
菜豆种皮开裂过程中木质素含量及相关酶活变化
2021-06-22 09:05:56安普南杨晓旭冯国军闫志山刘大军
黑龙江大学工程学报 2021年2期
安普南,杨晓旭,刘 畅,冯国军,闫志山,刘大军
(黑龙江大学 现代农业与生态环境学院,哈尔滨 150080)
0 引 言
菜豆(PhaseolusvulgarisL.)又名四季豆、棒豆及芸豆等,短日型植物,在豆科植物种植面积中仅次于大豆[1-2]。菜豆的种皮在菜豆籽粒的形成和生长发育过程中起着重要的保护作用,种皮开裂会导致子叶暴露在外,受到外界生物因素和非生物因素的破坏,导致种子结构的破损,严重影响到种子的活力和商品性。探究种皮开裂的原因,对提高菜豆种子质量有着重要的意义。
影响种皮开裂的因素有很多,籽粒性状的表型由基因型、生态环境以及基因型与生态环境互作多方面共同决定[3],其中低温[4]和酚类物质积累[5-6]被认为是导致大豆种皮开裂的主要原因。许多酚类物质是木质素合成的前体物质,参与调节植物体内木质素的合成[7]。有研究表明,相比于正常的种子,开裂的种皮中有更高含量的原花青素和木质素[4,8]。木质素是植物细胞中苯丙氨酸代谢途径产生的最重要的次生代谢产物之一[9];其中苯丙氨酸解氨酶(PAL)是连接初级代谢和苯丙烷类代谢、催化苯丙烷类代谢第一步反应的酶,是植物体木质素生物合成的的关键酶和限速酶[10]。菜豆PAL2基因被证明与木质素前体合成有关,木质素的合成来自苯丙烷类代谢途径中的苯丙氨酸[11]。PAL可被许多外界因素诱导,低温、机械损伤、光、昆虫取食等都可以诱导PAL基因的表达[12]。过氧化物酶(POD)[13]是该途径木质素合成的最后一步酶,木质素通常通过过氧化物酶(POD)和漆酶(LAC)与3种主要类型的单木酚醇聚合。种皮开裂的机理研究在花生[14]、鹰嘴豆[15]、大豆[3-4,8,16]中均有研究,但在菜豆中研究较少,本研究旨在探究菜豆种皮开裂的潜在原因,为改良菜豆种子质量奠定理论基础。
1 材料方法
1.1 实验材料
通过筛选收获的菜豆种子,选取了黑大盛冠、黑大黛冠和1871-11 3个具有种皮开裂缺陷的品种为实验材料。其中黑大黛冠和黑大盛冠拥有相似的荚型,均为绿色宽荚;1871为黄色窄荚。黑大黛冠和1871种皮多为白色,1871的种脐部位有少许黑色;黑大盛冠种皮表面有大量花纹浮现。黑大黛冠和1871种皮的自然开裂率相近,黑大盛冠种皮的自然开裂率相对较低。
1.2 低温处理
在采收的相同品种的菜豆种子中,发现生殖生长时期平均气温低的种子开裂率较高,结合这一现象,低温促进植物木质素生物合成的研究也均有报道。因此推测低温是导致菜豆种皮木质素含量升高的环境因素之一,并进行了实验验证。种皮的颜色与花青素的含量相关,花青素的合成也会受到许多环境因素的诱导,为了减少其对实验的干扰,选取种皮自然开裂率相近、种皮颜色相近的黑大黛冠和1871-11的鼓粒期(R6)豆荚进行低温处理,同一品种常温处理作对照放置于人工气候箱内。
