
黄山市稽灵山次生马尾松种数量动态
2014-10-29 00:40万志兵刘成功祖浩浩
井冈山大学学报(自然科学版) 2014年6期
万志兵,刘成功,祖浩浩
黄山市稽灵山次生马尾松种数量动态
*万志兵,刘成功,祖浩浩
(黄山学院生命与环境科学学院,安徽,黄山 245041)
为掌握稽灵山马尾松次生林种群的数量动态和生存状况,采用“空间取代时间”的分析方法来编制稽灵山马尾松种群静态生命表,并以生存分析为理论基础,对稽灵山马尾松种群动态进行了研究。结果表明:(1)稽灵山马尾松种群结构是纺锤形,幼龄个体和老年个体数量少,中龄个体较多,种群在一定程度上表现为衰退型;(2)种群的死亡率随着龄级的增加而不断增长,死亡率峰值出现在第8龄级;(3)该马尾松种群的存活曲线属于Deevey II型。马尾松是亚热带次生林的过渡树种,其群落的衰退,有利于其他物种的生长,适当辅以人工抚育,对于丰富该地区树种多样性具有重要意义。
静态生命表;年龄级;种群;动态;马尾松
马尾松(Lamb),别名青松、山松,属松科松属常绿乔木,为我国亚热带森林植被演替的先锋树种之一。马尾松种群分布范围极广,北沿秦岭淮河一线,南至海南岛,东到台湾,西达四川,一般在海拔700 m以下生长,并有若干地理变种[1]。马尾松喜微酸性土壤,但其本身对土壤的要求不高,在各种土壤甚至岩石缝中也能生长。马尾松是我国长江流域各省的荒山造林、水土保持、生态建设等方面的重要树种,也是我国南方地区重要的城市绿化和天然景观树种之一[2]。
李根前等[3]研究表明马尾松在山坡中上部及山脊等立地条件较差的生境中能够形成稳定的群落,随着群落的演替,马尾松种群在增长之后趋于衰退。马尾松人工种植时为均匀分布,中龄林阶段变为集群分布,到了成林阶段则成了随机分布[4]。董鸣[5]对缙云山马尾松种群数量动态进行了初步研究,朱守谦等[6]研究了天然更新幼龄种群年龄结构的类型和形成过程。这些研究集中在天然林或人工林的情况,而处于城市边缘的次生林,不断地被人为干扰或破坏导致森林种群各项指数均与天然种群有较大的差异。为此本研究选择黄山市屯溪区稽灵山马尾松种群为研究对象,对该种群的种群动态和年龄结构进行了研究和分析,旨在为该地的森林经营管理提供科学参考依据。
1 材料与方法
1.1 马尾松研究种群概况
稽灵山位于安徽省黄山市屯溪区和休宁县交界处,地处中亚热带北缘,处于天目山、黄山之间的休屯盆地,是安徽、浙江、江西三省的交界处,也是横江、率水、新安江交汇之处。该地四季分明,气候温和,雨量充沛,年平均气温16.3 ℃,全年无霜期237 d,年降水量1670 mm。稽灵山植被除了马尾松外,伴生的树种还有枫香(杉木()、青冈()、小构树()、檵木()等。
1.2 研究方法
1.2.1 调查方法
2013年5月,在稽灵山植被较为完整的地带选择10个样方,每个样方的面积为1 hm2(100 m × 100 m),总面积为10 hm2。并对样方内的所有成活马尾松进行每木调查,记录每一株马尾松的树高和胸径。
1.2.2 龄级结构分析
采用“空间取代时间”的方法,即用立木的胸径结构代替年龄结构的方法对该马尾松种群进行动态分析,按照1960年森林专业调查办法草案要求并结合本研究实际情况,以所统计的马尾松立木的胸径大小为标准将样地内的马尾松划为9个龄级。本研究中调查到的个体胸径最小为4.1 cm,没有小于4 cm的马尾松个体,具体划分情况如下:4 cm ≤ 胸径< 10 cm为第I径级;10 cm ≤ 胸径< 15 cm为第II径级;15 cm ≤ 胸径< 20 cm为第III径级;以此类推至第VII径级;40 ≤ 胸径< 50 cm为第VIII径级;50 cm以上为第IX径级。然后第I径级对应第1龄级,第II径级对应第2龄级,……,如此一一对应,分别对各个径级的马尾松株数进行统计,并编制稽灵山马尾松种群的静态生命表,分析其种群的动态变化。
本研究采用陈晓德[7]通过数学模型推导出的种群年龄结构的动态指数对稽灵山马尾松进行定量描述,其计算方法如下:

