
滇产纳罗克非洲狗尾草种子种壳透性及种胚休眠性研究
2014-12-21 03:25罗富成段新慧许文花张亚当范素汝郭凤根
草原与草坪 2014年4期
罗富成,彭 健,段新慧,许文花,何 超,张亚当,范素汝,郭凤根
(云南农业大学 动物科技学院草业科学系,云南 昆明 650201)
纳罗克非洲狗尾草(Setariasphacelatacv.Narok)于1974年从澳大利亚引入中国广西、广东、江西等地试种,生长良好;1983年,中澳合作开展云南牲畜与草地改良项目,再次从澳大利亚引入[1]。推广应用表明,纳罗克非洲狗尾草是南方热带、亚热带和暖温带地区草山改良、人工草地建设和高速公路护坡、水库护堤的骨干草种[2-7]。目前种子已国产化,但在大面积生产条件下,其种子产量只有45.0~60.0kg/hm2,发芽率在10%[8],休眠程度深[9],严重制约了优良牧草的推广利用。尽管国内曾围绕纳罗克非洲狗尾草“种子生产关键”技术问题,开展了一系列研究[10,11],使其种性在一定程度上得到了复壮[11],种子产量在小区试验条件下达到了326.4kg/hm2[10],并提出了多项行之有效的栽培管理措施和收种技术,但种子休眠的原因尚不清楚。以云南自产纳罗克非洲狗尾草种子为试验材料,通过机械处理去除稃片、擦破种皮、去除种壳,观测处理前后种子吸水率、呼吸强度的变化及种胚(含胚乳)的萌发特性,检验种壳对种子萌发的影响,判断种胚的休眠性,为纳罗克非洲狗尾草种子休眠机理提供理论依据。
1 材料和方法
1.1 供试材料
试验材料为云南省自产纳罗克非洲狗尾草种子,2013年秋在成熟期收获于昆明市黑龙潭云南农业大学草学基地。种子净度为36.8%,发芽率为26%。收获后,立即将杂质,包括空瘪种子完全清除,用1.5%硫酸铜溶液浸泡消毒10min,晾干,于室内自然贮藏,备用。
1.2 试验方法
1.2.1 种子处理 从消毒后的种子中随机抽取2份材料,一份用于种壳透性测定,处理措施包括用大头针擦破种皮,徒手搓去稃片,搓去稃片后再擦破种皮,获得带稃片擦破种皮种子、去除稃片种子(即裸种子)、擦破种皮的裸种子;另一份经徒手搓去稃片后,再用磨砂纸完全磨去种皮,然后挑选种胚(带胚乳)完好的材料用于检测种胚的萌发性。
1.2.2 种子透水性测定 取带稃片的完整种子为对照(CK)、带稃片擦破种皮种子、裸种子、擦破种皮的裸种子各0.1g(精确到万分之一),分别置于不同的烧杯中,加入蒸馏水,在室温下浸泡0、4、8、12、24、28、32、36h,取出后用滤纸吸干表面水分,然后,迅速用分析天平称重。毎处理重复4次。根据种子吸水前后的重量变化计算种子的即时吸水率,种子开始萌动时的吸水率视为该种子的标准吸水率[12]。计算公式[13]:
种子吸水率(%)=(W2-W1)/W1×100%式中:W2为种子吸水后重量;W1为种子吸水前重量。1.2.3 种子透气性测定 取带稃片的完整种子为对照(CK)、带稃片擦破种皮种子各0.2g(精确到万分之一),在室温下用蒸馏水浸泡0、6、12、24、36、48、60h后,用美国LICOR公司生产的LI-6400光合作用仪测定呼吸强度,待其数据稳定后记录,每个处理4次重复,每个重复记录3次数据。比较擦破种皮前后种子呼吸强度的变化,确定种皮透气性的高低。按仪器说明书,呼吸强度的计算公式:

