
外源钙(Ca)对毛葱耐镉(Cd)胁迫能力的影响
2014-08-11 14:41王巧玲邹金华刘东华岳洁瑜
生态学报 2014年5期
王巧玲,邹金华,刘东华,岳洁瑜
(天津市动植物抗性重点实验室,天津师范大学,天津 300387)
外源钙(Ca)对毛葱耐镉(Cd)胁迫能力的影响
王巧玲,邹金华,刘东华,岳洁瑜*
(天津市动植物抗性重点实验室,天津师范大学,天津 300387)
以对环境敏感的毛葱(Alliumcepavar.agrogarumL.)为材料,通过水培试验分别研究不同浓度Ca(0、0.1、1、10 mmol/L)对 Cd(10 μmol/L、100 μmol/L和300 μmol/L)胁迫下毛葱幼苗生长、叶片光合特性、体内 Cd 积累和矿质营养的变化,探讨Ca 缓解敏感植物Cd 毒害的生理生化机制。结果表明:(1) Cd显著抑制了毛葱的生长并导致其根端弯曲、发黄,叶片绿色加深;外源Ca显著削弱了Cd的毒害,缓解了其对毛葱生长的抑制;(2)Cd导致毛葱叶片光合色素含量大幅度上升却显著降低了毛葱的光合作用;外源Ca延缓了光合色素上升的速度,提高了毛葱的净光合速率、气孔导度和蒸腾速率;(3)Cd胁迫导致毛葱体内Cd 含量显著增加,并造成明显的矿质营养失衡,主要表现为显著降低了毛葱根中Mg、Mn,叶中Ca、Mg、Mn、Zn等元素的含量,毛葱根中Ca、Fe、Zn,叶中Fe元素含量的显著增加,扰乱了毛葱体内矿质营养的内稳态;外源Ca削弱了毛葱对 Cd 的积累,减轻了Cd 胁迫所造成的矿质营养失衡。因此,外源Ca能通过抑制 Cd的吸收,促进叶片光合作用及气体交换速率,维持植物体的含水量、植物叶片光合色素含量及矿质营养的平衡等途径来增强毛葱对Cd 胁迫的耐性。
毛葱;Cd;Ca;胁迫;耐性
镉(Cd)是一种毒性很大的重金属元素,对植物的生长发育造成伤害,如抑制植物对水分的吸收和运输,抑制光合作用和呼吸作用;对能量转化和蛋白合成造成紊乱;导致营养失衡和超微结构损伤等[1- 5]。植物亦通过不同的生理、生化机制对Cd胁迫作出反应。鉴于此,国内外对如何解除或缓解Cd胁迫的重视程度日益加深。钙(Ca)是植物生长发育所必需的营养元素, 而且能够作为第二信使与 CaM(Calmodulin)结合偶联胞外信号与胞内生理生化反应,通过稳定细胞壁、细胞膜结构[6]及诱导特异基因的表达来提高植物对逆境的抗性[7]。已有研究表明,外源钙可增强植物对许多非生物逆境的适应性,提高植物对重金属毒害的耐性,减轻重金属对植物所造成的伤害[8- 9]。
洋葱(Alliumcepa)不仅是非常重要的蔬菜作物,而且对环境变化敏感,是环境监测的优选植物[10]。毛葱(Alliumcepavar.agrogarumL.)作为洋葱的一个变种(百合科葱属),除了具有洋葱易培养,根系粗壮发达,对环境因子敏感,染色体数目较少(2n=16),形态特征明显等优点外,毛葱还具有体积小,休眠期短的独特优点[11]。目前,有关钙调节Cd对毛葱胁迫方面的研究鲜有报道。本研究以毛葱为试验材料,探讨外源Ca 对Cd胁迫下毛葱幼苗生理生化特性的影响,为缓解重金属Cd对生物的毒害找到经济、有效的方法提供基础数据和理论支持。
