
对耐力训练中“有氧”与“无氧”若干问题的重新审视
2014-10-18 10:11褚云芳陈小平
体育科学 2014年4期
褚云芳,陈小平
有氧和无氧能力是专项耐力的基础,是各个运动项目耐力训练的重要内容。研究已经证明,不同运动项目对有氧和无氧能力的需求不同,高水平专项耐力的获得不仅需要出色的有氧和无氧能力,而且,还必须形成有氧与无氧能力的最佳比例组合。当前的耐力训练,尤其是在我国的耐力训练中,对有氧或无氧单一能力的训练已有比较统一的认识和方法,而在两种能力的组合比例关系上仍然存在大量问题,这些问题直接影响到专项耐力的训练,影响到耐力性项目运动水平的提高。因此,本研究以当前耐力训练中有氧与无氧能力的关系为切入点,以其大量相关研究成果为线索,结合我国耐力训练中存在的问题,梳理和分析有氧与无氧能力对专项耐力水平的作用和影响。
1 有氧与无氧能力训练的发展
在竞技运动训练的发展过程中,人们对有氧与无氧能力及其训练,无论是在理论还是方法上都经历了一个认识和实践的过程(图1)。
在Fox等人[51]提出“能量统一体”的概念之前,世界耐力训练基本还是以训练方法的探索为主,相继出现了持续训练、法特莱克式训练和间歇训练等一系列经典的耐力训练方法[1],这些方法的出现更多的偏重于实践应用,其效果的体现也主要以优秀运动员的获胜为标志。例如,运用持续训练获得成功的芬兰中长跑运动员帕弗·努米(Paavo Nurmi)、法特莱克训练方法的创始人瑞典中长跑运动员贡德·海格(Gunder Hägg)[80]、第一个进行间歇训练的德国运动员鲁道夫·哈彼格(Rudolph Habig)以及长期接受间歇训练并获得巨大成功的捷克运动员扎托皮克(Zátopek)等[2]。20世纪60年代之后,随着测试设备和科学技术的发展,人们逐渐开始关注耐力训练能量代谢等基础性问题,其中较著名的是瑞典生理学家阿斯特拉德(Åstrand)对间歇训练方法的研究[7-9],证实了合理安排运动和休息时间能够促进机体达到最佳的生理适应,提出了间歇训练中不同运动和间歇时间对运动强度的影响,将间歇训练进一步划分为高、中、低强度三种类型。
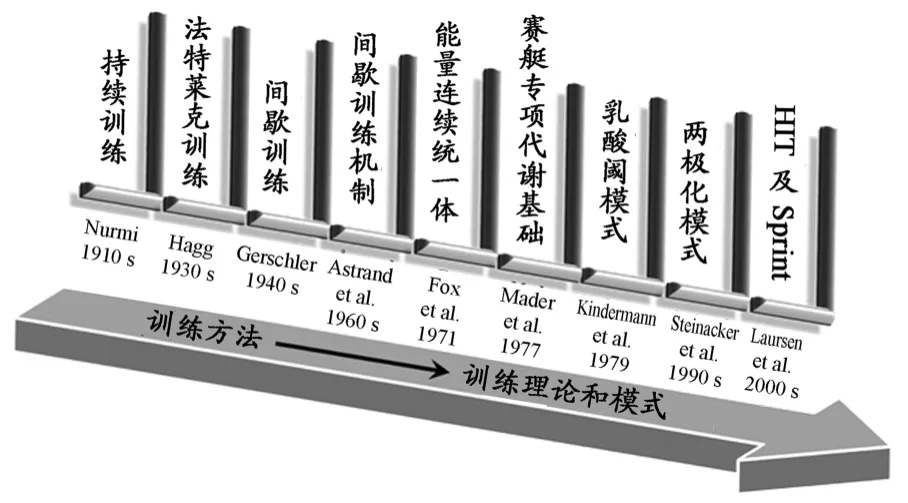
图1 耐力训练“科学化”发展进程图Figure 1.Development Process of Scientific Endurance Training
20世纪70年代之后,人们的研究视域从“方法”逐渐转向“理论”。东德的马达尔(Mader)等人[47]率先在赛艇的专项能量代谢基础研究上取得突破,他们对赛艇进行了2000m(7min)的实验室模拟测试,获取并计算了整个运动过程中耗氧量、有氧和无氧功率、血乳酸等相关数据指标,发现赛艇专项比赛中无氧乳酸供能只占约17%的份额,其余的能量供应均来自于有氧供能。该研究成果不仅改变了当时赛艇项目的训练,将以往以无氧为主的高强度耐力训练变为以有氧为主的中、低强度训练,而且,其影响逐渐辐射到其他耐力项目的训练中,与之相似的田径中长跑、自行车和游泳等项目均开始注重有氧能力的训练。对耐力训练能量代谢理论研究的另一个重要成果,是Kindermann等研究者提出的“乳酸阈”训练模式,他们的研究表明,采用有氧-无氧供能过渡区域(即血乳酸值在2~4mmol/L之间)的训练强度,能够更有效地促进有氧耐力的提高[24,30,42,46]。与此同时,Seiler等人[62]对耐力训练 的强度分布进行了梳理和总结。他们认为,在20世纪90年代后,相当数量的赛艇、自行车和马拉松等耐力项目世界优秀运动员并没有遵循Kindermann等人提出的“乳酸阈”模式进行训练,而是采用了被称为“两极化”的耐力训练模式[13,64,69,70],即低于乳酸阈强度的总训练时间或总训练距离约占75%,高于乳酸阈强度的训练约占15%~20%,而乳酸阈强度的训练份额最少,只占约5%。至此,耐力训练中的有氧训练在理论认识和实践尝试两个方面都受到世界范围内的重视,有氧训练成为这一时期的“主旋律”。
