次为单株分枝数;测定各品种的产草量,样方2.0m2,重复3次。产量以单位面积的干草产量来计;每次测产时将各品种约100g鲜样阴干,将茎、叶分离,测定。
茎叶比=干样中茎量/干样叶量。
鲜干比=干样重量/鲜样叶量
取新鲜植物叶片,擦净组织表面污物,剪碎(去掉中脉),混匀,参照文献[17]常规方法测定叶绿素。
1.3 统计分析
数据采用Excel、SAS 8.0软件处理,并进行LSD检验和方差分析。同时分别对光合因子与环境因子、产量因子与环境因子之间进行相关分析,最终通过主成分分析确定影响苜蓿生产性能的因素。
2 结果与分析
2.1 4种苜蓿产量因子
分析4个苜蓿品种产量因子,结果表明(表1),株高以龙牧801最低,为44.33cm,黄花苜蓿最高,为62.33cm,其他2品种介于53.67~60.67cm;苜蓿品种变化规律与株高基本一致,龙牧801宽幅最小,杂花苜蓿最宽,是龙牧801的1.1倍;单株分枝数以龙牧801分枝数最少,为34.67枝/株,以肇东苜蓿枝条数最多,达48.33枝/株,高出龙牧801品种39.40%;单株干重、单株茎干重、单株叶干重和产量的变化规律一致,龙牧801最低,黄花苜蓿最高,4种苜蓿单株干重、单株茎干重、单株叶干重和产量分别为18.05~46.01g,8.56~25.53g,8.43~18.59 g和6.73~14.15t/hm2;茎叶比龙牧801最低,为0.82,说明龙牧801单位叶量丰富,黄花苜蓿最高,为1.10,其余2品种茎叶比为0.95~1.02;鲜干比以杂花苜蓿最高,黄花苜蓿最低,杂花苜蓿高出黄花苜蓿26.71%;叶绿素含量,杂花苜蓿最高,达42.56 mg/g FW,龙牧801最低,为37.54mg/g FW。综合比较4种苜蓿形态学指标,与本土品种黄花苜蓿相比,杂花苜蓿与黄花苜蓿的各项指标数值上相近,且略高于黄花苜蓿,龙牧801的各项指标数值上均为最低。
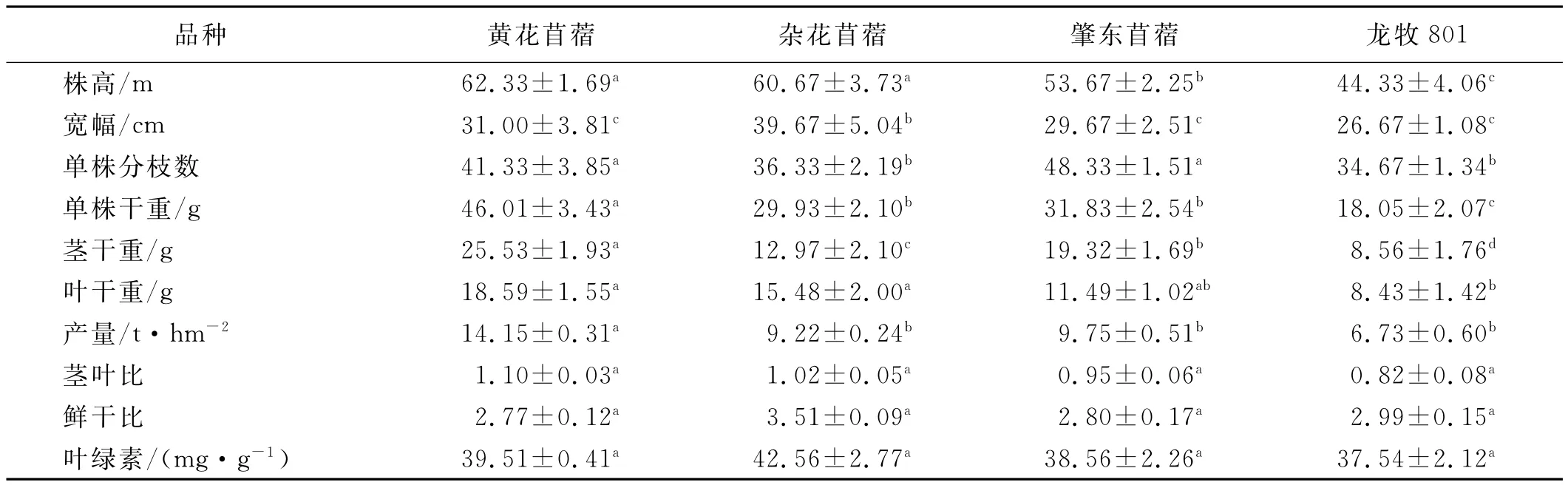
表1 4种苜蓿产量诸因子Table1 Yield afectingfactors of 4alfalfa varieties
2.2 光合特性
2.2.1 光合速率 从4种苜蓿光合速率日动态变化规律可以看出(图1-a),光合速率日动态变化曲线均呈双峰曲线,出现最大值的时间均分别在上午10∶00和下午的2∶00。比较光合速率日动态平均值,结果显示龙牧801>杂花苜蓿>黄花苜蓿>肇东苜蓿,其相应的光合速率值分别为23.70,23.05,22.35和21.28 μmol/(cm2·s)CO2。
2.2.2 蒸腾速率 分析4个苜蓿品种蒸腾速率日动态的变化规律可以看出(图1-b),4个苜蓿品种蒸腾速率日动态变化规律差异性较大,黄花苜蓿、龙牧801和肇东苜蓿蒸腾速率最大值出现在早上6∶00,杂花苜蓿蒸腾速率最大值出现在14∶00。比较蒸腾速率日平均值,结果显示龙牧801>黄花苜蓿>杂花苜蓿>肇东苜蓿,其相应的蒸腾速率值分别为12.40,12.01,10.67和10.52mmol/(m2·s)H2O。