式中:Vn为种群从n到n+1级的个体数量变化动态;Vpi为整个种群结构的数量变化动态指数;Sn为第n 个龄级种群个体数;Sn+1为第n+1个龄级种群个体数;
当考虑未来的外部干扰时,

式中:K为种群龄级数量;Vpi与Vn取正、负、零值的意义分别反映种群或相邻龄级个体数量的增长、衰退及稳定的动态关系[8]。
1.2.3 静态生命表的编制

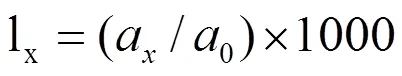

根据上述公示所计算出来的函数值,我们可以分别绘制种群的生存率曲线、累计死亡率曲线、死亡密度曲线和危险率曲线,并以此对稽灵山马尾松次生林种群进行更深层次的探讨和分析。
2 结果与分析
2.1 龄级结构分析
由图1可以看出,稽灵山马尾松次生林种群的龄级结构呈现正态分布。调查样地内马尾松共计4490株。由图1还可以看出,稽灵山马尾松种群中龄林株数较多,尤其是第3龄级,共有1312株,占总调查株数的29.22%。马尾松种群在每一龄级都有分布,第一龄级个体数为96株,占总株数的2.14%,在幼龄阶段,该马尾松种群的个体数目较少;从第3龄级开始,各龄级马尾松的株数随着龄级的增加而大幅减少,第9龄级时马尾松仅余7株,这有可能是由于环境选择、自然和人为干扰以及马尾松自身的生物及生理学特性所决定的,表明该马尾松种群的幼龄阶段发展不稳定,幼龄阶段的马尾松数量太少,所以该马尾松种群在一定程度上有表现为衰退型种群的趋势。
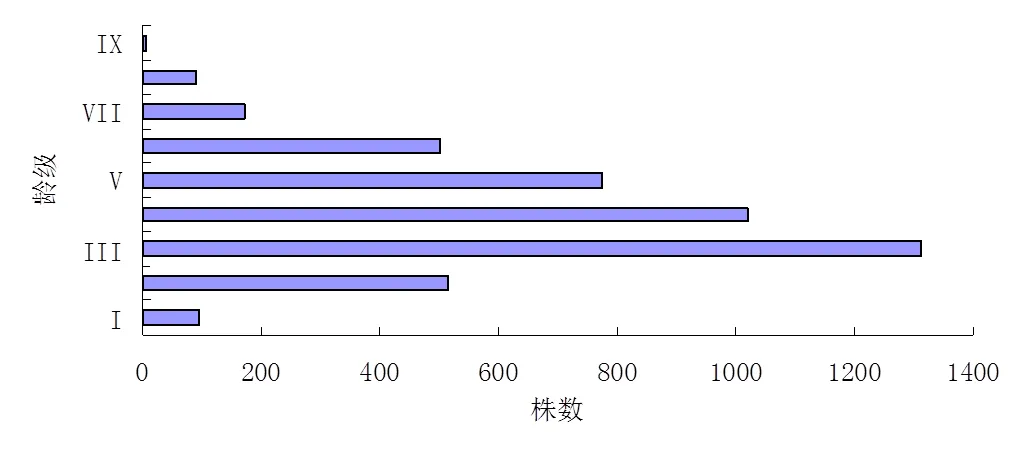
图1 马尾松种群龄级结构
稽灵山马尾松种群的平均胸径为22.53 cm,稽灵山马尾松种群的平均树高为14.673 m,该马尾松种群中个体胸径最大值为65.9 cm,最小值为4.1 cm,树高最大值为24.4 m。
根据数据量化分析的方法,该马尾松种群各相邻龄级间的个体数量的变化动态如下:
2.2 生命表编制及分析
从表1可以看出,该马尾松种群总体生命期望值在第1龄级时为最大值,而生命期望值是衡量单个植株平均生存能力的指标[12]。因此说明,此阶段的马尾松是处在生理活动最旺盛、生存状况最好的阶段。但是从第2到第9龄级这一阶段内,生命期望值都是下降趋势,前一阶段下降可能是由于植株的竞争增强,受到环境筛选和外界干扰的影响;后一阶段可能是由于马尾松种群个体的生理衰退和生存质量的下降所致。稽灵山马尾松种群,其生命期望值都随龄级的增加而逐级减少,可能是由于该马尾松种群处于第1龄级的个体太少,而导致对该种群中第1龄级的生命期望值计算偏高的可能。