但是,随着竞技水平的快速发展,“耐力提高的瓶颈”又让人们陷入新一轮训练方法和手段的探索。20世纪90年代之后,体育科研人员又将目光重新投向了“高强度训练(High-Intensity Training,HIT)”,其研究力度和深度在20世纪60~70年代的基础上有了很大的提升。20世纪90年代至今,该类研究论文的数量上升趋势明显,尤其在2000年之后出现快速增长(图2)。总体而言,高强度训练的研究有以下发展特点:在研究对象上,20世纪90年代中期以及之前的研究成果主要来自于普通人或低运动水平 人 群的研究[20,34,37,40,65],从90年代后期开始 ,其研究逐步转向对高水平运动员的研究[14,66,72,76,77,81];在研究类型上,主要分为一次性高强度训练[73]和多次高强度间歇训练[43]的研究;在研究内容上,主要围绕一次或多次高强度运动时3个能量代谢系统的供能比例以及随着运动能力下降该 比例的变化及关系展 开讨论[12,16,29,60]。
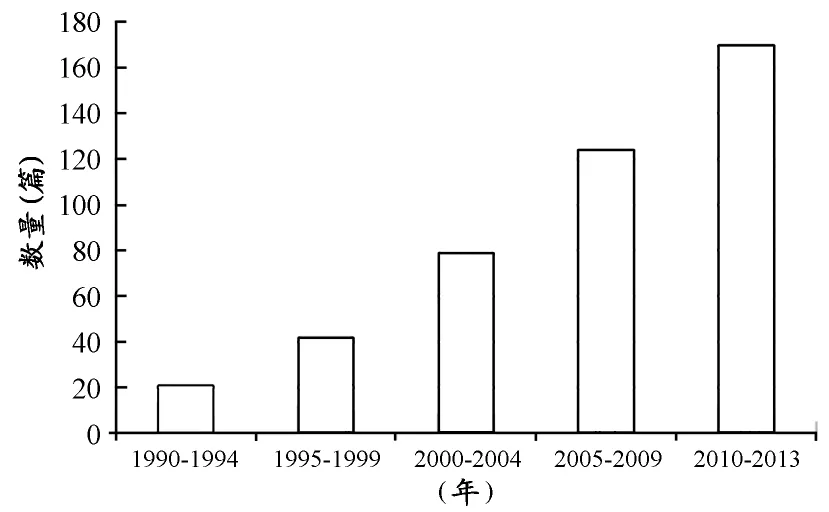
图2 “高强度训练”研究成果数量统计示意图Figure2.Quantity Statistical of Research on“High Intensity Training”
近年来,人们对耐力训练的研究逐渐向分子水平深入(图3)[44]。Burgomaster等人[19]的研究表明,机体对低强度有氧训练和高强度间歇训练所表现出的生理适应没有显著性差异,有氧耐力均能得到大幅度提高,该结果也得到其他研究的支持[32]。他们认为,在分子层面,有氧能力的提高可以通过两种训练方式(途径)来实现——低强度的有氧训练和高强度的无氧训练,这两种训练都可以促进肌纤 维 中PGC-1α蛋白含量的增 加[45,61,74],该蛋白可诱导骨骼肌线粒体的生物合成[6,71]及肌纤维类型的转化[63],同时提高机体对脂肪的利用[11],并且强烈激发骨骼肌细胞内源性GLUT4的表达[4],从而增加糖原的存储。
有氧与无氧训练在认识和实践上经历了一个不断深入并螺旋式上升的过程。有氧和无氧能力是一个能量代谢的“统一体”,对于任何一个运动项目来说,尽管由于当时的科学认识水平以及运动训练实践的发展所带来的局限性,在不同的时期对它们的关注有所不同,但无论是有氧能力还是无氧能力都不能单独成为一个项目制胜的惟一要素,形成符合专项代谢特点的有氧与无氧平衡才是耐力训练的根本任务。

图3 AMP-蛋白激酶(AMPK)和Ca-钙调蛋白激酶(CaMK)信号通路模型示意图Figure 3.Simplified Model of the Adenosine Monophosphate Kinase(AMPK)and Calcium-calmodulin Kinase(CaMK)Signaling Pathways(根据 Laursen,2010)
2 不同专项运动形式与有氧和无氧能力
运动中耗氧量的测定[38]、糖原与乳酸的关系[55-57]以及高能磷酸化合物的发现[26],使得人体在运动过程中能量供应的渠道和方式逐渐清晰。20世纪70~80年代,瑞典及挪威的生理学家阿斯特拉德(Åstrand)和罗道尔(Rodahl)基于早期的研究数据和成果,提出了人体在10s~120min全力运动中有氧和无氧的供能比例(表1)[9]。这一成果为当时的耐力训练提供了理论依据,同时,也为进一步的研究奠定了基础。随后,以福克斯(Fox)为代表的美国学者提出了能量连续统一体(energy-continuum)的概念[51],并依据运动持续时间的长短将此划分为4个不同的区域(表2)[28],从宏观上界定了不同能量供应系统在不同运动中的参与情况。至此,基于运动持续时间的不同运动项目有氧和无氧能量供应比例逐渐得到人们的认识,成为判断运动项目能量代谢特征的主要依据,并出现在大量文献、资料和教科书中[5,10,17]。

表1 最大负荷运动中供能比例一览表Table 1 Aerobic and Anaerobic Energy Supply Contributions in Maximal Exercise