常温条件:25 ℃白天(08:00—18:00);20 ℃黑夜(18:00—08:00)。低温条件:15 ℃白天(08:00—18:00);10 ℃黑夜(18:00—08:00)。分别取低温处理后0 d、3 d、6 d、9 d、12 d的种皮测定其木质素含量、PAL酶活性,比较分析低温与常温状态对菜豆种皮木质素以及PAL酶活性的差异性。
1.3 木质素含量测定
选取自然开裂率差距较大的黑大黛冠和黑大盛冠始粒期(R5)、鼓粒期(R6)、初熟期(R7)、完熟期(R8)4个时期的菜豆种皮,将完整的菜豆种皮剥出,加入液氮研磨成粉末,并参考张水明的方法进行木质素含量测定。将木质素标准品配制成0.125、0.062 5、0.031 25、0.015 625、0.007 812 5、0.003 906 25 mg·mL-1共6个浓度梯度,测定其OD值,绘制标准曲线,得线性回归方程为y=0.059 1x-0.000 5,R2=0.999 5,线性关系良好。
1.4 POD活性测定
1.4.1 酶液的制备
将初熟期(R7)黑大黛冠和黑大盛冠的种皮(开裂种皮和正常种皮互为对照)完整剥出,加入液氮研磨成粉末,称取0.2 g的种皮粉末,加入含有1% PVP的pH=7.0的磷酸缓冲液4 mL,研磨成匀浆,4 000 rmp离心10 min,上清液即为所需酶液。
1.4.2 反应液的制备
取pH=7.0的磷酸缓冲液50 mL,加入28 μL的愈创木酚,19 μL的30%的过氧化氢。取3 mL反应液,0.1 mL酶液,在吸光值为470 nm处测定吸光值,每0.5 min测定1次。
1.5 PAL活性测定
在开裂种皮与正常种皮酶活性的对比中,将初熟期(R7)黑大黛冠和黑大盛冠的种皮(开裂种皮和正常种皮互为对照)完整剥出,用索莱宝PAL活性检测试剂盒测定PAL酶活性。
在低温对种皮PAL酶活性的影响实验中,将低温处理时长为6 d、9 d、12 d黑大黛冠和黑大盛冠种皮(相同时长的低温处理与常温处理互为对照)完整剥出,用索莱宝PAL活性检测试剂盒测定PAL酶活性。1 g组织在1 mL反应体系中1 min使 290 nm 下吸光值变化 0.1 定义为一个酶活性单位。
1.6 组织学观察
取黑大黛冠初熟期(R7)的正常和开裂的种皮,进行石蜡切片。传统石蜡切片在包埋前进行的脱色、透明等处理对种皮内物质有影响,结合传统石蜡切片与Senda等人的方法进行修改。
1.6.1 脱色、包埋与切片