表1 马尾松种群静态生命表
2.3 存活曲线和死亡曲线
图2 表明,稽灵山马尾松种群各龄级存活个体数量下降趋势基本持平,各龄级的死亡个体数量在第5龄级前也基本持平,但在第6龄级和第7龄级两个龄级死亡个体数量略有增长,可能是由于马尾松自身的生理机能衰退所导致的结果。Deevey[13]划分了三种类型的存活曲线:(1)凸型,采用指数方程式来表达;(2)对角线型,用直线方程式来表达;(3)凹型,用幂函数方程式来表达,含义是植株的幼体死亡率高。选用以上三种数学模型对马尾松种群的存活曲线类型进行检验,检验结果表明稽灵山马尾松种群存活曲线为Deevey-II型。

图2 马尾松种群的存活曲线(lx)和死亡曲线(dx)
2.4 死亡率和亏损率曲线
根据表1中的数据,可绘制马尾松种群死亡率和亏损率曲线(图3),其中,横坐标为该马尾松种群的各个龄级,纵坐标为计算所得的死亡率和亏损率函数值。图3显示,马尾松种群的死亡率和亏损率曲线在走势上基本一致;伴随着龄级的增加,各龄级死亡率也在增加;但从第1龄级到第5龄级,死亡率增长的涨幅并不大,从第5龄级之后开始,各龄级间的死亡率涨幅变大,死亡率迅速升高。