表2 能量连续统一体的四个区域一览表Table 2 Four Areas of Energy Continuum
上述研究成果的生理生化依据,是人体运动时3种供能系统各自所表现出的特性。总体上,3种供能系统在运动开始阶段被同时动员,但在动员的速度、输出功率和总量方面存在明显差别。从单位时间输出功率上看,磷酸原系统在运动开始即刻就能给予肌肉最直接和高效的能量供给,其次是糖酵解系统,运动初期动员速度最慢、输出功率最小的是有氧系统。从能量输出功率的总量和供能持续时间上看,有氧系统的代谢底物存储量巨大,且不会产生有碍持续供能的代谢产物,因此,其供能持续时间最长。无氧糖酵解系统供能的同时会产生乳酸,在高强度运动下乳酸得不到及时清除,堆积造成机体酸性环境加剧(pH值下降),从而影响糖酵解供能的总量和持续时间。高能磷酸化合物在机体内存储量较少,ATP及PCr在高强度运动中仅够维持7~10s[82],供能时间最短。
在该理论背景下,我们往往会根据运动项目,尤其是体能周期类项目比赛时间的长短推断其供能特征,然后“对号入座”的指导训练。但是,这种思维模式显然没有考虑到人体运动动作结构的差异性(跑、游、骑、划、滑等)。表3列举了4个同为周期性速度耐力项目的供能比例值,尽管运动持续时间均在90~120s之间,男、女最新世界纪录(最好成绩)相差都不到7s,但其有氧与无氧供能比例却存在着显著性差异,500m皮艇和200m自由泳的有氧供能比例明显高于800m跑和1500m速度滑冰。

表3 90~120s典型动作结构项目世界纪录(最好成绩)与供能比例一览表Table 3 World Record(Best Record)and Energy Supply of Typical Movement Sports in 90~120s
上述项目呈现出不同供能比例的主要原因,可以归结为身体素质和专项技术对专项成绩具有不同的影响作用。身体素质涉及人体各种能力及其之间的匹配关系,是运动的动力来源;而专项技术涉及神经对肌肉的支配,是身体素质最佳发挥的“平台”和“杠杆”。一般来说,动作结构越简单,技术难度越低的项目,越有利于身体素质的发挥。上述4个项目中,由于游泳和皮划艇是水上项目,运动员是在不稳定的无支撑环境下进行运动,技术难度明显高于跑步和速度滑冰,所以,运动员的单位运动强度低于另外两个项目,其无氧代谢供能的比例也相应低于后者,这也许是导致其有氧与无氧供能比例产生差异的主要原因。
由此可见,相同运动时间但不同运动模式的力竭性运动对有氧和无氧能量的需求不同,技术难度以及运动工作条件是造成这种不同的主要原因。表4是赛艇水上和测功仪测试的对比研究结果[23],尽管两种测试的距离同为2000m,但由于水上划的技术难度和肌肉工作条件明显高于测功仪,因此,其单位强度也低于测功仪,在完成时间上水上划明显慢于测功仪,供能比例上无氧代谢(包括ATPCP和无氧糖酵解系统)也比测功仪低3%。

表4 不同条件下2000m划艇生理指标对比一览表Table 4 Rowing Physiological Indicators in the 2000min the Different Situations
上述研究结果对运动训练的启示:首先,不能简单地以运动持续时间作为不同项目能量代谢特征相互借鉴的依据,如不能将田径径赛项目的能量代谢特点应用于与之同样运动时间的游泳、皮划艇等水上项目。其次,在训练方法和手段上,同样不能只考虑运动的时间,如拳击、柔道和摔跤等项目经常运用与之专项比赛时间一样的跑步、自行车等训练手段发展运动员的专项耐力,显然,这样的训练由于没有充分考虑到技术和肌肉工作条件等因素,很可能达不到预期的训练效果,甚至造成训练的失误。再次,跑台和测功仪是目前竞技运动测试中经常运用的设备[15,21,67,78],它们具有规范、简单和易行等特点。但是,该类测试同样存在技术和肌肉用力方式与项目本身存在差异性的问题,所以,应将项目的技术特点纳入其能力检测和评价体系,从而更加客观地反映出专项能力水平。
3 不同运动时间、强度与有氧和无氧供能
