
汞在海洋浮游植物中的生物累积和毒性效应
2014-09-21 01:59王文雄
生态毒理学报 2014年5期
毋 赟,王文雄
1. 中国科学院南海海洋研究所 热带海洋生物资源与生态重点实验室,广州 510301 2. 香港科技大学生命科学学部 香港九龙清水湾
汞在海洋浮游植物中的生物累积和毒性效应
毋 赟1,王文雄2, *
1. 中国科学院南海海洋研究所 热带海洋生物资源与生态重点实验室,广州 510301 2. 香港科技大学生命科学学部 香港九龙清水湾
浮游植物是海洋生态系统的主要初级生产者,同时作为食物也是许多水生生物摄取汞的主要途径。本文综述了近年来汞在海洋浮游植物中的最新研究进展,包括汞在浮游植物中的吸收、累积规律及其影响因素,汞对浮游植物的毒性效应(生长抑制、光合作用影响)以及生物的适应机制(汞的还原、螯合解毒、矿化固定等),最后对浮游植物中汞累积和毒性的未来研究方向进行了展望。
汞;海洋浮游植物;生物累积;毒性
上世纪以来,随着工农业(如燃煤发电、冶金、电池工业、造纸业、塑料工业和化肥生产等)的快速发展,大量的含汞污染物经地表径流、大气干湿沉降、沿岸直接排放等方式进入海洋。20世纪50年代日本暴发了由于居民食用含有高浓度甲基汞的海产品导致的“水俣病”后,汞污染危害引起了全世界的震惊和广泛关注,促使人们开展汞在水体中行为及其生态毒理学的研究。
水体中的汞主要以元素汞(Hg0)、二价汞(Hg2+)和甲基汞(MeHg)形态存在。由于元素汞的强挥发性和低生物可利用性,对这种汞形态的研究主要集中于元素地球化学循环领域。在海水中,二价无机汞的浓度远高于甲基汞,如在河口沿岸地带,甲基汞只占总汞的小部分(1%~10%),但因为甲基汞具有严重的神经毒性,经生物富集和食物链传递可直接或间接地危害到所有营养级海洋生物以及人类健康[1],故两者都是生态毒理学研究的主要汞形态(下文无机汞特指二价无机汞)。
浮游植物是一类具有色素或色素体,通过光合作用制造有机物的自养型浮游生物。由于透入海水中的阳光随深度迅速衰减,浮游植物分布于整个海洋上层,并受营养盐、温度、光照等因素影响。海洋浮游植物主要包括原核生物的细菌、蓝藻和真核生物,其中,硅藻(diatom)、甲藻(dinoflagellates)和金藻(haptophytes)是海洋大陆架区域的优势种,也是温带和极地的主要赤潮种[2]。作为海洋生态系统的主要初级生产者,浮游植物也是汞进入海洋食物网的主要途径,它对汞的累积和生理毒性响应制约着整个生态系统的发展,影响着汞在海洋环境以及与之相连的大气环境中的分布、迁移和转化。海洋浮游植物具有极强的汞富集能力,通过生物浓缩作用,其汞浓度可达环境浓度的103~106倍[3]。因而开展汞在海洋浮游植物中的吸收、累积、分布、细胞归趋、转化等方面的研究,对于理解汞的生物富集和食物链传递特性、预测汞的毒性水平和生态风险有着至关重要的意义。
近年来,得益于新实验技术的推广(如放射性同位素示踪技术探索金属生物累积、差速离心法分析金属亚细胞分布、流式细胞仪技术研究细胞生理生化状态、汞形态分析技术预测汞的生物形态转化),人们对于汞在海洋浮游植物中的累积和毒性效应有了更为深入的理解。本文总结近20多年来国内外有关汞在海洋浮游植物中的研究进展,提出该领域研究中存在的问题,希望可以促进该领域的研究。
1 汞在海洋浮游植物中的累积
海洋浮游植物作为水生食物网结构和功能的主要组成部分,同时也是汞沿食物链累积和放大过程中最重要的一环,因此在过去几十年中汞在其中的生物累积一直是一个非常重要的研究领域。有关汞在海洋浮游植物中累积所需解决的关键科学问题包括:1)吸收机制;2)累积规律;3)细胞内分布。
1.1 海洋浮游植物吸收汞的基本模式
海洋浮游植物对汞的吸收途径与其水相化学形态密切相关。天然海水由于具有充足的天然有机质(dissolved organic matter, DOM),无机汞主要以Hg-DOM形态存在,而甲基汞与DOM的结合稳定常数较低(1012.6~1013.6),仍主要以氯化物形态存在[1,4]。有关海洋浮游植物对无机汞和甲基汞的吸收已有不少研究,然而目前对其吸收机制仍无定论。早期的研究认为无机汞和甲基汞都是以具有很强脂溶性的电中性化合物形态存在,因而可能通过被动扩散方式被海洋浮游植物细胞内化。Mason[5]等估算了Thalassiosira weissflogii对于无机汞和甲基汞的细胞膜渗透性(membrane permeability),发现它们与汞化合物辛醇-水分配系数(Kow)的关系很好地符合Stein[6]得到的有机化合物被动扩散经验公式。但是,无机汞的有机配体形态和甲基汞的氯化物形态具有非常相近的Kow,而无机汞的内化率又比甲基汞低,这样就很难得出无机汞和甲基汞都是以被动扩散方式进入藻细胞[7-8]。近年来,一些研究者提出甲基汞可以被浮游植物主动吸收。Moye等[9]将两种绿藻(Selenastrum Capricornutum和Cosmarium botrytis)、一种蓝绿藻(Schizothrix calcicola)和一种硅藻(Thalassiosira weissflogii)分别进行光合作用抑制剂(CCCP,DCMU,DNP和百草枯)、γ射线、暗条件等处理,发现四种浮游植物的甲基汞吸收均有显著降低,即藻类对甲基汞的吸收是一个耗能的过程,涉及主动运输。这可能是由于甲基汞与半胱氨酸形成的复合物(MeHg-L-cysteine)与蛋氨酸(L-methionine)结构相似,进而被细胞膜上的转动蛋白错误识别和运输[10]。此外,近来的研究发现无机汞可以Hg-cysteine和β-HgS形态被汞甲基化细菌吸收,而具体吸收机制及是否在浮游植物存在仍需要进一步研究[11-12]。
1.2 海洋浮游植物的汞累积及其主要影响因素
大量的汞累积研究主要通过滤网采集野外浮游植物样品或收集室内汞暴露后的藻细胞来测定汞的生物浓缩因子(bioconcentration factor, BCF)[13-14]。为了准确预测海洋浮游植物中汞的生物累积,需要深入了解汞累积规律及其影响因素。近十多年来,随着检测技术的进步以及放射性同位素标记法等的应用,浮游植物对重金属汞累积的室内研究取得了较大进展。根据描述金属在生物体内随时间的累积动力学模型[15],海洋浮游植物的金属累积可表示如下:

其中C为细胞内的金属浓度;t为暴露时间; ku为水相吸收常数; Cw为水相金属浓度; g为生长速率常数; ke为排出速率常数。
一般对于浮游植物, g>> ke[16],上式可简化为:
由该模型可知,海洋浮游植物的汞累积由水相吸收和生物生长稀释(biodilution)共同决定。其中,金属的水相吸收可通过测定较短时间内的金属累积,通过线性回归得到的斜率计算,生长率则通过长期暴露过程中细胞密度的自然对数与时间回归定量。这两个参数均受到汞本身形态、生物自身因素、环境因子等多种因素的影响。一方面,水相吸收受到上述几种因素的影响:将硅藻细胞分别进行短期(3 h)无机汞和甲基汞暴露,发现甲基汞的吸收速率是无机汞的9.2倍[8];水相吸收也与藻细胞大小密切相关,Thalassiosira pseudonana (4~6 μm)由于具有更大的细胞比表面积,较T. weissflogii (一般15 μm)而言对甲基汞的吸收速率更大(0.2 μg·L-1: 36.6 μg·(g·h)-1vs 0.1 μg·L-1: 0.8 μg·(g·h)-1)[4,8];此外,环境因子包括温度、光照、溶解有机物及其他主要离子(如H+、Cl-)等,可通过改变水相金属形态、影响浮游植物生理生化过程作用于汞的水相吸收[17]。另一方面,细胞生长同样受到上述因素的影响:产生同样生长抑制作用(IC50)所需的无机汞浓度为甲基汞浓度的59~197倍[18];同一种汞形态对不同浮游植物生长的抑制作用差别很大(无机汞在四种海洋浮游植物中的EC50值波动可达103.8)[19];并且金属对细胞的生长抑制受到环境因子的显著影响,例如温度升高增加无机汞对Scenedesmus acutus的生长抑制,而经甲基汞预暴露后的硅藻T. weissflogii对甲基汞的耐受性也显著提高[20-21]。在业已研究的各种金属中,无机汞具有较高的水相吸收速率和生物生长抑制能力,因此其在海洋浮游植物中的平均BCF值(105)高于其它金属,而甲基汞又远高于无机汞(1~2个数量级)[22-23]。值得注意的是,传统的观点认为金属的吸收率与生物生长率并不存在关联,而近来的研究指出Cd、Zn等金属的水相吸收与藻细胞生长率显著正相关,因此浮游植物的金属累积取决于水相吸收和生长率的相对变化[24-25]。
1.3 海洋浮游植物中汞的亚细胞分布
浮游植物累积的汞,主要是以与细胞内的可溶性巯基化合物结合、被矿化物固定、与膜结合等形式存在于不同的亚细胞组分或结构,进而改变汞的吸收和累积,最终对整个食物链产生影响[26]。目前最常用的定量研究金属亚细胞分布的方法是五步分离法,通过使用差速离心、热处理和化学处理,分离得到五个亚细胞组分,包括富金属矿体(metal-rich granule,MRG)、细胞碎屑(cellular debris,主要是细胞壁、膜结构)、细胞器(organelle,包括线粒体、溶酶体)、热稳定蛋白(heat-stable protein,HSP,包括植物螯合肽等)和热变性蛋白(heat-denatured protein,HDP,包括酶等蛋白质)[27-28]。在生态毒理学领域,研究者又根据各部分的生理生态意义,将这五部分划分为生物非活跃金属(金属解毒组分,biological detoxified metal,BDM)、生物活跃相金属(生物代谢活跃金属或金属敏感组分,metal sensitive fraction,MSF)、可沿食物链传递组分等(图1)[29-30]。按照这种定义,累积的汞首先进入细胞的代谢活跃相,然后进入解毒相。

图1 五种亚细胞组分Fig. 1 Five subcellular fractions
根据上述方法,Wu和Wang[8]结合放射性同位素示踪技术测定了汞在海洋浮游植物中的亚细胞分布:经过72 h无机汞暴露,硅藻T. pseudonana中高达38%~56%的细胞内汞结合在细胞碎屑部分,而在绿藻Chlorella autotrophica和金藻Isochrysis galbana中,更多汞被分布在热稳定蛋白组分(> 67%)。不同于无机汞,与热稳定蛋白结合是甲基汞在所有三种海洋浮游植物中的最主要储存方式(> 44%)。结果表明,汞的亚细胞分布受其自身形态和生物种类的影响,细胞碎屑是无机汞在海洋浮游植物中的重要存储位点,而热稳定蛋白对于无机汞和甲基汞而言都是重要的结合组分。在这项研究中,亚细胞金属也被探索用于预测汞的生物毒性。汞的生物毒性具有物种特异性,不同浮游植物用IC50表达金属毒性的差别为1.8倍,而无机汞和甲基汞分别用HSP/BDM和organalles/MSF/MRG中浓度表达时,汞的种间毒性差异最小(1.4倍和1.6倍),说明汞的亚细胞分布与金属毒性密切相关,可以很好地解释或预测汞的毒性。然而,金属在不同亚细胞组分的分布并不是处于静止或平衡状态,而是不断变化的。Ng和Wang[26]对贻贝的研究结果表明:开始暴露时Cd、Ag和Zn的吸收速率与其在热敏感蛋白中的分布呈正相关,随着暴露时间的延长,金属从金属敏感组分再分配到热稳定蛋白组分,且金属从这些亚细胞组分排出的速率也不相同。目前还没有研究报道汞在海洋浮游植物中亚细胞分布的动力学变化,以及这种变化在汞的生物累积、毒性效应中所起的作用。
2 汞对海洋浮游植物的毒性效应
汞的毒理学研究已有很长的历史。已有的研究表明汞对海洋浮游植物的生理毒性效应具有非特异性:汞对巯基(—SH)有很强的亲和力,通过与酶等蛋白质中的—SH结合,可导致酶活丧失、蛋白质变性、DNA结构改变等;汞也会取代必需金属(如叶绿素中心原子Mg)、诱导细胞内活性自由基产生(如H2O2等),导致蛋白质功能改变、膜脂过氧化、DNA损伤等[31-33]。分子生化水平的毒性表现在细胞、个体水平,即为叶绿素含量降低、光合系统损伤、代谢过程紊乱、生长发育受阻、细胞膜结构破坏甚至衰老死亡[34-35],其中生物生长与光合作用参数由于生态意义重大且方便易测,是目前最常使用的毒理学测试指标。
2.1 汞对海洋浮游植物的生长抑制效应
海洋浮游植物由于其广泛存在性和生态重要性,被美国材料与试验协会(American Society for Testing and Materials)作为评价污染物毒性水平的模式生物[36]。从1970s以来,大量的毒性测试以生长抑制的50%剂量(IC50)为效应终端,研究了汞对海洋浮游植物的急性毒性效应[5,18,37-38]。在之前的研究中,细胞密度的测定主要采用血球计数板或库尔特粒度仪(Coulter counter),然而,这些方法将进行生长试验的所有藻细胞个体行为等同于其所表现出来的种群生长行为,忽视了污染物可能对浮游植物生长抑制影响方式的不同[39-40]:导致细胞死亡或抑制细胞分裂。2012年,Wu等[41]通过引入荧光染色和流式细胞仪分析技术,将活细胞和死亡细胞区分开来,揭示了无机汞和甲基汞对生长率抑制的不同作用方式:无机汞暴露导致T. weissflogii分化为两个亚种群,一个亚种群的细胞膜结构遭到破坏,细胞死亡,另一个亚种群的生长则未受到抑制。而甲基汞的暴露并未导致种群的分化,所有细胞具有相似的分裂速率,但与对照组相比显著降低[41]。这种现象的一种可能解释是不同的汞形态具有不同的作用位点:无机汞主要作用于原生质的细胞膜,而甲基汞则主要破坏细胞质组织的完整性和影响新陈代谢,这也与它们的亚细胞分布模式相一致。
2.2 汞对海洋浮游植物光合作用的影响