取黛冠新鲜种皮,修剪成长约1 cm的细条状,立即浸泡在乙醇冰醋酸溶液(3∶1,v/v)中进行脱色。清水冲洗2次进行石蜡包埋,冷却后用石蜡切片机制备10 μm切片;在载玻片上涂上蛋白胶,把蜡带转移到载玻片上,右端贴标签,在42 ℃烫板上展片。
1.6.2 脱蜡、染色
将2~5张待染的片子放入二甲苯中脱蜡。停留3~10 min;之后依次放入等量二甲苯和无水乙醇;无水乙醇、乙醇(95%、83%、70%、50%、35%)、蒸馏水中各3~5 min。载玻片从水中取出后晾干,放置一滴DMACA染液10 min用于检测PA,放置盖玻片,用光学显微镜观察到PA被染成蓝色;放置一滴间苯三酚-盐酸染液,浸泡10 min,然后在光学显微镜下观察木质素被染成粉红或紫红色。
1.6.3 染色剂配制
间苯三酚—盐酸染液:在CH3OH和10 M HCl(3∶1,v / v)的混合液中加入1%(w / v)间苯三酚。
2 结果分析
2.1 菜豆种皮开裂发生的时期
菜豆生殖生长可划分为初花期(R1)、盛花期(R2)、初荚期(R3)、盛荚期(R4)、始粒期(R5)、鼓粒期(R6)、初熟期(R7)、完熟期(R8)(图1(a))。通过对不同时期的菜豆农艺性状的观察,在R7时期发现了种皮开裂的菜豆种子(图1(b))。在黛冠开裂的种皮上往往有褐变出现(图1(b),(c)),用间苯三酚—盐酸染液对黛冠正常种子和开裂种子整染后,发现开裂种皮在裂口边缘部位变成红色,在没有发生开裂但出现褐变的种皮上也出现了红色,正常种子却没有发生任何变化(图1(d)),这说明木质素在种皮褐变和开裂的部位沉积。
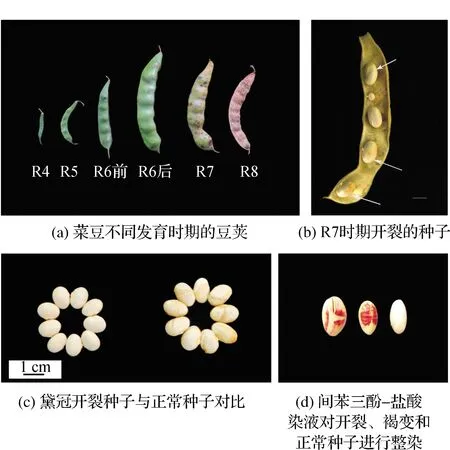
图1 农艺性状观察Fig.1 Agronomic traits observation
2.2 低温处理种皮木质素及酶活测定
对黑大黛冠,1871-11的R6时期豆荚进行低温处理,常温处理作对照,测定木质素含量、PAL酶活性分析比较差异。结合图1发现种皮开裂最先在R7时期出现,选择R6时期的豆荚作为低温处理材料。
在常温处理组中,随着时间的增长种皮中木质素的含量也逐渐升高,结合图4种子发育过程中木质素的含量的变化,可以推测是储存过程中种子的成熟老化导致的。相对于常温处理组,低温处理的种皮木质素含量显著升高。在低温处理的第6天发现黛冠品种的种皮的木质素含量显著上升,为对照组的1.85倍;在第9天与常温处理组相比1871冷处理种皮的木质素含量也达到了显著差异水平,是常温处理组含量的2.13倍。直到第12天,木质素含量持续升高,并且同一品种间低温胁迫的种皮木质素含量同样显著高于常温处理组(图2)。

图2 不同时间的低温处理菜豆种皮木质素含量变化 Fig.2 Changes of Lignin content in Phaseolus vulgaris seed coats treated with low temperature for different time 注:不同小写字母表黛冠品种数据差异显著 (P<0.05) , 不同大写字母表示1871品种数据差异显著 (P<0.05)。下同