图3 稽灵山马尾松种群死亡率(qx)和亏损率(Kx)曲线
2.5 种群生存函数分析

表2 稽灵山马尾松种群生存分析函数

图4 马尾松种群生存函数和积累死亡函数曲线

图5 马尾松种群死亡密度函数(fi)和危险率(λ)曲线
依据稽灵山马尾松种群生存分析函数(见表2)可绘制得到4个生存函数曲线,见图4。马尾松种群的生存率表现出一直下降的趋势,累计死亡率呈现单调上升的趋势,两者表现互补形式。生存率和累积死亡率变化幅度始终都比较大,说明该种群数量不稳定。在第8龄级时,马尾松种群的生存率小于5%,累计死亡率大于95%,说明马尾松种群进入生理衰老阶段,其种群数量减少,从而呈现衰退的特征。
由稽灵山马尾松种群死亡密度函数和危险率曲线(图5)可以看出,马尾松种群的死亡密度函数曲线起伏不定。第1龄级时为1.87%,从第2龄级到第5龄级比较稳定,一直处于2.24%~2.26%之间波动;第6龄级出现较高的增长,上升到3.44%;从第7龄级开始以后,该马尾松种群的死亡密度函数又出现迅速下降的趋势,从第7龄级的3.46%降到第8龄级的0.87%。相较于死亡密度函数曲线,该马尾松种群的危险率函数曲线就没有多少起伏,从第1到第7龄级,该种群的危险率持续增长,且从第5龄级开始,到第7龄级结束,该马尾松种群的危险率增长较为迅速,从4.53%迅速增长到19.32%,从第1到第5龄级的增长速度与之相比则较为平缓;从第7龄级往后,该马尾松种群的危险率函数则开始出现下降的趋势。
3 讨论
3.1 马尾松种群动态特征
静态生命表反映多个世代重叠的年龄动态历程中的一个特定时间段内的特点,而不是对这一种群的全部生活史追踪[14]。生命表中径级划分是基于不同年龄径级每年等量增长的假设[15]。生命期望反映植物种群的平均生存能力[16]。本研究中马尾松种群在低龄级时具有较高的生命期望,是因为马尾松个体适应力强。当有林窗出现,光照足够时,马尾松幼株能迅速生长。所以虽然缺乏幼苗,但是一旦幼苗长成幼树,其成活率较高。稽灵山的马尾松种群,其死亡率随着龄级的增加而不断升高,到第6龄级时,稽灵山马尾松种群整体死亡率达到39.3%,第7龄级达到65.1%,而到第8龄级时更是达到93.8%。马尾松林早期是单优势种构成的群落,林分郁闭后,个体间竞争激烈,导致死亡,林下常绿阔叶林树迅速生长。对稽灵山马尾松种群存活曲线的统计和检验,发现其更接近Deevey II型,表明该马尾松种群动态目前正处于一个动态稳定状态。
3.2 影响马尾松种群年龄结构的因素
种群的年龄结构能够在相当大的程度上反映该种群与其所处的环境之间的相互关系和其在该生态群落中的作用和地位[17-18]。本研究结果表明稽灵山马尾松种群径级结构呈现纺锤形,幼龄个体和老年个体数量少,中龄个体较多,表明该马尾松种群幼苗确实很严重,该种群的更新难度很大,中等径级的马尾松对于维持种群结构具有十分重要的意义。由于径级在4cm以下的幼苗幼树缺失,种群的稳定是稳定是暂时的,平衡状态必将会被打破,根本原因与马尾松是先锋树种及其生物学特性有关。马尾松作为先锋树种成了该片次生林的优势树种,当其他一些树种枫香、青冈、小构树等树种进入后,整个群落的郁闭度提高,林内马尾松幼苗无法得到足够的光照,马尾松不耐庇荫,其幼苗无法正常生长[19],而后来进入的植物的幼苗、幼树在群落的生态位中占据了更大的位置,这加剧了马尾松种群径级小的个体比例变小。随着群落的发展和种群自身的变化,大径级马尾松受外界干扰的机会增大,同时由于处于城市的边缘,人为干扰的加剧,导致大量的倒木或折枝的出现。植物种群是一个有限的系统,它是在一定的生境条件下运动,并不断受到环境及人为因素干扰[20]。马尾松作为过渡树种,最终会被其他树种所替代,而且人为干扰活动加速了演替进程。
[1] 郑万钧.中国树木志:第1卷[M].北京:中国林业出版社, 1997:294-297.
[2] 谢春平.南京仙林地区次生林马尾松种群结构分析[J].四川农业大学学报,2012,30(2):156-160.
[3] 李根前,唐德瑞,王迪海,等. 陕南马尾松种群结构与动态的初步研究[J].西北植物学报,1994,14(4):307-313.
[4] 杨心兵,覃逸明,刘胜祥. 武汉市马鞍山森林公园马尾松年龄种群结构与分布格局[J].华中师范大学学报:自然科学版,2001,35(2):209-213.
[5] 董鸣.缙云山马尾松种群数量动态初步研究[J].植物生态学与地植物学学报,1986,10(4):283-293.
[6] 朱守谦,龙翠玲.马尾松幼龄种群年龄结构研究[J].山地农业生物学报,1999,18(5):289-295.
[7] 陈晓德. 植物种群与群落结构动态量化分析方法研究[J].生态学报,1998,18(2):214-217.
[8] 凌云,张光富,王锐.南京老山国家森林公园朴树种群动态[J].生态与农村环境学报,2011,27(2):28-34.
[9] 刘金福,洪伟,吴则焰,等.孑遗植物水松种群生命表和谱分析[J].武汉植物学研究,2008,26(3):259-263.
[10] 洪江.云杉种群动态[M].北京:中国林业出版社,1992:1-7.
[11] 张兴旺,张小平,郭传友,等.安徽琅琊山青檀种群数量动态[J].生态与农村环境学报,2012,28(5):493-499.
[12] 祝宁,植物种群生态学导论[M].哈尔滨:东北林业大学出版社,1982:19-74.
[13] Deevey E S. Life tables for natural populations of animals[J]. The Quarterly Review of Biology, 1947, 22(4): 283-314.