光合作用是地球上最重要的一种能量转化过程,海洋浮游植物的光合作用维系着整个生态系统的正常运转,因此也是研究最多的污染物毒性效应指标。汞对海洋浮游植物光合作用的影响主要表现在:1)干扰叶绿素的合成,破坏叶绿体膜结构。经无机汞和甲基汞处理后,硅藻T. weissflogii的叶绿素含量显著降低[41]。汞使叶绿素含量下降原因可能有二:一是它抑制原叶绿素酸酯酶活性、影响氨基-γ-酮戊酸的合成,直接干扰叶绿素的生物合成[42-43];二是作用于叶绿体膜,破坏了叶绿体的完整结构。2)抑制光合电子传递链。目前大多数研究通过使用脉冲调制叶绿素荧光仪(pulse-amplitude-modulated fluorescence,PAM),报道了无机汞在光合电子传递过程中的作用位点:包括光合系统II(photosystem II,PSII)的电子供体端、电子受体端、PSII反应中心蛋白,光合系统I(photosystem I,PSI)的电子供体端、电子受体端、PSI反应中心,还有类囊体膜上的捕光复合物II[44-48]。这些研究结果说明无机汞的抑制是多位点、全电子传递链的。而有关甲基汞对光合电子传递作用的研究较少,有研究认为甲基汞影响PSII电子受体端、PSII反应中心D1蛋白[49-50]。PAM测定虽然具有生物学上的意义,却耗时且并不灵敏,不能准确反映金属对细胞的毒性效应。近年来,一种更为快捷灵敏、可视化的方法,即双光子激发荧光寿命成像技术(two-photon excitation fluorescence lifetime imaging microscopy)被初步尝试运用于估测污染物的光合毒性[51-52]。利用这种技术,Wu等[41]比较了无机汞和甲基汞对T. weissflogii全电子传递链的影响,发现无机汞在最低无抑制浓度(no observed effect concentration)时即可导致非光化学淬灭增加,叶绿素荧光寿命延长,证明了无机汞对光合电子传递的抑制作用;而甲基汞即使在细胞完全不能生长的浓度下对叶绿素的荧光寿命也并无影响,即甲基汞与无机汞的光合作用位点并不相同。这同样可能是由于膜结构是无机汞细胞内的主要分布位置和作用位点,因而类囊体膜上的电子传递链更易被无机汞显著影响。
2.3 海洋浮游植物对汞的生理生化耐受机制
为适应环境中不断升高的汞浓度,生物在长期进化过程中形成了独特的汞耐受机制。目前,对于海洋浮游植物的汞适应途径和机制的了解主要受细菌和高等植物研究的启发,包括浓度(减少汞累积)和形态(生物转化、胞内隔离)的调控。前者通过减少汞吸收和增加汞排出实现,后者则是通过生物螯合、固定、隔离等方式降低汞的生物毒性。海洋浮游植物细胞对汞的生理生化耐受机制如图2所示。
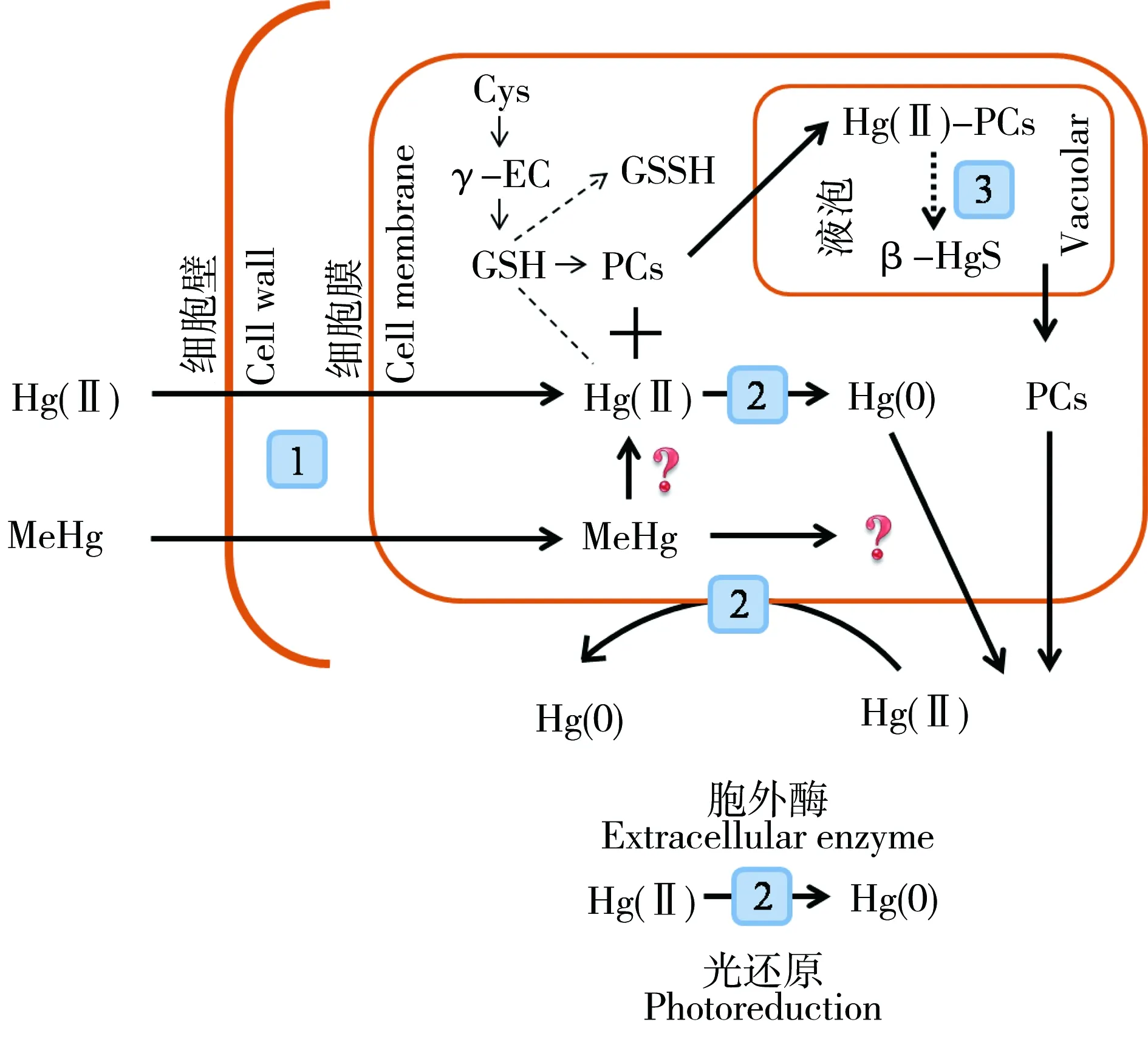
图2 海洋浮游植物对汞的生理生化适应机制:1)汞吸收进入藻细胞;2)细胞外、细胞内、细胞膜上酶催化的二价无机汞还原;3)无机汞被巯基固定并进一步形成β-HgSFig. 2 the adaptation of marine phytoplankton to environmental mercury: 1) mercury uptake into algal cells; 2) enzymatically mediated Hg(II) reduction; 3) β-HgS formation via-SH complex
在汞的生物地球化学循环研究中,人们很早就发现了表面水的溶解性气态汞(dissolved gaseous mercury,DGM,主要为元素汞)挥发是一种重要的汞的自然来源。Mason和Sheu[53]评估了海洋在全球汞循环中的作用,指出每年从海洋挥发进入大气的汞与人为过程排放的汞大致相当。早期的研究认为这种DGM的产生主要是光化学反应和细菌活动共同作用的结果。1975年,Ben-Bassat和Mayer[54]首次证实了绿藻Chlorella暴露于μmol·L-1无机汞能够产生DGM,随后的大量现场调查和室内试验均证实,真核藻类可以将二价无机汞还原为低毒、高挥发性的DGM,从而降低无机汞的生物可利用性[55-56]。Wu和Wang[57]在最新的一篇文章中系统地研究了不同浮游植物种类(T. weissflogii、C. autotrophica和I. galbana)的汞还原能力。通过测定暗处理十分钟内产生的元素汞,他们发现所研究的三种海洋浮游植物都能够将无机汞转化为DGM,且转化能力随汞浓度的升高而增加,但一般小于添加无机汞浓度的5% d-1。使用间接评估法(质量平衡法)进一步证实了这一结论:72 h内产生的DGM不超过添加无机汞浓度的30%。无机汞被藻类转化的可能机制包括胞外分泌酶、细胞表面酶过程和胞内还原过程等。对于甲基汞,一些汞耐受细菌可以利用有机汞裂解酶将甲基汞的C-Hg键切断,再对生成的无机汞进行还原解毒[58],然而这种甲基汞的转化途径目前尚未在真核浮游植物中发现。此外,汞也可通过影响细胞膜上的磷脂改变细胞膜的通透性,进而改变汞的吸收和排出[59]。
汞进入细胞后,可诱导一类富含半胱氨酸(cysteine,Cys)的多肽—植物螯合肽(phytochelatins,PCs)的合成。PCs的主要作用是维持细胞内必需金属平衡和缓解非必需金属毒性[60]。此外,PCs还可作为胞外分泌物改变水相的重金属形态,进而影响重金属的生物可利用性[61]。PCs于1985年最先在Cd处理过的印度萝芙木(Rauvolfia serpentina)悬浮培养细胞中被发现,其结构通式为 (γ-Glu-Cys)n-Gly (n=2~11)[62]。现在较为普遍的PCs测定方法是使用monobromobimane荧光标记-HPLC分离检测。之前的研究早已发现,无机汞可以诱导浮游植物中PCs的合成[63]。Wu和Wang[8]结合汞的累积动力学,全面研究了浮游植物中PCs的诱导规律,特别是对比了无机汞和甲基汞的差异:无机汞的PCs诱导是一个非常灵敏的过程,将T. weissflogii暴露于1.1 μmol·L-1无机汞,30 min 后即可检测到PC2-3及其前体化合物Cys、谷氨酰半胱氨酸(γ-EC)的显著诱导,12 h后又发现了PC4的合成,符合PCs的合成机制(PCn以PCn-1为底物进行肽链延伸)。甲基汞对PCs的诱导能力远低于无机汞,与无机汞产生相同生长抑制作用(40%)的甲基汞浓度(23 n mol·L-196 h只能够显著诱导GSH的合成,而另一项研究将T. weissflogii暴露于2 n mol·L-1甲基汞18 d后也只检测到PC2的诱导响应[21]。这可能是由两方面原因导致:一是甲基汞只能与一个PCs-SH结合,其强度低于无机汞与两个-SH的结合;二是甲基汞只能通过去甲基化产生的无机汞间接诱导PCs的生物合成[64]。因而依照所测得的PCs浓度和汞生物累积浓度,只估计了无机汞的细胞归趋:T. weissflogii 96 h暴露期内,PC-SH : 细胞内汞为0.1~0.3,远低于胞外的饱和结合比2;三种海洋浮游植物的研究中也发现Hg-PCs复合物占细胞累积汞浓度< 20%,说明无机汞的生物解毒过程中还存在其他机制或者PCs复合物只是汞解毒过程中的中间形态[57]。