已有研究证实,在任何运动开始时,3个能量供应系统都同时开始工作[31,35],它们各自的动员速度或供能多少主要取决于两个因素,一是各个能量系统本身所具有的供能速度,二是所进行运动的强度大小。由于前者是人体所共有的生理生化特性,因此,后者成为决定3个能量系统供能比例的主要因素。
表5是田径径赛项目供能比例与血乳酸值对应表,从供能比例相对值来看,无氧糖酵解供能居首位的是100 m,约占总供能量的70%,短或长于该项目的无氧糖酵解供能比例均出现逐步减少的趋势。该结果表明,尽管ATP-CP是100m项目最重要的能量物质,但其在肌肉中含量有限。有研究表明[29],在一个6s的全力运动中,除去极少量的有氧代谢,无氧代谢部分ATP和PCr为肌肉运动分别提供约6.3%和49.6%的能量,剩下的44.1%由糖酵解途径供能。对于田径100m这样一个10s左右的全力冲刺项目,其后程需要相当部分的无氧糖酵解来提供能量,其重要作用不容忽视[39,53,68]。从血乳酸绝对值 的角度来看,400m是产生乳酸最多的项目,可以达到24 mmol/L左右,该项目也是田径径赛项目血乳酸值的分界线,距离短或长于该项目的血乳酸绝对值均呈现逐渐降低的趋势。该结果表明,尽管短于400m的项目(如100m、200m)其糖酵解供能比例都大于400m,但由于其利用该途径供能的总时间短于400m,所以,血乳酸生成总量较少,血乳酸的绝对值较低。无氧糖酵解供能系统在大强度运动中大约可以维持45~90s,恰恰可以覆盖400m跑的全过程,而对于距离长于400m的项目,有氧供能开始成为供能的重要角色,血乳酸的绝对值和无氧糖酵解供能比例均出现下降趋势。绝对值和相对值分别从不同角度来反映运动项目的代谢特点,不能单纯凭借血乳酸的绝对值来判断无氧糖酵解供能在项目中作用的高低[33,36]。

表5 田径(径赛)项目供能比例及血乳酸值一览表Table 5 Energy Contributions and Blood Lactate of Track Sports
中、短程运动项目的距离或强度是决定能量供应,尤其是无氧无乳酸(ATP-CP)和无氧乳酸供能比例的关键因素。对于100m跑这类高强度运动项目而言,磷酸原(PCr)是最快生成ATP支持运动的供能物质,有研究显示[52],PCr能够在1.3s就达到输出峰值,然后开始下降,至5s时糖酵解供能达到峰值,并一直保持至20s左右。这说明,高强度100m跑的前程(加速段)主要的供能物质为PCr,中程和后程糖酵解乳酸供能成为主要的能量来源。400m和800m项目是典型的中距离运动项目,其既需要速度又需要速度保持能力,因此,对有氧和无氧代谢系统都有很高的要求。400m的无氧供能(包括磷酸原和糖酵解)比例高于800m,其血乳酸绝对值相比于800m也更高(表5),同时有研究显示[25],400m有氧与无氧各占50%比例的“交汇点”也明显晚于800m(图4)。这表明,400m跑不仅在糖酵解供能比例上高于800m,而且在其整个运动过程中的绝大部分时间该种供能途径占据能量输出的主导地位。另一方面,我们还应注意到,对于800m运动员而言,无氧供能主要表现在前程约1/3部分(320m左右),之后有氧代谢便开始逐渐成为能量供应的主要来源,尤其在600m之后有氧能力成为运动成绩的决定性因素。
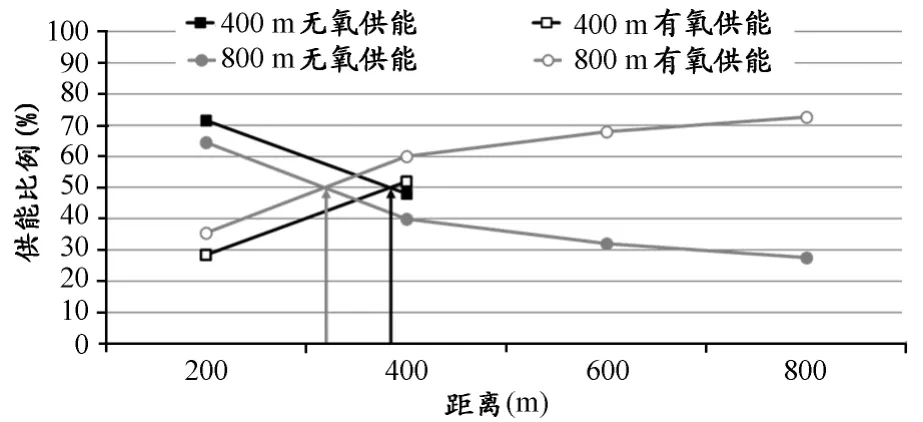
图4 400m和800m能量供应动态比例图Figure 4.Dynamic Variation of Energy Supply in 400mand 800m(根据 Duffield,2005)
上述中、短距离项目运动过程中的能量代谢特征显示,专项输出功率是决定不同能量供应途径的主要因素。短距离项目的单位输出功率高,对磷酸原和糖酵解的需求大,而随着运动距离(时间)的增加,无氧供能的比例逐渐降低,有氧代谢成为供能的主要来源。与此同时,还应该关注不同能量在运动过程中的动态变化特征,并将其作为发展某种代谢能力的训练依据。
4 有氧与无氧供能50%比例的演变及其对耐力训练的启示