[14] 吴承祯,洪伟,谢金寿,等. 珍稀濒危植物长苞铁杉种群生命表分析[J]. 应用生态学报,2000,11(3):333-336.
[15] 闫桂琴,赵桂仿,胡正海. 秦岭太白红杉种群结构与动态的研究[J]. 应用生态学报,2001,12(6):824-828.
[16] 郭连金,徐卫红. 武夷山米槠种群结构及谱分析[J]. 植物研究,2007,27(3):325-330.
[17] 邓贤兰,曾晓辉,吴新年,等. 井冈山观光木所在群落特征研究[J].井冈山大学学报:自然科学版,2010,31(4): 113-117.
[18] 邓贤兰,赖弥源,叶青富,等. 井冈山交让木种群结构与分布格局研究[J]. 井冈山大学学报:自然科学版,2012,33(6):102-106.
[19] 丁圣彦,宋永昌. 常绿阔叶林演替过程中马尾松消退的原因[J].植物学报,1998,40(8):755-760.
[20] 封磊,洪伟,吴承祯,等.珍稀濒危植物南方铁杉种群动态研究[J].武汉植物学研究,2003,21(5):401-405.
Population structure dynamics ofsecondary forest in Jiling mountain of Huangshan
*WAN Zhi-bing, LIU Cheng-gong, ZU Hao-hao
(College of Life and Environment Sciences, Huangshan University, Huangshan, Anhui 245041, China)
Population structure dynamic and Quantitative statistics ofwas determined in Jiling Mountain. Based on the theory of population life table and survival analysis, the life table ofpopulation was constructed and age classes were replaced with size-classes, thus the population structure dynamic ofwas explored. The results showed that: (1) The population structure ofin Jiling Mountain was fusiform, characterized by the existences of juveniles and older individuals in small numbers, middle age individuals in a larger part, which to a certain degree reflects a recession form of the species; (2) The mortality of such species increased along with the age and has peaked in the eighth age class; (3) The survival curve of thepopulation belonged to the Deevey II type. As a transition population in the subtropic secondary forest, it was a normal phenomenon that the population ofis substituted. It would benefit to the growth of the other species and to enrich species diversity in this region with appropriate forest tending by humans.
static life table;age class;population;dynamics;
Q145/S718.54+2
A
10.3969/j.issn.1674-8085.2014.06.023
1674-8085(2014)06-0107-07
2014-09-01;
2014-10-22
安徽省级优秀青年人才基金项目(2012SQRL185);安徽高校省厅自然科学研究重点项目(KJ2013A241)
*万志兵(1980-),男,安徽无为人,副教授,博士生,主要从事植物栽培和育种研究(E-mail: wanzb626@hsu.edu.cn);
刘成功(1990-),男,安徽合肥人,黄山学院生命与环境科学学院林学专业2011级本科生(E-mail:970974659@qq.com);
祖浩浩(1992-),男,安徽淮北人,黄山学院生命与环境科学学院林学专业2010级本科生(E-mail:867108233@qq.com).
猜你喜欢
湖南生态科学学报(2022年4期)2022-11-11
广西林业科学(2022年4期)2022-11-07
宝藏(2022年1期)2022-08-01
林业调查规划(2022年3期)2022-06-13
广西林业科学(2021年6期)2022-01-21
河南农业大学学报(2020年2期)2020-05-22
山西林业科技(2019年1期)2019-05-27
商周刊(2017年11期)2017-06-13
岷峨诗稿(2017年4期)2017-04-20
西北植物学报(2015年9期)2015-07-05