2.2.3 水分利用效率(WUE)由图1-c可知,肇东苜蓿和龙牧801水分利用效率变化规律基本一致,在16∶00达到最大值,分别为4.97和4.92μmol/mmol CO2,黄花苜蓿和杂花苜蓿在14∶00达到全天最大值,分别为3.68和2.96μmol/mmol。综合比较4种苜蓿水分利用效率的高低,得出龙牧801>肇东苜蓿>黄花苜蓿>杂花苜蓿。

2.2.4 胞间二氧化碳浓度(Ci)从图1-d中可以看出,4种苜蓿胞间二氧化碳浓度的变化规律基本一致,在6∶00达到全天的最高值,14∶00出现全天最低值,品种间差异不显著(P>0.05)。比较4种苜蓿胞间二氧化碳浓度的平均数值大小,总体变化杂花苜蓿>黄花苜蓿>肇东苜蓿>龙牧801,相应的数值分别为253.67,245.67,227.33和219.67μmol/mol。
2.2.5 气孔导度(Gs)图1-e表明,不同苜蓿品种气孔导度总体变化规律基本一致,均呈“多峰”曲线,第1高峰值出现在8∶00(黄花苜蓿除外,第一峰值出现在10∶00),第2高峰值均出现在18∶00,第1低峰值出现在12∶00。综合比较4种苜蓿气孔导度的平均数值大小,总体为杂花苜蓿>黄花苜蓿>龙牧801>肇东苜蓿。
2.3 植物的光合、蒸腾性能与环境因子的关系
分析光合速率、蒸腾速率与环境因子的相关性,结果表明(表2),黄花苜蓿Pn与蒸腾速率、气孔导度、胞间二氧化碳浓度、气温和叶绿素呈显著正相关(P<0.05),相关系数r均在0.850以上,与水分利用效率、相对湿度、有效辐射显著负相关(P<0.05),相关系数r介于0.923~0.995,与水分亏缺相关性不大;杂花苜蓿Pn与叶绿素极显著正相关(P<0.01),与其他因子不相关或负相关;肇东苜蓿Pn与水分利用效率、相对湿度、有效辐射、叶绿素显著正相关(P<0.05),相关系数0.849以上,与胞间二氧化碳浓度、相对湿度负相关;龙牧801的Pn与蒸腾速率、气孔导度、水分亏缺、叶绿素正相关,与相对湿度显著负相关(P<0.05,r=-0.766~-0.878)。蒸腾速率与各因子的相关性基本与光合速率的相同,其中,肇东苜蓿除外,肇东苜蓿Tr与气孔导度、叶绿素正相关,与水分亏缺显著负相关(P<0.05)。
比较分析,4种苜蓿Pn和Tr与各因子的相关性基本一致,即气孔导度、叶绿素对苜蓿光合、蒸腾影响较大,即受植物自身内在因素的影响较大,水分利用效率、相对湿度和气温等外界环境因素(水、温)对苜蓿光合、蒸腾作用具有一定的抑制作用。

表2 不同苜蓿净光合速率、蒸腾速率与相关因子间的相关分析Table2 Correlation analysis among Pn,Trand its affecting factors for tested alfalfa varieties
2.4 产量与植株形态因子间的相关性
分析苜蓿产量与植株形态因子的相关性,结果表明(表3),黄花苜蓿产量与叶干重、单株干重、宽幅、茎干重和鲜干比的相关系数较高(r=0.853~0.999),与茎叶比成负相关(r=-0.877);杂花苜蓿产量与叶干重、宽幅、单株分枝数、单株干重、鲜干比成正相关(r=0.955~0.999),而与茎叶比呈显著负相关(P<0.05,r=-0.998);肇东苜蓿产量与宽幅显著正相关(P<0.05,r=0.999),与株高、单株干重和茎干重相关系数相对较低(r=0.616~0.751);龙牧801产量因子与单株干重、叶干重、茎干重呈显著正相关(P<0.05,r>0.950),而与株高呈显著负相关(P<0.05,r=-0.999)。综合分析4个苜蓿品种产量与各因子间相关性,可知产量与叶干重、单株干重、宽幅和茎干重相关性较好,与茎叶比、株高的相关性较差。