自专项运动中的能量代谢问题被提出以来[27],有氧与无氧供能比例,尤其是各占50%的临界点,就成为该领域的一个广受关注并极具争议的问题。该问题不仅涉及到不同专项的能量代谢特点,而且,对不同专项耐力的训练也具有至关重要的影响。
自20世纪70年代,有氧、无氧各占50%临界点问题就受到人们的关注。最初普遍认为,该点出现在2~4min之间[41,51],随着时间的推移和研究的深入,该临界点被不断地更新[3,18,22,31,58,59]。从“临界点”问题提出至今的40 年中,有氧与无氧1∶1临界点的时间(距离)越来越短,即有氧供能比例所占份额不断增加(图5)。以田径项目为例,过去认为1500m项目的有氧与无氧供能平分秋色,之后又认为800m是有氧与无氧能量各占50%的项目,但目前最新研究认为400m才是有氧和无氧供能1∶1的分水岭。

图5 有氧-无氧供能比例50%临界点研究结果进展示意图Figure 5.Evolution of Equal Contribution from the Aerobic and Anaerobic Systems
测试设备和计算方法的不断改进和完善是造成该变化的主要原因。19世纪末,功率自行车和跑台的问世[75],使运动能力测量成为可能,经过一个多世纪的发展,各种测功仪在测试精度和测试条件控制等方面都有了快速发展,计算机技术以及遥感技术的应用,使能量代谢的检测更加便捷真实,趋于专项化。在计算方法上,从20世纪60年代就开始运用的“氧债法(Oxygen-Debt)”[79],到沿用至今的“累计氧亏法(Accumulated Oxygen-Deficit,AOD)”[54],无论是在认识上还是在计算的准确性上都有了长足的进步。
有氧与无氧50%临界点的前移,不仅揭示了运动专项的代谢特征,而且改变了人们的训练观念,其中最为重要的是提高了对有氧能力的重视程度,一些短距离(时间)的运动项目开始重视有氧能力的训练。图6是根据Maglischo于1982年、1993年和2003年分别在其专著中对游泳专项供能比例的描述绘制。从图6可知,20世纪80~90年代游泳各项目供能比例特点的变化不大,最大浮动为5%,而1993—2003年的10年中,两种代谢系统在项目中的权重出现较大变化,过去一些以无氧糖酵解能力见长的短距离项目(如50~200m项目)有氧供能比例大幅度增加。有氧比例的增长对运动训练产生了重要影响,它使得训练负荷中量与强度的安排以及相应的训练方法和手段均出现了显著变化。同时,这种变化也极大促进了专项运动水平的发展,100m和200m项目在2000年前后的变化最为明显,以自由泳为例,在1993—2003年的10年中,100m项目世界纪录男、女分别提高了0.58s和0.71s,200m项目分别提高了1.34s和0.91s,其提高幅度明显高于前一个10年。
当然,在有氧能力被普遍重视的今天,单一供能能力的强大已不足以再次冲破耐力训练的瓶颈,体育与运动科学的各种研究其最终目标都体现在运动成绩和竞技能力的提高上,有氧与无氧供能比例50%临界点的演变,不仅说明有氧能力在更多的运动项目中发挥着重要作用,同时也启示我们一些过去被认为以无氧糖酵解供能为主的运动项目需要更多的关注混合供能能力及有氧和无氧能力的平衡问题。
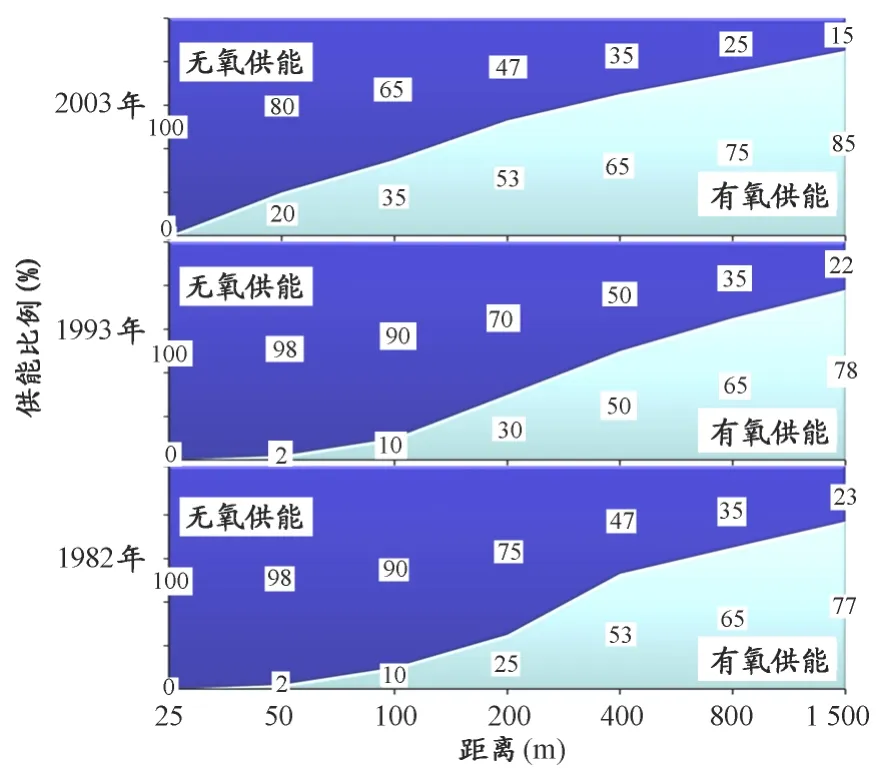
图6 游泳项目供能比例演变示意图Figure 6.Evolution of Energy Supply of Swimming(根据 Maglischo,1982、1993、2003)
5 结语
有氧与无氧能力训练的理论与实践发展及其对若干重大问题的研究与认识,不仅反映了耐力训练方法与理论不断交替发展和螺旋式上升的过程,而且,从一个侧面折射出世界竞技训练的科学化发展历程。应该深入了解并认识这个过程,同时,还应该针对我国耐力训练中存在的问题进行深刻反思,在理论和实践上紧跟世界的发展,尽快提高耐力训练水平。
[1]陈小平.当代运动训练热点问题研究:理论与实践亟待解决的问题[M].北京:北京体育大学出版社,2005:4-5.