在第6天分别测定了2个品种种皮PAL活性,发现低温处理的种皮PAL活性明显增强。随着低温处理时间的增长,PAL活性的差异逐渐加大。在第12天,与常温处理组相比,同一品种内冷处理的PAL酶活性差异达到最大值,其中1871低温处理酶活性为27.15 U·g-1,是常温组的2.65倍,黛冠冷温处理组酶活为6.08 U·g-1,达到常温组的6.06倍,这与图2所测定的木质素含量变化呈相同趋势(图3)。
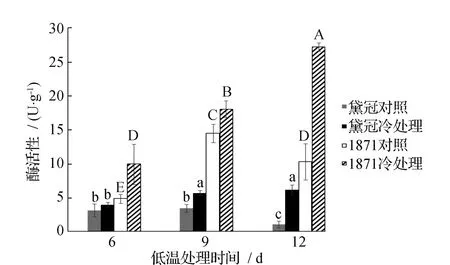
图3 不同时间的低温处理菜豆种皮PAL活性的变化 Fig.3 Changes of PAL activity in bean seed coats treated with low temperature for different time
2.3 发育时期种皮木质素含量变化
分别取黛冠和盛冠R5~R8 4个时期的种皮,利用巯基乙酸法测定菜豆种皮中总木质素的含量。随着种子的成熟,种皮中木质素的含量逐渐升高,在R7阶段出现了开裂的种子(图1b)。相对于正常种皮,开裂种皮的木质素含量明显上升。R7时期盛冠开裂种皮木质素含量为对照组的2.69倍,黛冠开裂种皮木质素含量为对照组的2.51倍;R8时期种皮木质素含量再次升高,盛冠开裂种皮木质素含量达到3.35 mg·g-1,为对照组的1.15倍;黛冠开裂木质素含量升至4.05 mg·g-1,为对照组的4.41倍(图4)。在R7和R8两个时期中,开裂种皮中木质素的含量相比正常种皮显著升高,定量分析结果与上述切片观察结果一致。可推测木质素的含量对种皮开裂的发生有影响。

图4 不同生殖生长时期种皮木质素含量Fig.4 Lignin content of seed coat at different reproductive growth stages注:不同小写字母表盛冠品种数据差异显著 (P<0.05) , 不同大写字母表示黛冠品种数据差异显著 (P<0.05)
PAL是这条代谢途径的第一个也是最重要的酶;POD催化细胞壁木质素生物合成的最后一步。分别测定R8时期的黛冠对照、黛冠开裂、盛冠对照和盛冠开裂种皮的上述酶活性,分析比较其差异性。结果表明在R8时期,黛冠开裂种皮的PAL、POD活性分别为对照组的1.18倍和20.84倍,差异达到显著水平,其中POD活性差异达到极显著水平;盛冠开裂种皮的PAL、POD活性分别为对照组的1.13倍和2.80倍,差异性达到显著水平,其中POD活性差异也达到了极显著水平(图5)。作为木质素合成途径的关键酶,PAL、POD活性的增强促进苯丙氨酸次生代谢途径进行,导致木质素在种皮中积累。