表3 不同苜蓿产量与相关因子间的相关分析Table3 Correlation analysis between yield and its affecting factors for different alfalfa varieties
2.5 产量与光合、蒸腾及环境因子的相关性
从表4可以看出,黄花苜蓿产量与相对湿度、有效辐射呈正相关(P<0.05,r=0.969~0.995),与光合速率、蒸腾速率、胞间二氧化碳浓度、气温呈负相关;杂花苜蓿产量与有效辐射、气孔导度、水分亏缺呈显著负相关(P<0.05,r>0.996);肇东苜蓿产量与蒸腾速率、气孔导度显著正相关(P<0.05,r>0.998),与水分亏缺负相关;龙牧801产量因子与有效辐射呈显著正相关(P<0.05,r=0.952),而与胞间二氧化碳浓度、气温呈负相关。

表4 不同苜蓿品种产量与相关因子分析Table4 Correlation analysis between yield and its affecting factors for different alfalfa varieties
2.6 影响产量因子主成分分析
通过对4个苜蓿品种的19个构成因子进行主成分分析,结果得出19个特征根和特征向量,按最小特征根>1的原则,从中选出3个特征根,其累积率达85%。主成分1对变异的累积贡献率为40.9%,其中,x2和x13所占的权重较大,说明株高和有效辐射对苜蓿产量的影响最大;主成分2对变异的累积贡献率为23.6%,其中,x1,x3,x5,x6和x7所占的权重较大,说明产量、宽幅、单株干重、茎干重和叶干重之间存在相互促进关系;主成分3对变异的累积贡献率为13.52%,其中,x9所占的权重最大,即为鲜干比;主成分4对变异的累积贡献率为11.54%,其中,x12和x14所占的权重最大,分别代表水分利用效率和胞间二氧化碳浓度。从结果可以看出,影响苜蓿产量主要是植株的形态指标,即植株的生长状况,生理因子在其中占有的比例较小。

表5 影响因子主成分分析Table5 Equation and the proportion of main characters
3 讨论与结论
3.1 光合特性
植物的生理特性主要是植物的光合,蒸腾特性以及水分利用率。研究植物的光合特性,有利于了解植物对该光能的利用效率,阐明植物光合的生态学特征[18]。比较4种苜蓿日平均Pn,得出杂花苜蓿与龙牧801日平均Pn较高,日平均Pn反映了植物光合能力的大小,说明杂花苜蓿与龙牧801光合能力较强。试验中,4种苜蓿的Pn日变化表现出“双峰”型曲线,与前人研究结果中典型“双峰”型或“单峰”型曲线的结论基本一致[4-7,19],蒸腾速率日变化规律与时晓霞等[20]结果不一致,究其原因,可能与品种、外界环境条件和测定时间有关。而且,植物光合能力的强弱也因物种遗传特性差异而不同,因而苜蓿日光合能力因品种间的差异而呈现不同的峰型。