式中:RI为种子的呼吸强度,μmol/(g·min);R(CO2)为 参 比 室 二 氧 化 碳 浓 度,μmol CO2/mol;S(CO2)为样品室二氧化碳浓度 μmol CO2/mol;R(H2O)为参比室水浓度,mmol H2O/mol;S(H2O)为样品室水浓度,mmol H2O/mol;flow为气体流进样室的流速,μmol/s;W为样品重量g。
1.2.4 种胚萌发性测定 以离体种胚(含胚乳)为研究对象,以完整种子为对照(CK),于人工气候箱内在MS培养基上进行培养,4次重复,每重复100粒。箱内相对湿度为70%,温度为(30±1)℃,24h光照。第7d和14d分别统计种子发芽势和发芽率。发芽结束后,将幼苗置于105℃的烘箱中杀青15min,然后在80℃的烘箱中干燥24h[8],称其干重,并计算种子发芽指数和活力指数[14]。
1.3 数据处理
所有数据整理和作图用Excel完成,统计分析用Spss 17.0软件进行。
2 结果与分析
2.1 纳罗克非洲狗尾草种子的透水性
随浸种时间的延长,带稃片的完整种子(CK)、带稃片擦破种皮种子、裸种子和擦破种皮的裸种子,吸水速率的变化趋势相同(图1)。吸水初期,各处理种子的即时吸水率均快速上升,浸种32h后,吸水趋于稳定,接近饱和状态,此时种子逐渐开始萌动,吸水率不再增加,但均显著(P<0.01)高于浸种初期(0~12h)。无论是否擦破种皮,只要带稃片,种子吸水率均保持在55.0%,去稃片后种子吸水率则均停留在30.5%,前者较后者高24.5%;不仅如此,在种子整个吸水过程中,带稃片种子的即时吸水率均远大于去稃片种子。究其原因,是由于纳罗克非洲狗尾草种子具有能充分吸水、半张开、比重较大的附属物——稃片所致(图2);此外,无论是否去除稃片,只要擦破种皮,种子的吸水速度明显加快,说明擦破种皮可改善种子的透水性。这一点可由其致密而坚硬的种壳结构加以佐证(图2、3)。

图1 纳罗克非洲狗尾草种子吸水率Fig.1 The change of water absorption on'Narok'seeds

图2 纳罗克非洲狗尾草种子电镜扫描图Fig.2 ‘Narok’seed SEM image

图3 纳罗克非洲狗尾草种子体视显微图Fig.3 ‘Narok’seed stereo microscope image
2.2 纳罗克非洲狗尾草种子的透气性
浸种时间为0h(即干种子)时,CK与擦破种皮种子的呼吸强度分别为 0.044 3μmol/(g·min)和0.227 4μmol/(g·min),后者是前者的5.1倍,擦破种皮可显著提高种子透气性(P<0.01)。从种子浸泡、吸水膨胀过程来看,浸泡初期(0~6h),擦破种皮种子的呼吸强度显著高于完整种子(P<0.01),之后,二者的呼吸强度具有一致的变化趋势,即随着浸种时间的延长,呼吸强度逐步提高,至36h后逐渐开始下降,接近60h达到同一呼吸水平(图4)。呼吸强度的下降与此时体内积累的CO2浓度有关。整个吸胀过程,擦破种皮种子的呼吸强度均高于完整种子,说明擦破种皮可增强种子透性,从而促进种子呼吸。浸种36h,完整种子和擦破种皮种子的呼吸强度达到最高值,分别为0.416 7μmol/(g·min)和0.402 1μmol/(g·min),是浸种前的1.83倍和9.07倍。浸种,尤其是擦破种皮后再浸种,对促进种子呼吸的效果十分显著(P<0.01)。
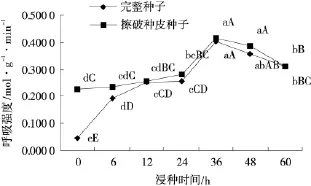
图4 不同浸种时间纳罗克非洲狗尾草种子的呼吸强度Fig.4 The Change of respiratory on Setaria sphacelata cv.‘Narok’seeds
2.3 纳罗克非洲狗尾草离体种胚的萌发性
2.3.1 离体种胚发芽势和发芽率 纳罗克非洲狗尾草离体种胚与完整种子(CK)的发芽势和发芽率均随着贮藏时间的延长而逐步递增,最后稳定在一定水平上(图5,6)。离体种胚和完整种子均存在休眠现象。
种子收获时(贮藏0d),离体种胚的发芽势与完整种子无差异(P>0.05),发芽率已有差异(P<0.05);贮藏30~150d,其发芽势较完整种子有显著差异;贮藏0~180d,其发芽率均高于完整种子。种壳限制了种子的萌发。
从贮藏时间对材料本身的影响来看,贮藏30d后,离体种胚的发芽势为40%,发芽率为49%,与种子收获时差异极显著(P<0.01),休眠的种胚已开始萌动,至120d前后发芽势、发芽率分别稳定在60%和70%,分别是种子收获时的2.8倍和1.7倍,种胚休眠已基本解除。贮藏60d后,对照——完整种子的发芽势、发芽率分别达到36%和44%,较种子收获时均有显著提高(P<0.01),同时随贮藏时间的延长种壳对种胚的束缚已逐渐削弱,至210d前后已完全失去其负面影响,发芽势、发芽率分别稳定在67%和74%,分别是种子收获时的3.0倍和2.8倍。
2.3.2 离体种胚的发芽指数和活力指数 随贮藏时间的延长,离体种胚及完整种子(CK)的发芽指数和活力指数均呈现逐渐增加的趋势(表1)。离体种胚贮藏至60d,发芽指数、活力指数递增变缓,指标值较前期有显著差异(P<0.05),较后期差异不大(P>0.05),此时种胚休眠多已释放;贮藏至150d活力指数、贮藏至180d发芽指数分别达到最大值(P<0.05),说明种胚休眠已完全解除。完整种子贮藏180d,活力指数递增变缓,贮藏至210d,发芽指数递增减缓,两指标较前期均有显著差异(P<0.05),较后期差异不大(P>0.05),此时种子的综合休眠已基本解除。
种子收获时(贮藏0d),离体种胚的发芽指数、活力指数较完整种子(CK)均无差异(P>0.05)。此时,影响种子萌发的主要原因并非种壳的束缚,而是种胚的不成熟。贮藏30~180d,离体种胚的发芽指数、贮藏30~150d离体种胚的活力指数均显著大于完整种子(P<0.05),之后无差异(P>0.05)。说明此段时间,种壳对种子萌发也同样具有显著的抑制作用。之后的各个贮藏时期,离体种胚的发芽指数和活力指数较完整种子均无显著差异(P>0.05),并稳定在一定水平上(表1)。