[2]陈小平,褚云芳.田径运动训练经典理论与方法的演变与发展[J].体育科学,2013,33(4):91-封三.
[3]陈小平,梁世雷,李亮.当代运动训练理论热点问题及对我国训练实践的启示——2011杭州国际运动训练理论与实践创新论坛评述[J].体育科学,2012,32(2):3-13.
[4]宋娟,贾伟平.PGC-1α与能量平衡及糖,脂代谢的关系[J].上海医学,2004,27(11):869-871.
[5]王步标,华明.运动生理学[M].北京:高等教育出版社,2006:145-153.
[6]ADHIHETTY P J,IRRCHER I,JOSEPH A M,et al.Plasticity of skeletal muscle mitochondria in response to contractile activity[J].Experimental Physiol,2003,88(1):99-107.
[7]ÅSTRAND I,ÅSTRAND P O,CHRISTENSEN E H,et al.Intermittent muscular work[J].Acta Physiologica Scandinavica,1960,48(3-4):448-453.
[8]ÅSTRAND I,ÅSTRAND P O,CHRISTENSEN E H,et al.Myohemoglobin as an Oxygen-Store in Man[J].Acta Physiologica Scand,1960,48(3-4):454-460.
[9]ÅSTRAND P O,RODAHL K.Textbook of Work Physiology[M].New York:McGraw-Hill Book Company,1970.
[10]ÅSTRAND P O,RODAH L K,H A DAHL,et al.Textbook of Work Physiology[M].4ed.Champaign:Human Kinetics,2003.
[11]BAAR K.To perform your best:work hard not long[J].J Physiol,2006,575(3):690-690.
[12]BALSOM P D,SEGER J Y,SJODIN B,et al.Maximal-intensity intermittent exercise:effect of recovery duration[J].Int J Sports Med,1992,13(7):528-533.
[13]BILLAT V L,DEMARLE A,SLAWINSKI J,et al.Physical and training characteristics of top-class marathon runners[J].Med Sci Sports Exe,2001,33(12):2089-2097.
[14]BILLAT V L,FLECHET B,PETIT B,et al.Interval training at˙VO2max:effects on aerobic performance and overtraining markers[J].Med Sci Sports Exe,1999,31:156-163.
[15]BISHOP D.Physiological Predictors of Flat-Water Kayak Performance in Women[J].Eur J Appl Physiol,2000,82(1-2):91-97.
[16]BOGDANIS G C,NEVILL M E,BOOBIS L H,et al.Contribution of phosphocreatine and aerobic metabolism to energy supply during repeated sprint exercise[J].J Applied Physiol,1996,80(3):876-884.
[17]BOMPA T O,HAFF G G.Periodization-Theory and Methodology of Training[M].4ed.Champaigh:Human Kinetics,2009.
[18]BOUCHARD C,TAYLOR A W,SIMONEAU J A,et al.Testing Anaerobic Power and Capacity[M].In:MacDougall J D,Wenger H A,Green H J.Physiological testing of the high-performance athlete.Champaign,IL:Human Kinetics Books,1991,2:175-221.
[19]BURGOMASTER K A,HOWARTH K R,PHILLIPS S M,et al.Similar metabolic adaptations during exercise after low volume sprint interval and traditional endurance training in humans[J].J Physiol,2008,586(1):151-160.
[20]BURKER J,THAYER R,BELCAMINO M.Comparison of effects of two interval-training programmes on lactate and ventilatory thresholds[J].British J Sports Med,1994,28(1):18-21.
[21]CRAIG I S,MORGAN D W.Relationship between 800-m Running Performance and Accumulated Oxygen Deficit in Middle-Distance Runners[J].Med Sci Sports Exe,1998,30(11):1631-1636.
[22]CRAIG N P,NORTON K I,CONYERS R A,et al.Influence of Test Duration and Event Specificity on Maximal Accumulated Oxygen Deficit of High Performance Track Cyclists[J].Int J Sports Med,1995,16(8):534-540.
[23]DE CAMPOS MELLO F,DE MORAES BERTUZZI R C,GRANGEIRO P M,et al.Energy systems contributions in 2,000mrace simulation:a comparison among rowing ergometers and water[J].Eur J Appl Physiol,2009,107(5):615-619.
[24]DENIS C,DORMOIS D,LACOUR J R.Endurance Training,˙VO2max,and OBLA:A Longitudinal Study of Two Different Age Groups[J].Int J Sports Med,1984,5(4):167-173.
[25]DUFFIELD R,DAWSON B,GOODMAN C.Energy system contribution to 400-metre and 800-metre track running[J].J Sports Sci,2005,23(3):299-307.
[26]EGGLETON P,EGGLETON G P.The Inorganic Phosphate and a Labile Form of Organic Phosphate in the Gastrocnemius of the Frog[J].Biochem J,1927,21:190-195.
[27]FOX E L,ROBINSON S,WIEGMAN D L.Metabolic energy sources during continuous and interval running[J].J Appl Physiol,1969,27(2):174-178.
[28]FOX E L,MATHEWS D K.The Physiological Basis of Physical Education and Athletics[M].Philadelphia,Penn:Saunders College Pub,1981:29.
[29]GAITANOS G C,WILLIAMS C,BOOBIS L H,et al.Human muscle metabolism during intermittent maximal exercise[J].J Appl Physiol,1993,75(2):712-719.