金属与PCs结合形成的低分子量复合物,可在液泡膜金属转运蛋白的作用下,进入液泡形成高分子量巯基复合物,并与液泡内的金属(独立进入液泡)、硫(H2S形成)作用最终被生物矿化固定[65]。早期的研究揭示了CdS晶体的存在,但由于检测方法的限制,有关汞的生物矿化作用并不明了,其在汞的生物解毒过程中的相对重要性也并不知晓。近几年,Kelly等[66]改进了无机汞的传统冷原子荧光检测法,发现碱还原可以检测除有机汞外的所有汞形态,而酸还原则无法检测有机汞和β-HgS,利用这两种测定方法的差值,他们首次定量了浮游植物中的β-HgS化合物,并认为这是实验室培养的原核和真核藻细胞内汞存在的主要形态(20%~90%)。Carrasco-Gil等[67]又通过扩展X射线吸收精细结构谱(Extended X-ray Absorption Fine Structure,EXAFS),证实了β-HgS在从汞污染土壤中采集的Horehound组织中的大量存在(28%~55%)。在此基础上,Wu和Wang[57]研究了无机汞在几种海洋浮游植物中的生物转化,同样发现β-HgS是藻细胞内最主要的汞累积形态(20%~90%),且β-HgS占细胞内累积汞的比例与藻细胞生长率抑制呈负相关关系,由此说明合成β-HgS是浮游植物适应无机汞胁迫的重要机制。大量的β-HgS形成也与之前所述的无机汞的亚细胞分布相一致,因为β-HgS可在细胞壁上形成单晶或多晶颗粒,经五步分离法后进入细胞壁所在的细胞碎屑,故细胞碎屑是无机汞结合的主要亚细胞组分。酸碱还原法测定汞的矿化作用目前刚刚起步,由于缺乏纯化步骤,可能高估β-HgS的作用(包含一些低分子量巯基化合物),有待技术的进一步提高,有条件的话可以使用较为昂贵的EXAFS法。而有关甲基汞最终能否转化为硫化物晶体的研究目前尚属空白。
3 总结与展望
汞是少数具有生物放大能力的金属,但一般认为无机汞不会被生物放大,而甲基汞则存在生物放大现象[68]。对海洋浮游植物的研究从食物链底端生物对不同汞形态的食物链传递差异提供了一定解释:1)甲基汞比无机汞更容易在海洋浮游植物中累积(甲基汞的水相吸收远高于无机汞,相同浓度下对浮游植物的生长抑制能力更强);2)甲基汞比无机汞更多地分布在可沿食物链传递部分,具有较大的食物链放大潜能。3)汞的食物链传递受浮游植物种类、密度影响[69-70]。但是,目前对汞在海洋浮游植物中研究的系统性和全面性都有待加强,今后应从以下几个方向深入开展:
(1)汞在海洋浮游植物中的累积规律。前人对浮游植物的汞累积在实验室和野外都做了大量的工作,但仍然缺乏汞水相吸收和生物生长速率这两个决定汞累积的重要动力学参数方面的研究。
(2)浮游植物致毒汞水平的研究。目前的研究认为金属的毒性不仅决定于其生物累积量,也取决于其亚细胞分布[71-72],如何有效地结合这些理论和方法评估汞的毒性仍是一个很大的挑战。
(3)汞在海洋浮游植物体内的转化及对食物链高营养级生物的影响。汞在细菌细胞内的生物转化研究较多,而在海洋浮游植物中的研究尚处于起步阶段,且其转化机理仍不明确。
(4)室内模拟实验与现场观测试验的有机结合。目前有关浮游植物对汞累积和毒性响应的研究,多在实验室内进行,处理时间较短、暴露浓度较高,在今后的工作中有必要加强真实环境浓度条件下汞对海洋浮游植物的长期效应研究。
另外,在海洋污染日趋严重的情况下,仅仅从单一汞污染研究得到的结论并不能解决我们遇到的实际问题,复合污染下汞的累积和毒性研究将成为生态毒理学领域的一个重要课题。
[1] Fitzgerald W F, Lamborg C H, Hammerschmidt C R. Marine biogeochemical cycling of mercury [J]. Chemical Reviews, 2007, 107(2): 641-662
[2] Simon N, Cras A L, Foulon E, et al. Diversity and evolution of marine phytoplankton [J]. Comptes Rendus Biologies, 2009, 332(2-3): 159-170
[3] Pickhardt P C, Fisher N S. Accumulation of inorganic and methylmercury by freshwater phytoplankton in two contrasting water bodies [J]. Environmental Science & Technology, 2007, 41(1): 125-131
[4] Zhong H, Wang W X. Controls of dissolved organic matter and chloride on mercury uptake by a marine diatom [J]. Environmental Science & Technology, 2009, 43(23): 8998-9003
[5] Mason R P, Reinfelder J R, Morel F M M. Uptake, toxicity, and trophic transfer of mercury in a coastal diatom [J]. Environmental Science & Technology, 1996, 30(6): 1835-1845
[6] Stein W. Transport and Diffusion Across Cell Membranes [M]. Elsevier, 1986: 1-68
[7] Benoit J M, Mason R P, Gilmour C C, et al. Constants for mercury binding by dissolved organic matter isolates from the Florida Everglades [J]. Geochimica et Cosmochimica Acta, 2001, 65(24): 4445-4451
[8] Wu Y, Wang W X. Thiol compounds induction kinetics in marine phytoplankton during and after mercury exposure [J]. Journal of Hazardous Materials, 2012, 217: 271-278
[9] Moye H A, Miles C J, Phlips E J, et al. Kinetics and uptake mechanisms for monomethylmercury between freshwater algae and water [J]. Environmental Science & Technology, 2002, 36(16): 3550-3555