图5 纳罗克非洲狗尾草离体种胚的发芽势Fig.5 The germination potential of embryo in vitro of‘Narok’seeds

图6 纳罗克非洲狗尾草离体胚的发芽率Fig.6 The germination rat of embryo in vitro of‘Narok’seeds

表1 不同贮藏时间纳罗克非洲狗尾草离体种胚的发芽指数与活力指数Table1 The change of different storage time for embryo of‘Narok’seeds on germination index and vigor index
综上所述,纳罗克非洲狗尾草种子的休眠属于综合休眠,除种壳阻碍种子萌发外,种胚(含胚乳)也会引起种子休眠。在种子贮藏60d后,休眠的种胚多数已开始自然释放休眠,至180d,种胚休眠已完全解除;在种子贮藏210d后,种壳基本失去对种子萌发的阻碍作用。因此,纳罗克非洲狗尾草种子休眠期210d。
3 讨论
(1)由于纳罗克非洲狗尾草种子具有能充分吸水、半张开、比重较大的附属物稃片,无论是否擦破种皮,带稃片种子的吸水率较去稃片种子高24.5%;无论是否去除稃片,只要擦破种皮,均可改善种子透水性,种子吸水速度明显加快,这些特性都与热带牧草木质化程度高有关,这与温带牧草[15]稍有不同。擦破种皮后,纳罗克非洲狗尾草干种子的呼吸强度较擦破种皮前提高4.1倍;无论是否擦破种皮,浸种时间(36h)均能显著提高种子的呼吸强度(P<0.01)。说明种子稃片及致密的种皮阻碍了种子内外气体交换,而吸水膨胀能够改善种子内外气体交换状况。说明纳罗克非洲狗尾草的种壳(含种皮)是引起种子休眠的一个重要原因。
(2)在种子收获后不同的贮藏阶段(0~240d),离体种胚和完整种子的发芽势、发芽率、发芽指数和活力指数均随贮藏时间的延长而逐步递增,最后稳定在一定水平上。说明离体种胚和完整种子均存在休眠现象。种子发芽指数是对发芽率的细化和深化,它放大了种子活力特征,使优劣种子的差异加大[16],评判结果更可靠。种子收获时(贮藏0d),离体种胚的发芽指数、活力指数较完整种子(CK)均无差异(P>0.05),说明此时影响种子萌发的主要原因除种壳障碍外,尚存在着种胚休眠。离体种胚贮藏至60d,发芽指数、活力指数递增同时变缓,指标值较前期有显著差异(P<0.05),较后期差异不大(P>0.05),说明此时种胚休眠多已开始释放,这与曹帮华等[17]对银杏种子、张恩和等[18]对野生羌活种子的研究结果一致;贮藏30~180d,离体种胚的发芽指数、贮藏30~150d离体种胚的活力指数均显著大于完整种子(P<0.05),之后无差异(P>0.05)。说明此时种壳对种子萌发仍然存在显著的抑制作用,这与冯毓琴等[19]对苜蓿、王永春等[20]对肥皂草的研究结果一致。之后的各个贮藏时期,离体种胚的发芽指数和活力指数较完整种子均无显著差异(P>0.05),并稳定在一定水平上,说明此时种子综合休眠已基本解除。但种胚(带胚乳)引起种子休眠的具体原因有待于进一步研究。
4 结论
(1)纳罗克非洲狗尾草的种壳(含种皮)有阻碍种子吸收水分、进行气体交换的负面作用,是引起种子休眠的重要原因之一,种胚(含胚乳)也具有休眠特性,其种子休眠属于综合休眠。
(2)在纳罗克非洲狗尾草种子收获后不同的贮藏阶段(0~240d),离体种胚和完整种子的活力指标均随贮藏时间的延长而逐步递增,最后稳定在一定水平上。在种子贮藏后60d,种胚休眠多数已开始释放;贮藏至180d,已释放完毕。种壳在种子贮藏210d已基本失去对种子萌发的束缚作用,纳罗克非洲狗尾草种子的休眠期至少为210d。