[30]GASKILL S E,WALKER A J,SERFASS R A,et al.Changes in ventilatory threshold with exercise training in a sedentary population:the HERITAGE Family Study[J].Int J Sports Med,2001,22(8):586-592.
[31]GASTIN P B.Energy system interaction and relative contribution during maximal exercise[J].Sports Med,2001,31(10):725-741.
[32]GIBALA M J,LITTLE J P,VAN ESSEN M,et al.Short-term sprint interval versus traditional endurance training:similar initial adaptations in human skeletal muscle and exercise performance[J].J Physiol,2006,575(3):901-911.
[33]GREEN S,DAWSON B.Measurement of anaerobic capacities in humans[J].Sports Med,1993,15(5):312-327.
[34]GREEN H J,FRASER I G.Differential effects of exercise intensity on serum uric acid concentration[J].Med Sci Sports Exe,1988,20(1):55-59.
[35]HARGREAVES M,SPRIET L L.Exercise Metabolism[M].Human Kinetics Publishers,2006:8-26.
[36]HERMANSEN L,VAAGE O.Lactate disappearance and glycogen synthesis in human muscle after maximal exercise[J].Am J Physiology-Gastrointestinal Liver Physiol,1977,233(5):G422-G429.
[37]HICKSON R C,BOMZE H A,HOLLOSZY J O.Linear increase in aerobic power induced by a strenuous program of endurance exercise[J].J Appl Physiol,1977,42(3):372-376.
[38]HILL A V,LUPTON H.Muscular Exercise,Lactic Acid,and the Supply and Utilization of Oxygen[J].Oxford J Med,1923(62):135-171.
[39]JONES N L,MCCARTNEY N,GRAHAM T,et al.Muscle performance and metabolism in maximal isokinetic cycling at slow and fast speeds[J].J Appl Physiol,1985,59(1):132-136.
[40]KEITH S P,JACOBS I,MCLELLAN T M.Adaptations to training at the individual anaerobic threshold[J].Eur J Appl Physiol Occupational Physiol,1992,65(4):316-323.
[41]KEUL J.Kohlenhydrate zur Leistungsbeeinflussung in der Sportmedizin[J].Natr Metabol,1975,18 (1):157-170.
[42]KINDERMANN W,SIMON G,KEUL J.The significance of the aerobic-anaerobic determination of work load intensities during endurance training[J].Eur J Appl Physiol,1979,42:25-34.
[43]LAURSEN P B,BLANCHARD M A,JENKINS D G.Acute high-intensity interval training improves Tvent and peak power output in highly trained males[J].Canadian J Appl Physiol,2002,27(4):336-348.
[44]LAURSEN P B.Training for intense exercise performance:high‐intensity or high‐volume training?[J].Scandinavian J Med Sci Sports,2010,20(s2):1-10.
[45]LITTLE J P,SAFDAR A,BISHOP D,et al.An acute bout of high-intensity interval training increases the nuclear abundance of PGC-1αand activates mitochondrial biogenesis in human skeletal muscle[J].Am J Physiology-Regulatory,Integrative Comparative Physiol,2011,300(6):R1303-R1310.
[46]LONDEREE B R.Effect of training on lactate/ventilatory thresholds:a meta-analysis[J].Med Sci Sports Exe,1997,29(6):837-843.
[47]MADER A,HOLLMANN W.Zur Bedeutung der Stoffwechselleistungsfähigkeit des Eliteruderers im Training und Wettkampf[J].Leistungssport Suppl,1977,9:9-62.
[48]MAGLISCHO E W.Swimming Faster:a Comprehensive Guide to the Science of Swimming[M].Mountain View,CA:Mayfield Publishing Company,1982:267.
[49]MAGLISCHO E W.Swimming Even Faster[M].Mountain View,CA:Mayfield Publishing Company,1993:228.
[50]MAGLISCHO E W.Swimming Fastest:The Essential Reference on Technique,Training,and Program Design[M].Champaign,IL:Human Kinetics Publishers,2003:369.
[51]MATHEWS D K,FOX E L.The Physiological Basis of Physical Education and Athletics[M].Philadelphia,Penn:W.B.Saunders,1971:34-63.
[52]MAUGHAN R J,GLEESON M,GREENHAFF P L.Biochemistry of Exercise and Training[M].New York:Oxford University Press,1997:142-143.
[53]MCCARTNEY N,SPRIET L L,HEIGENHAUSER G J,et al.Muscle power and metabolism in maximal intermittent exercise[J].J Appl Physiol,1986,60(4):1164-1169.
[54]MEDBO J I,MOHN A C,TABATA I,et al.Anaerobic capacity determined by maximal accumulated O2deficit[J].J Appl Physiol,1988,64(1):50-60.
[55]MEYERHOF O.Die Energieumwandlungen im Muskel[J].Pflügers Archiv Eur J Physiol,1920,182(1):232-283.
[56]MEYERHOF O.Über die Energieumwandlungen im Muskel.II.Schicksal der Erholungsperiode des Muskels[J].Pflugers Arch,1920,182:284-317.
[57]MEYERHOF O.Die Energieumwandlungen im Muskel.III[J].Pflüger's Archiv Für die Gesamte Physiologie des Menschen und der Tiere,1920,185(1):11-32.
[58]MEDBØJ O N I,SEJERSTED O L E M.Acid-base and electrolyte balance after exhausting exercise in endurance-trained and sprint-trained subjects[J].Acta Physiologica Scand,1985,125(1):97-109.