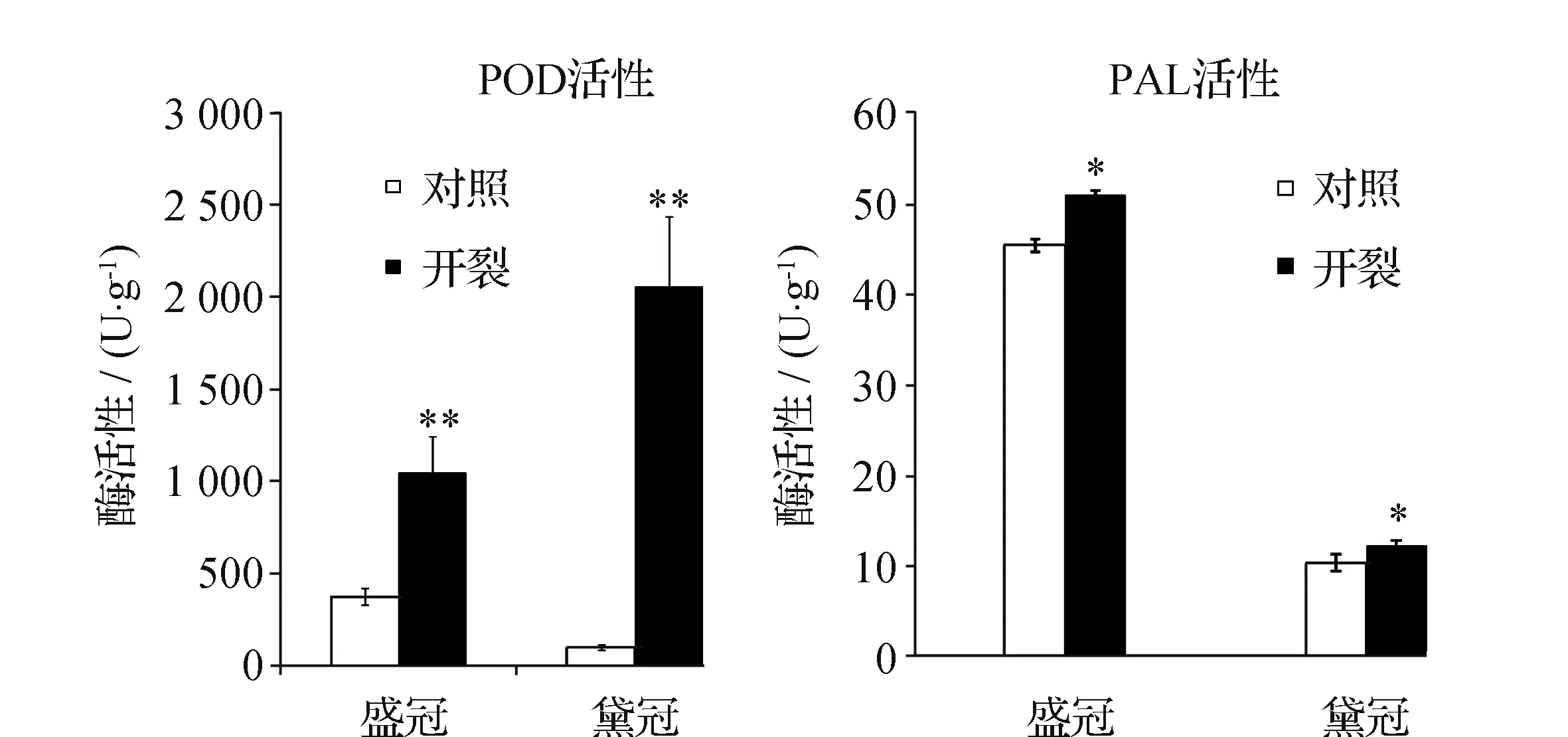
图5 R8时期开裂种子与正常种子种皮PAL和POD活性 Fig.5 PAL and POD activities of seed coats of cracked seeds and normal seeds in R8 注:*表示差异显著 (P<0.05) , **表示差异极显著 (P<0.01)。
2.4 切片观察
用间苯三酚-盐酸染色观察种皮中木质素的积累情况(图6),在种皮的背部切片中,正常种皮切片没有颜色变化(图6(c));开裂种皮的角质层、栅栏层和垫状物都被染成不同程度的红色(图6(a)、6(b)),说明木质素在开裂种皮的背部累积。种脐切片中可发现种脐部位被间苯三酚盐酸染液染成分红或紫红色(图6(d)、6(e)),这与报道的木质素会在种脐部位沉积一致;种脐部位的种皮有两层栅栏层,开裂种皮的上栅栏层和下栅栏层的连接部位染成红色(图6(d)),说明木质素在上下栅栏层的连接部位沉积。并且,在开裂种皮切片的过程中往往会出现栅栏细胞与垫状物细胞分离、错位,垫状物细胞基部与其下部的薄壁细胞分离的现象(图6(f)),这也说明木质素的沉积会降低种皮中各个细胞层连接紧密程度,在外力的作用下更容易发生开裂。
3 讨 论
菜豆种皮开裂严重影响菜豆品种种子质量,结合前人在大豆等豆科植物中的研究,对菜豆种皮开裂发生的时期和主要原因进行讨论。
3.1 菜豆种皮开裂的时期
种皮的分化是在双受精之后开始的,受精完成后中央细胞会受到一个受精信号,该信号使得珠被细胞的细胞层开始分化形成种皮。在豆类植株生殖生长时期(R1~R8)都有可能发生种皮开裂的现象[17],在大豆种皮开裂的研究中最先在始粒期(R5)发现了种皮开裂的种子[18];本研究在R7时期发现了裂种皮的籽粒,种皮开裂的同时往往伴随着种皮褐变的发生,在同一个豆荚中的种子没有发生开裂的种皮背部也会有褐变现象出现(图1(b))
3.2 低温促进种皮木质化
木质化现象是果实在受到低温冷害等胁迫时的一种症状。通过对R6时期的豆荚低温处理发现,随着低温时间的延长,种皮中木质素的含量升高。发现低温会加快菜豆的苯丙氨酸代谢途径,增强该途径中的酶活性,导致木质素在组织中累积。在拟南芥的叶片中发现低温会影响AtPAL1和AtPAL2的表达[19];并且低温胁迫也会导致烟草中PAL等苯丙烷途径表达基因的上调,从而导致烟草木质素合成增加[20]。实验结果表明低温增加苯丙烷途径中PAL酶活性,加快木质素的合成,导致木质素在种皮中沉积。
3.3 种皮内PAL和POD酶活性增强促进了木质素合成