[1] 邓菊芬,马兴跃,尹俊,等.混合选择对纳罗克非洲狗尾草草种性复壮的影响[J].草业科学,2008,25(1):43-46.
[2] 陈宝书.牧草饲料作物栽培学[M].北京:中国农业出版社,2001:386-387.
[3] 崔阁英,邓菊芬,尹俊.纳罗克非洲狗尾草及种收获时间的研究[J].草业与畜牧,2009(1):11-14.
[4] 杨卫良,张贤,张红映,等.非洲狗尾草遗传分化调查[J].草业与畜牧,2006(9):20-22.
[5] 袁福锦,奎嘉祥,谢有标,等.云南南亚热带多雨区引进禾本科牧草的适应性及评价[J].草原与草坪,2011,31(5):60-64.
[6] 袁福锦,徐驰,杨雪梅,等.云南暖温带不同季节饲草混播组合筛选及养分测定[J].草原与草坪,2005(6):23-26.
[7] 罗富成,蒋德芸,唐忠华.提高纳罗克非洲狗尾草种子发芽率的研究[J].草业科学,2001,18(1):25-27.
[8] 罗富成,段新慧,彭健,等.纳罗克非洲狗尾草种子休眠人工破除方法[J].草业科学,2014(2):256-263.
[9] 邓菊芬,张美艳.应用灰色系统理论评价纳罗克非洲狗尾草种子生产的田间栽培技术[J].草原与草坪,2009(1):27-32.
[10] 邓菊芬,尹俊,张美艳,等.纳罗克非洲狗尾草种子生产关键技术研究[J].草业与畜牧,2010(5):1-6.
[11] 邓菊芬,马兴跃,尹俊,等.混合选择对纳罗克非洲狗尾草草种性复壮的影响[J].草业科学,2008,25(1):43-46.
[12] 罗富成,毕玉芬,黄必志.草业科学实践教学指导书[M].昆明:云南科技出版社,2008:19.
[13] 张永清,顾振新,张颖,等.豆芽生产中大豆浸泡条件与吸水率和发芽率的关系研究[J].食品研究与开发,2007,28(11):26-29.
[14] 邢妍妍.玉米种子活力形成及休眠机理研究[D].泰安:山东农业大学,2006.
[15] 何学青,胡小文,王彦荣.羊草种子休眠机制及破除方法研究[J].西北植物学报,2010(1):120-125.
[16] 张迄博,杨璇,谭茜.中性笔芯残留物对水稻种子萌发的影响[J].发明与创新,2013(7):23-25.
[17] 曹帮华,蔡春菊.银杏种子后熟生理与内源激素变化的研究[J].林业科学,2006,42(2):32-37.
[18] 张恩和,陈小莉,方子森,等.野生羌活种子休眠机理及破除休眠技术研究[J].草地学报,2007,15(1):509-514.
[19] 冯毓琴,曹致中.天蓝苜蓿种子休眠特性的研究[J].草业科学,2003,20(1):20-23.
[20] 王永春,罗铮,曲超,等.肥皂草种子的休眠和萌发特性初探[J].植物生理学通讯,2007,43(3):491-493.
猜你喜欢
粮油与饲料科技(2022年2期)2022-11-24
中国油料作物学报(2022年2期)2022-05-13
黑龙江大学工程学报(2021年2期)2021-06-22
作文周刊·小学一年级版(2020年40期)2020-10-19
山东林业科技(2019年2期)2019-06-03
现代园艺(2017年11期)2017-06-28
奥秘(2016年10期)2016-12-17
小学生作文选刊·低年级版(2016年6期)2016-06-24
中国卫生标准管理(2015年6期)2016-01-14
中国粮油学报(2014年8期)2014-02-06