[59]MORTON D P.Quantification of Anaerobic Capacity on the Swim Bench Ergometer[D].Ballarat(VIC):Ballarat University College,1992:56-58.
[60]PAROLIN M L,CHESLEY A,MATSOS M P,et al.Regulation of skeletal muscle glycogen phosphorylase and PDH during maximal intermittent exercise[J].Am J Physiology-Endocrinology Metabolism,1999,277(5):E890-E900.
[61]RUSSELL A P,FEILCHENFELDT J,SCHREIBER S,et al.Endurance training in humans leads to fiber type-specific increases in levels of peroxisome proliferator-activated receptor-γ coactivator-1and peroxisome proliferator-activated receptor-α in skeletal muscle[J].Diabetes,2003,52(12):2874-2881.
[62]SEILER K S,KJERLAND G Ø.Quantifying training intensity distribution in elite endurance athletes:is there evidence for an“optimal”distribution?[J].Scand J Med Sci Sports,2006,16(1):49-56.
[63]SELSBY J T,MORINE K J,PENDRAK K,et al.Rescue of dystrophic skeletal muscle by PGC-1αinvolves a fast to slow fiber type shift in the mdx mouse[J].PloS One,2012,7(1):e30063.
[64]SCHUMACHER Y O,MUELLER P.The 4000-m team pursuit cycling world record:theoretical and practical aspects[J].Med Sci Sports Exe,2002,34(6):1029-1036.
[65]SIMONEAU J A,LORTIE G,BOULAY M R,et al.Human skeletal muscle fiber type alteration with high-intensity intermittent training[J].Eur J Appl Physiol Occupational Physiol,1985,54(3):250-253.
[66]SMITH T P,MCNAUGHTON L R,MARSHALL K J.Effects of 4-week training using Vmax/Tmax on ˙VO2max and performance in athletes[J].Med Sci Sports Exe,1999,31(6):892-896.
[67]SPENCER M R,GASTIN P B.Energy System Contribution during 200-to 1500-m Running in Highly Trained Athletes[J].Med Sci Sports Exe,2001,33(1):157-162.
[68]SPRIET L L,SODERLUND K,BERGSTROM M,et al.Anaerobic energy release in skeletal muscle during electrical stimulation in men[J].J Appl Physiol,1987,62(2):611-615.
[69]STEINACKER J M.Physiological aspects of training in rowing[J].Int J Sports Med,1993,14:S3-10.
[70]STEINACKER J M,LORMES W,LEHMANN M,et al.Training of rowers before world championships[J].Med Sci Sports Exe,1998,30(7):1158-1163.
[71]STEINBERG G R,KEMP B E.AMPK in Health and Disease[J].Physiol Rev,2009,89(3):1025-1078.
[72]STEPTO N K,HAWLEY J A,DENNIS S C,et al.Effects of different interval-training programs on cycling time-trial performance[J].Med Sci Sports Exe,1999,31:736-741.
[73]STEPTO N K,MARTIN D T,FALLON K E,et al.Metabolic demands of intense aerobic interval training in competitive cyclists[J].Med Sci Sports Exe,2001,33(2):303-310.
[74]TERADA S,KAWANAKA K,GOTO M,et al.Effects of highintensity intermittent swimming on PGC-1αprotein expression in rat skeletal muscle[J].Acta Physiol Scand,2005,184(1):59-65.
[75]TIPTON C M.Exercise Physiology:People and Ideas[M].New York:Oxford University Press,2003:255-284.
[76]WESTON A R,MYBURGH K H,LINDSAY F H,et al.Skeletal muscle buffering capacity and endurance performance after high-intensity interval training by well-trained cyclists[J].Eur J Appl Physiol Occupational Physiol,1996,75(1):7-13.
[77]WESTGARTH T C,HAWLEY J A,RICKARD S,et al.Metabolic and performance adaptations to interval training in endurance-trained cyclists[J].Eur J Appl Physiol Occupational Physiol,1997,75(4):298-304.
[78]WEYAND P,CURCTON K,CONLEY D,et al.Percentage Anaerobic Energy Utilized During Track Running Events[J].Med Sci Sports Exe,1993,25(5):S105.
[79]WHIPP B J,SEARD C,WASSERMAN K.Oxygen deficit-oxygen debt relationships and efficiency of anaerobic work[J].J Appl Physiol,1970,28(4):452-456.
[80]WILT F.How They Train:Half Mile to Six Mile[M].Los Altos,Calif:Track and Field News,1959:23-28.
[81]ZAVORSKY G S,MONTGOMERY D L,PEARSALL D J.Effect of intense interval workouts on running economy using three recovery durations[J].Eur J Appl Physiol Occupational Physiol,1998,77(3):224-230.
[82]ZINTL F.Ausdauertraining:Grundlagen,Methoden,Trainingssteuerung[M].München:BLV,1997:103.
猜你喜欢
中老年保健(2022年3期)2022-11-21
中老年保健(2022年2期)2022-08-24
中老年保健(2022年4期)2022-08-22
中老年保健(2022年6期)2022-08-19
食品安全导刊(2021年33期)2021-11-27
阅读(科学探秘)(2020年9期)2020-11-06
家庭医学·下半月(2019年5期)2019-07-12
读与写·上旬刊(2018年1期)2018-05-06
教育教学论坛(2016年51期)2017-03-22
教育教学论坛(2016年49期)2017-02-27