植物体内木质素合成的最主要的途径是苯丙烷途径,种皮内木质素的累计与苯丙烷途径的促进有很大的关系。对R8时期开裂种皮和正常种皮中PAL和POD两种酶活性以及木质素含量进行比较,发现无论是黛冠还是盛冠,裂种皮的两种酶活性都要显著高于正常种皮,其中POD活性的差别最为明显,最高可达到正常种皮的20.80倍。在无壳美洲南瓜的研究中,薛应钰等[21]发现无壳美洲南瓜种皮中缺乏木质素的合成,并且PAL和POD酶活性的变化与木质素合成呈正相关性。PAL和POD作为苯丙烷途径合成木质素的关键酶,在许多植物的研究中也证实了木质化与 PAL活性呈正相关[17]。研究结果也证实了PAL和POD的活性与木质素含量呈正相关这一观点。
3.4 种皮木质素沉积导致种皮开裂
开裂的种皮往往伴随着木质素含量的升高。通过石蜡切片染色观察,发现在黛冠开裂种皮的背部都有木质素的沉积,并且在木质素沉积的种皮中,出现了栅栏细胞、垫状物细胞和薄壁细胞之间错位分离的现象。说明木质素的沉积会降低种皮中各个细胞层连接紧密性,在外力的作用下更容易发生开裂。种皮木质素定量分析与切片观察的结果一致,随着植株生殖生长的进行,种皮中木质素含量逐渐升高,并且开裂种皮中的木质素含量要显著高于正常种皮。也有研究发现木质素含量增加导致种皮的开裂:Mineo S等[8]在大豆的种皮切片中发现,大豆开裂种皮的栅栏层和垫状物细胞层的连接处有木质素的沉积。文献[4]指出原花青素和木质素在种皮内沉积会降低种皮的物理强度,随着子叶的生长发育在种皮腹部形成裂纹。木质素的沉积是种皮开裂的主要原因。
4 结 论
在菜豆经低温处理之后种皮木质素的含量显著上升,但由于菜豆的种皮开裂也受到子叶发育和菜豆生殖生长等方面的影响,在短时间内没有观察到开裂种子的产生。综上所述,得到了两个结论:①低温增强了菜豆种皮内PAL酶活性,导致木质素在菜豆的种皮背部沉积;②开裂的种皮中木质素含量及相关酶活性要显著高于正常种皮,木质素的沉积使得种皮各细胞层之间更容易发生分离从而导致种皮开裂。
猜你喜欢
粮油与饲料科技(2022年2期)2022-11-24 13:12:07
钻井液与完井液(2022年4期)2022-10-26 06:39:38
哈尔滨轴承(2021年1期)2021-07-21 05:43:14
福建中医药(2017年6期)2018-01-09 06:49:54
电镀与环保(2017年3期)2017-06-23 08:24:51
西南农业学报(2016年4期)2016-05-17 05:42:14
金属矿山(2014年7期)2014-03-20 14:19:52
中国粮油学报(2014年8期)2014-02-06 01:34:17
食品科学(2013年13期)2013-03-11 18:24:41
长江蔬菜(2013年23期)2013-03-11 15:09:39