1 材料与方法
1.1 植物材料的培养与处理
试验材料为毛葱(Alliumcepavar.agrogarumL.)。选取100头大小均匀一致、饱满、色泽匀称、表皮光滑、健康无损伤、未萌发根也未长绿叶的毛葱鳞茎为实验材料。剥去外部的干燥膜质鳞叶,洗净,在室温下(25 ℃)用自来水培养,每隔12 h换1次水。经自来水培养至刚长出叶片后,从中选取80头生长一致的毛葱,平均分成16组进行处理。在预试验中,筛选出10、100 μmol/L和300 μmol/L CdCl2为适合的Cd胁迫浓度,其中10 μmol/L CdCl2对毛葱幼苗的生长无明显的胁迫作用,100 μmol/L CdCl2既对毛葱幼苗产生胁迫作用,又不会造成严重的Cd毒害,300 μmol/L CdCl2可对毛葱幼苗产生严重的毒害。在3个Cd处理组中,分别加入0、0.1(低钙)、1(中钙)、10 mmol/L(高钙)的Ca,以不施Cd和Ca为对照,具体处理分组见表1。对照及实验组材料均在50% Hoagland营养液中培养,实验组中Cd和Ca是直接溶解于50% Hoagland营养液中,实验周期为12 d,整个实验过程中对培养液通气,每4 d更换1次培养液。
1.2 实验方法
1.2.1 形态观察及株高、根长测定
实验期内每天记录毛葱幼苗生长状况,第12天时测定毛葱幼苗的株高、根长,每处理组测量5个植株,每个植株测量5组数据。数码相机拍照。

表1 实验组编号
1.2.2 光合色素含量测定
样品称重后,于研钵中加入80%丙酮10 mL,将其充分研磨,移入锥形瓶后用封口膜封口,4℃过夜萃取至材料残渣变白,4000×g离心20 min,取上清液,即为色素提取液[12]。测定提取液在波长663,646,470 nm处的光吸收值,按 Lichtenthaler[13]公式计算。
1.2.3 光合速率及气体交换数据的测定
处理12 d时,对16组毛葱进行光合作用及气体交换指数的测定,净光合速率(Pn, μmol CO2·m-2·s-1)、气孔导度(Gs, mmol H2O ·m-2·s-1)及蒸腾速率(Tr, mmol H2O·m-2·s-1)等用LI- 340(CID Co.,USA)手持式光合仪进行测定。测定每株毛葱长势最好的功能叶,每组测定5株,每株测量5个叶片。测量要迅速,测定时间控制在10:00—12:00。
1.2.4 电感耦合等离子发射光谱(ICP-AES)制样及分析