水分利用率是表征植物经济用水的能力。不同苜蓿品种的水分利用率不同。气孔导度大的品种适合该地区的环境,水分不是其限制因子,气孔导度最大的品种的水分利用率不一定是最高[18]。所以,水分利用率是评价不同苜蓿品种的一个重要指标,但是必须和气孔导度、光合速率结合起来考虑才能得到较为客观的结果[21,22]。试验研究表明,杂花苜蓿日平均Pn和气孔导度均较高,日平均Tr,WUE与其他3种苜蓿相比相对较低,综合比较来看4种苜蓿,杂花苜蓿经济用水能力相对较强,说明杂花苜蓿较适宜在海拉尔地区种植。
3.2 影响产量要素
试验研究结果显示苜蓿产草量与光合速率、蒸腾速率、气孔导度、胞间二氧化碳浓度呈显著负相关,与水分利用效率正相关,分析其中的原因,可能是由于测量方法造成的误差,试验在进行光合测定的过程中,没有进行相应的产量测定,而是在进行植株形态指标测定的过程中进行相应产量的测定,测产的植株与进行光合测定选取的植株不一致,这一原因可能是导致产量与光合因子间相关性分析结果产生差异的一个原因,在接下来的研究工作还需对该结论进行进一步的验证。
株高、根颈粗、单株重、叶面积、分枝数等因素均是构成苜蓿生物产量的重要因子,是苜蓿高产的主要性状。诸多研究表明苜蓿茎粗、茎长和分枝数与产量极显著相关。杨青川等[22]研究报道指出,分枝数对产量影响较大,叶片数和节间数对产量的影响较小。韩路等[23]应用灰色关联度分析了31个国内外优良苜蓿种质资源的生产特性,结果表明众多性状中茎粗对产量的贡献最大,其次为抗病能力,株高,分枝数。试验研究表明苜蓿产量与单株干重、茎、叶干重和植株宽幅呈显著正相关,与株高、分枝数、茎叶比呈正相关,但相关性不大,这一试验结果与前人报道基本一致。同时采用主成分分析法分析了4种苜蓿的影响产量的19个因素,结果表明株高、产量、宽幅、单株干重、茎干重、叶干重、鲜干比和有效辐射等指标能较好的反映和预测苜蓿的产量,光合因子对反映苜蓿产量的贡献率较低,所以在选择苜蓿产量构成要素的过程中,建议优先考虑植株的形态因子,之后再考虑光合因子。
(1)4种苜蓿光合特性存在一定的差异性。Pn,Tr,WUE,Ci和Gs日变化规律不完全相同,出现峰值的时间也存在一定差异性。总体变化规律是Pn出现高峰期在10∶00和14∶00,Tr高峰值出现在6∶00和14∶00,WUE最大值出现在14∶00~16∶00,Ci日变化为多峰型,最大值出现在6∶00,Gs最大值分别出现在8∶00和18∶00。
(2)4种苜蓿Pn和Tr与各因子的相关性基本一致,即Pn和Tr与气孔导度、叶绿素相关性较高,呈显著正相关(P<0.05),与水分利用效率、相对湿度和气温呈负相关(P<0.05)。产量与光合因子、环境因子相关性较低。
(3)通过主成分分析,株高、有效辐射、产量、宽幅、单株干重、茎干重、叶干重和鲜干比能够较好的反映和预测苜蓿产量。
(4)杂花苜蓿较适宜在内蒙古海拉尔地区种植。
[1] 苏加楷,张文淑.中国牧草品种审定工作的进展[J].草原与草坪,2002(3):3-5.
[2] 王刚,孙广玉.不同紫花苜蓿品种光合能力的比较[J].东北林业大学学报,2007,35(4):19-21.