[10] Yin Z, Jiang H, Syversen T, et al. The methylmercury-l-cysteine conjugate is a substrate for the L-type large neutral amino acid transporter [J]. Journal of Neurochemistry, 2008, 107(4): 1083-1090
[11] Schaefer J K, Morel F M M. High methylation rates of mercury bound to cysteine by Geobacter sulfurreducens [J]. Nature Geoscience, 2009, 2(2): 123-126
[12] Graham A M, Aiken G R, Gilmour C C. Dissolved organic matter enhances microbial mercury methylation under sulfidic conditions [J]. Environmental Science & Technology, 2012, 46(5): 2715-2723
[13] Di Beneditto A P M, Bittar V T, Camargo P B, et al. Mercury and nitrogen isotope in a marine species from a tropical coastal food web [J]. Archives of Environmental Contamination and Toxicology, 2012, 62(2): 264-271
[14] Mason R P, Choi A L, Fitzgerald W F, et al. Mercury biogeochemical cycling in the ocean and policy implications [J]. Environmental Research, 2012, 119: 101-117
[15] Luoma S N, Rainbow P S. Why is metal bioaccumulation so variable? biodynamics as a unifying concept [J]. Environmental Science & Technology, 2005, 39(7): 1921-1931
[16] Sunda W G, Huntsman S A. Processes regulating cellular metal accumulation and physiological effects: phytoplankton as model systems [J]. Science of the Total Environment, 1998, 219(2): 165-181
[17] Luengen A C, Fisher N S, Bergamaschi B A. Dissolved organic matter reduces algal accumulation of methylmercury [J]. Environmental Toxicology and Chemistry, 2012, 31(8): 1712-1719
[18] Wu Y, Wang W X. Accumulation, subcellular distribution and toxicity of inorganic mercury and methylmercury in marine phytoplankton [J]. Environmental Pollution, 2011, 159(10): 3097-3105
[19] Fisher N S, Bohe M, Teyssie J L. Accumulation and toxicity of Cd, Zn, Ag, and Hg in four marine phytoplankters [J]. Marine Ecology Progress Series, 1984, 19(3): 201-213
[20] Huisman J, Ten Hoopen H J G, Fuchs A. The effect of temperature upon the toxicity of mercuric chloride to Scenedesmus acutus [J]. Environmental Pollution Series A, Ecological and Biological, 1980, 22(2): 133-148
[21] Wu Y, Wang W X. Differential acclimation of a marine diatom to inorganic mercury and methylmercury exposure [J]. Aquatic Toxicology, 2013, 138: 52-59
[22] 王文雄. 微量金属生态毒理学和生物地球化学[M]. 科学出版社, 2011: 120-121
[23] IAEA (International Atomic Energy Agency). Sediment distribution coefficients and concentration factors for biota in the marine environment [J]. Technical Report Series, 2004, No. 422
[24] Miao A J, Wang W X. Relationships between cell-specific growth rate and uptake rate of cadmium and zinc by a coastal diatom [J]. Marine Ecology Progress Series, 2004, 275: 103-113
[25] Wang W X, Dei R C H, Hong H. Seasonal study on the Cd, Se, and Zn uptake by natural coastal phytoplankton assemblages [J]. Environmental Toxicology and Chemistry, 2005, 24(1): 161-169