材料处理12 d后收获。分别收集每个实验组毛葱的根系和叶片,共32个样品。每个样品分成相同质量的5组,称重后,将样品分别置于烘箱中烘干:45 ℃,4 d;80 ℃,2 d;105 ℃,过夜。烘干后的样品称重后研磨至粉末,移入干燥瓶中保存,用于消化分析。称0.2 g粉末于小烧杯中,加入5 mL浓硝酸(Analytical Reagent, AR)浸泡过夜;加入2 mL高氯酸(AR)后80 ℃加热3 h,温度升至120 ℃,将剩余硝酸挥发殆尽。冷却后再加入3 mL浓硝酸、2mL高氯酸(AR),缓慢加热,待烧杯中溶液呈橙黄色时,再加入2 mL高氯酸(AR),温度升至200 ℃,至烧杯中有白色晶体析出。样品冷却后,用10%的硝酸溶液溶解并准确定容至25 mL,用ICP-AES(LEEMAN LABS INC., New Hampshire, USA)测定Cd在毛葱各器官中的积累量,并测定根系、叶片中Ca、Mg、Mn、Fe、Zn的含量。每个样品重复测定3次。
1.3 数据统计方法
采用Sigmaplot 8.0软件处理除光合特性以外所有数据并作图,用excel 2003软件处理光合特性数据,并用SPSS 13.0软件进行统计学分析。
2 结果与分析
2.1 外源Ca对Cd胁迫下毛葱生长的影响
2.1.1 对毛葱形态特征的影响
不同浓度外源Ca对Cd胁迫下的毛葱处理12 d后,可明显缓解Cd胁迫造成的毒害(图1)。Cd对毛葱造成毒害,使其形态发生明显的变化,浓度越高其毒害作用越明显。与对照相比,Cd处理组的毛葱生长缓慢,植株矮小,叶片绿色加深,根长受到显著抑制,颜色暗黄,变粗变脆(图1 b, c, d)。在高浓度Cd(100 μmol/L和300 μmol/L)处理组中,与对照组叶片的黄绿色不同,其叶片呈暗绿色夹杂白色小枯斑,根端极度弯曲,发黄呈腐烂状(图1 c, d)。
外源Ca对Cd胁迫下毛葱形态的影响不同(图1):A组和B组毛葱并未因Ca的加入表现出明显的形态差异(图1 a, b);但在C组和D组中,外源Ca对毛葱的毒害有显著的缓解作用,且随外源Ca浓度的增加,其缓解效果也越明显(图1 c, d)。
2.1.2 对毛葱株高、根长的影响
Cd显著抑制毛葱的株高和根长(图2)。外源Ca对A、B组毛葱的株高和根长无明显的影响,但对C、D组毛葱表现出明显的缓解作用(图2)。A、B组中,施加外源Ca后毛葱的株高和根长均无显著性差异(P> 0.05)(图2)。C、D组中,对毛葱株高产生明显影响的外源Ca浓度与对根长产生明显影响的外源Ca浓度不同:10 mmol/L的外源Ca缓解Cd对毛葱株高的毒害作用,C4、D4组毛葱的株高分别比C1、D1组毛葱增高了26.5%和61.5%(图2);C组中,1 mmol/L的外源Ca显著缓解了Cd对毛葱根长的抑制作用,C3组毛葱的根长比C1组增加12.1%(图2)。

图1 外源Ca对Cd胁迫下毛葱生长及形态特征的影响(12 d)Fig.1 Effects of exogenous Ca on morphological characteristics of Allium cepa var. agrogarum L. under Cd stress over 12 days