[3] 王运涛,于林清,萨仁.苜蓿抗寒性研究进展[J].草原与草坪,2012,32(3):91-94.
[4] 刘玉华,贾志宽,史纪安,等.旱作条件下不同苜蓿品种光合作用的日变化[J].生态学报,2006,26(5):1468-1477.
[5] 陈玉香,周道玮,张玉芬.玉米、苜蓿间作的产草量及光合作用[J].草地学报,2004,12(2):107-112.
[6] 黄顶,张英俊,韩建国.白三叶、多年生黑麦草和鸭茅光合生理生态特性研究[J].中国农业大学学报,2006,11(2):44-48.
[7] 孟林,张国芳,赵风华,等.不同水分条件下紫花苜蓿光合与蒸腾特性研究[M].中国草学会六届二次会议暨国际学术研讨会论文集,2004:540-544.
[8] 赵金梅,周禾,郭继承.不同水分胁迫对紫花苜蓿分枝期光合性能的影响[J].中国草地学报,2007,29(2):41-44.
[9] 卜崇峰,刘国彬,赵姚阳.黄土丘陵沟壑区狼牙刺的光合特征及其水分利用效率[J].西北植物学报,2004,24(13):2189-2195.
[10] 高景慧,张颖,郭维,分枝期3个紫花苜蓿品种光合蒸腾日变化与相关因子的关系分析[J].西北农林科技大学学报(自然科学版),2007,35(10):29-34.
[11] 许大全,徐宝基.气孔限制在植物叶片光合诱导中的作用[J].植物生理学报,1989,15(3):275-280.
[12] 刘彦,曹连莆,丁荣荣,等.奎屯垦区苜蓿品种筛选试验[J].草原与草坪,2010,30(5):77-82.
[13] Lindsay W B,Aleida H W,Megan H R,etal.Water relations and adaptations to increasing water deficitin three perennial legumes,Medicago sativa,Dorycnium hirsutum and Dorycnium rectum[J].Plant Soil,2007,290:231-243.
[14] Sheridan K P.Intraspecific Variation in Apparent Net Photosynthesis of Seven Crop Species[M].Univpark:Pennsylvania State University,1966.
[15] 唐华俊,辛晓平,杨桂霞,等.现代数字草业理论与技术研究进展及展望[J].中国草地学报,2009(4):1-8.
[16] 上海植物生理学会.植物生理学实验手册[M].上海:上海科学技术出版社,2009:3.
[17] 温方.紫花苜蓿不同品种生产性能及其光合特性研究[D].北京:中国农业科学院,2007:5.
[18] 刘玉华,贾志宽,韩清芳,等.不同苜蓿品种头茬草产量及经济价值的综合评判[J].西北农业学报,2003,12(4):75-81.
[19] 徐丽君,王波,孙启忠.科尔沁沙地紫花苜蓿的光合日动态[J].应用生态学报,2008(1):2189-2193.
[20] 时晓霞.不同秋眠级紫花苜蓿品种在北方农牧交错区生产性能的比较研究[D].呼和浩特:内蒙古农业大学,2007:5.
[21] 董志新,韩清芳,贾志宽,等.不同苜蓿(MedicagosativaL.)品种光合速率对光和CO2浓度的响应特征[J].生态学报,2007(6):2272-2278.
[22] 杨青川,郭文山,康俊梅.17个紫花苜蓿品种产量比较试验[J].中国畜牧兽医,2004,31(12):15-16.
[23] 韩路,贾志宽,韩清芳,等.苜蓿种质资源特性的灰色关联度分析与评价[J].西北农林科技大学学报,2003,31(3):59-64.
猜你喜欢
农业工程学报(2022年3期)2022-04-16
现代畜牧科技(2021年9期)2021-10-13
西南农业学报(2021年7期)2021-08-23
现代畜牧科技(2021年4期)2021-07-21
节水灌溉(2021年5期)2021-05-26
生态学报(2021年6期)2021-04-25
农业工程学报(2021年3期)2021-04-15
节水灌溉(2020年9期)2020-09-16
中国农业科技导报(2020年12期)2020-03-15
中国三峡(2017年4期)2017-06-06