[26] Ng T Y, Wang W X. Dynamics of metal subcellular distribution and its relationship with metal uptake in marine mussels [J]. Environmental Toxicology and Chemistry, 2005, 24(9): 2365-2372
[27] Wallace W G, Lee B G, Luoma S N. Subcellular compartmentalization of Cd and Zn in two bivalves. I. significance of metal-sensitive fractions (MSF) and biologically detoxified metal (BDM) [J]. Marine Ecology Progress Series, 2003, 249: 183-197
[28] Lavoie M, Bernier J, Fortin C, et al. Cell homogenization and subcellular fractionation in two phytoplanktonic algae: Implications for the assessment of metal subcellular distributions [J]. Limnology and Oceanography: Methods, 2009, 7: 277-286
[29] Lavoie M, Le Faucheur S, Fortin C, et al. Cadmium detoxification strategies in two phytoplankton species: Metal binding by newly synthesized thiolated peptides and metal sequestration in granules [J]. Aquatic Toxicology, 2009, 92(2): 65-75
[30] Rainbow P S. Trace metal concentrations in aquatic invertebrates: Why and so what [J]. Environmental Pollution, 2002, 120(3): 497-507
[31] Nahar S, Tajmir-Riahi H A. Do metal ions alter the protein secondary structure of a light-harvesting complex of thylakoid membranes? [J]. Journal of Inorganic Biochemistry, 1995, 58(3): 223-234
[32] Satoh M, Matsumoto Y. Mercury-induced oxidative stress in marine phytoplankton Tetraselmis tetrathele (Prasinophyceae) [J]. Natural Science Research, Faculty of Integrated Arts and Sciences, The University of Tokushima, 2008, 22: 57-63
[33] Patra M, Sharma A. Mercury toxicity in plants [J]. The Botanical Review, 2000, 66(3): 379-422
[34] Graevskaya E E, Antal T K, Matorin D N, et al. Evaluation of diatomea algae Thalassiosira weissflogii sensitivity to chloride mercury and methylmercury by chlorophyll fluorescence analysis [C]. Journal de Physique IV (Proceedings), EDP sciences, 2003, 107: 569-572
[35] Singh D P, Khare P, Bisen P S. Effect of Ni2+, Hg2+and Cu2+on growth, oxygen evolution and photosynthetic electron transport in Cylindrospermum IU 942 [J]. Journal of Plant Physiology, 1989, 134(4): 406-412
[36] ASTM. Standard Guide for Conducting Static Toxicity Tests with Microalgae [S]. American Society for Testing and Materials, Philadelphia, PA, 2004, ASTM standard No. E1218-04
[37] Nuzzi R. Toxicity of mercury to phytoplankton [J]. Nature, 1972: 38-40
[39] Wiacek C, Müller S, Benndorf D. A cytomic approach reveals population heterogeneity of Cupriavidus necator in response to harmful phenol concentrations [J]. Proteomics, 2006, 6(22): 5983-5994
[40] Czechowska K, van der Meer J R. A flow cytometry based oligotrophic pollutant exposure test to detect bacterial growth inhibition and cell injury [J]. Environmental Science & Technology, 2011, 45(13): 5820-5827
[41] Wu Y, Zeng Y, Qu J Y, et al. Mercury effects on Thalassiosira weissflogii: Applications of two-photon excitation chlorophyll fluorescence lifetime imaging and flow cytometry [J]. Aquatic Toxicology, 2012, 110: 133-140
[42] Lenti K, Fodor F, Böddi B. Mercury inhibits the activity of the NADPH: Protochlorophyllide oxidoreductase (POR) [J]. Photosynthetica, 2002, 40(1): 145-151
[43] Gupta P, Jain M, Sarangthem J, et al. Inhibition of 5-aminolevulinic acid dehydratase by mercury in excised greening maize leaf segments [J]. Plant Physiology and Biochemistry, 2013, 62: 63-69
[44] Bernier M, Popovic R, Carpentier R. Mercury inhibition at the donor side of photosystem II is reversed by chloride [J]. FEBS Letters, 1993, 321(1): 19-23
[45] Prokowski Z. Effects of HgCl2on long-lived delayed luminescence in Scenedesmus quadricauda [J]. Photosynthetica, 1993, 28
[46] Murthy S D S, Mohanty N, Mohanty P. Prolonged incubation with low concentrations of mercury alters energy transfer and chlorophyll (Chl) a protein complexes in Synechococcus 6301: Changes in Chl a absorption and emission characteristics and loss of the F695 emission band [J]. Biometals, 1995, 8(3): 237-242
[47] Mi H, Klughammer C, Schreiber U. Light-induced dynamic changes of NADPH fluorescence in Synechocystis PCC 6803 and its ndhB-defective mutant M55 [J]. Plant and Cell Physiology, 2000, 41(10): 1129-1135
[49] Antal T K, Graevskaya E E, Matorin D N, et al. Assessment of the effects of methylmercury and copper ions on primary processes of photosynthesis in green microalga Chlamydomonas moewusii by analysis of the kinetic curves of variable chlorophyll fluorescence [J]. Biophysics, 2009, 54(4): 481-485
[50] Matorin D N, Osipov V A, Seifullina N K, et al. Increased toxic effect of methylmercury on Chlorella vulgaris under high light and cold stress conditions [J]. Microbiology, 2009, 78(3): 321-327
[51] Broess K, Borst J, Amerongen H. Applying two-photon excitation fluorescence lifetime imaging microscopy to study photosynthesis in plant leaves [J]. Photosynthesis Research, 2009, 100(2): 89-96
[52] Zeng Y, Wu Y, Li, D, et al. Two-photon excitation chlorophyll fluorescence lifetime imaging: A rapid and noninvasive method for in vivo assessment of cadmium toxicity in a marine diatom Thalassiosira weissflogii [J]. Planta, 2012, 236(5): 1653-1663
[53] Mason R P, Sheu G. Role of the ocean in the global mercury cycle [J]. Global Biogeochemical Cycles, 2002, 16(4): 40-41
[54] Ben-Bassat D, Mayer A M. Volatilization of mercury by algae [J]. Physiologia Plantarum, 1975, 33(2): 128-132
[55] Mason R P, Morel F M M, Hemond H F. The Role of Microorganisms in Elemental Mercury Formation in Natural Waters [M]. Mercury as a Global Pollutant. Springer Netherlands, 1995: 775-787
[56] Morelli E, Ferrara R, Bellini B, et al. Changes in the non-protein thiol pool and production of dissolved gaseous mercury in the marine diatom Thalassiosira weissflogii under mercury exposure [J]. Science of the Total Environment, 2009, 408(2): 286-293
[57] Wu Y, Wang W-X. Intracellular speciation and transformation of inorganic mercury in marine phytoplankton [J]. Aquatic Toxicology, 2014, 148: 122-129.
[58] Robinson J B, Tuovinen O H. Mechanisms of microbial resistance and detoxification of mercury and organomercury compounds: Physiological, biochemical, and genetic analyses [J]. Microbiological Reviews, 1984, 48(2): 95
[59] Vallee B L, Ulmer D D. Biochemical effects of mercury, cadmium, and lead [J]. Annual Review of Biochemistry, 1972, 41(1): 91-128
[60] Kneer R, Zenk M H. The formation of Cd-phytochelatin complexes in plant cell cultures [J]. Phytochemistry, 1997, 44(1): 69-74
[61] Lee J G, Ahner B A, Morel F M M. Export of cadmium and phytochelatin by the marine diatom Thalassiosira weissflogii [J]. Environmental science & technology, 1996, 30(6): 1814-1821
[62] Grill E, Winnacker E L, Zenk M H. Phytochelatins: The principal heavy-metal complexing peptides of higher plants [J]. Science, 1985, 230(4726): 674-676
[63] Ahner B A, Morel F M M. Phytochelatin production in marine algae. 2. induction by various metals [J]. Limnology and Oceanography, 1995, 40: 658
[64] Krupp E M, Mestrot A, Wielgus J, et al. The molecular form of mercury in biota: Identification of novel mercury peptide complexes in plants [J]. Chemical Communications, 2009(28): 4257-4259
[65] Aiking H, Govers H, Van't Riet J. Detoxification of mercury, cadmium, and lead in Klebsiella aerogenes NCTC 418 growing in continuous culture [J]. Applied and Environmental Microbiology, 1985, 50(5): 1262-1267
[66] Kelly D, Budd K, Lefebvre D D. Mercury analysis of acid- and alkaline-reduced biological samples: identification of meta-cinnabar as the major biotransformed compound in algae [J]. Applied and Environmental Microbiology, 2006, 72(1): 361-367
[67] Carrasco-Gil S, Siebner H, LeDuc D L, et al. Mercury localization and speciation in plants grown hydroponically or in a natural environment [J]. Environmental Science & Technology, 2013, 47(7): 3082-3090
[68] Wang W X. Interactions of trace metals and different marine food chains [J]. Marine Ecology Progress Series, 2002, 243: 295-309
[69] Chen C, Kamman N, Williams J, et al. Spatial and temporal variation in mercury bioaccumulation by zooplankton in Lake Champlain (North America) [J]. Environmental Pollution, 2012, 161: 343-349
[70] Faucheur S L, Campbell P G C, Fortin C, et al. Interactions between mercury and phytoplankton: Speciation, bioavailability and internal handling [J]. Environmental Toxicology and Chemistry, 2013
[71] Di Toro D M, Allen H E, Bergman H L, et al. Biotic ligand model of the acute toxicity of metals. 1. Technical basis [J]. Environmental Toxicology and Chemistry, 2001, 20(10): 2383-2396
[72] Wang M, Wang W-X. Cadmium toxicity in a marine diatom as predicted by the cellular metal sensitive fraction [J]. Environmental Science & Technology, 2008, 42(3): 940-946
◆
BioaccumulationandToxicityofMercuryinMarinePhytoplankton
Wu Yun1, Wang Wenxiong2,*
1. Key Laboratory of Tropical Marine Bio-resources and Ecology, Chinese Academy of Sciences, Guangzhou 510301, China2. Division of Life Science, The Hong Kong University of Science and Technology, Clear Water Bay, Kowloon, Hong Kong, China
7 May 2014accepted16 June 2014
Phytoplankton is the primary producer of marine ecosystem, and are also a source of mercury accumulation in many aquatic animals. This paper reviews the accumulation, toxicity and transformation of two major mercury species (inorganic mercury and methylmercury) in marine phytoplankton. Based on the biokinetic model, mercury accumulation in phytoplankton is a combined result of mercury uptake and growth dilution, which are dependent on the algal species, metal speciation and environmental factors. Accumulated mercury is then transported into different subcellular pools where mercury may produce toxicity or be detoxified by biotransformation (including mercury reduction, chelation and mineralization). Finally, perspectives for future research in the area of mercury-phytoplankton interactions are discussed.
mercury; marine phytoplankton; bioaccumulation; toxicity
国家重点基础研究发展计划(No. 2013CB430004)
毋赟(1987-),女,博士,助理研究员,研究方向为生态毒理学,E-mail: lanzhihe322@gmail.com;
*通讯作者(Corresponding author),E-mail: wwang@ust.hk
10.7524/AJE.1673-5897-20140507002
2014-05-07录用日期:2014-06-16
1673-5897(2014)5-810-09
: X171.5
: A
王文雄(1965—),讲座教授,长期从事金属生态毒理学研究。已发表SCI论文300多篇。
毋 赟, 王文雄. 汞在海洋浮游植物中的生物累积和毒性效应[J]. 生态毒理学报,2014, 9(5): 810-918
Wu Y, Wang W X. Bioaccumulation and toxicity of mercury in marine phytoplankton [J]. Asian Journal of Ecotoxicology, 2014, 9(5): 810-918 (in Chinese)
猜你喜欢
福建农业学报(2021年9期)2022-01-05
石材(2020年9期)2021-01-07
石油地质与工程(2019年3期)2019-09-10
中国钼业(2019年2期)2019-01-19
中国盐业(2018年20期)2019-01-14
海洋开发与管理(2017年9期)2017-12-05
采矿技术(2016年2期)2016-12-14
山东青年(2016年2期)2016-02-28
分析化学(2014年10期)2014-10-24
中国环境科学(2014年2期)2014-04-26