图2 外源Ca对Cd胁迫下毛葱株高和根长的影响Fig.2 Effects of exogenous Ca on plant height and root length of Allium cepa var. agrogarum L. under Cd stressA:A实验组(1,2,3,4:A1,A2,A3,A4); B:B实验组(1,2,3,4;B1,B2,B3,B4); C:C实验组(1,2,3,4:C1,C2,C3,C4); D:D实验组(1,2,3,4:D1,D2,D3,D4); 同一实验组中字母完全不同表示差异显著(n=15,P<0.05)
2.2 外源Ca对 Cd 胁迫下毛葱叶片光合色素含量的影响
图3表明,Cd胁迫毛葱12 d后导致其叶绿素a含量的显著增加,处理组与对照组之间差异显著(P<0.05)。不同浓度外源Ca对叶绿素a的增加幅度有明显的影响。外源Ca浓度为0.1 mmol/L时,对叶绿素a含量的增加没有明显影响;Ca浓度至1 mmol/L时,能有效缓解除10、100μmol/L处理组毛葱叶片叶绿素a含量的骤升;外施Ca浓度升至10 mmol/L时,对100、300 μmol/L Cd处理组毛葱的缓解作用反而减弱,只对10 μmol/L Cd处理毛葱叶片叶绿素a含量的骤升有明显的抑制作用,使其恢复到正常水平。
Cd对毛葱叶片叶绿素b含量的影响与对叶绿素a含量的影响明显不同(图3)。300 μmol/L Cd处理导致毛葱叶片的叶绿素b含量显著上升(P<0.05),其它浓度的Cd处理组与对照组之间无显著性差异(P> 0.05)。10 mmol/L外源Ca显著抑制Cd胁迫毛葱叶片叶绿素b含量的上升,使其含量与对照组无显著性差异(P> 0.05)(图3)。
类胡萝卜素存在于所有光合生物中,是光合作用的辅助色素,能将吸收的光能传递给叶绿素分子,在捕光色素复合体的组装中承担重要的结构性作用。高浓度的Cd(100 μmol/L和300 μmol/L)能显著增加类胡萝卜素的含量(P<0.05)(图3)。10 mmol/L外源Ca可缓解100 μmol/L Cd毒害,低于此浓度的Ca均未对类胡萝卜素的含量产生影响(图3)。
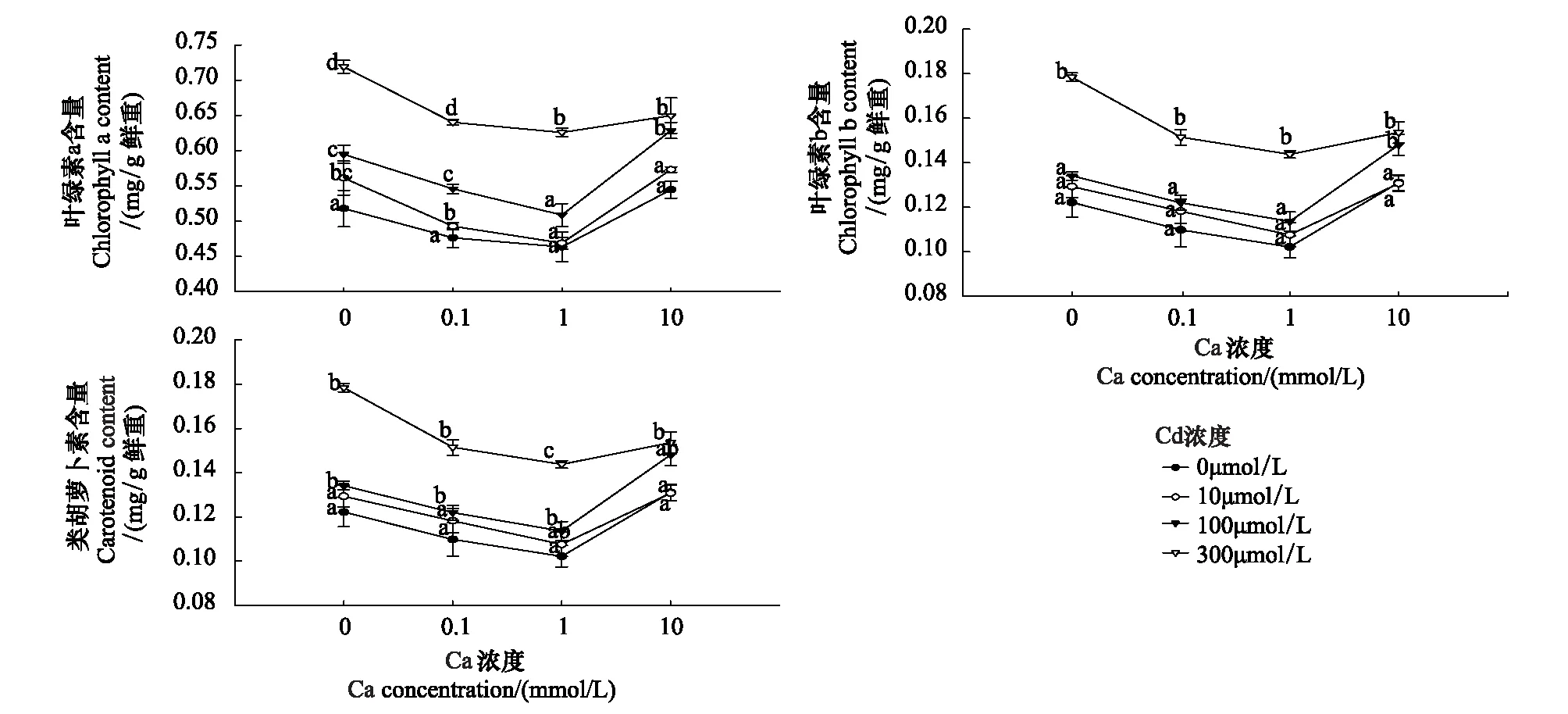
图3 外源Ca对Cd胁迫下毛葱光合色素含量的影响Fig.3 Effects of exogenous Ca on leaf pigment content of Allium cepa var. agrogarum L. under Cd stress同列字母完全不同表示差异显著(n=15,P<0.05)
2.3 外源Ca对Cd胁迫下毛葱叶片光合特性的影响
2.3.1 外源Ca对Cd胁迫下毛葱叶片净光合速率的影响
净光合速率(Pn)是反映光合作用强弱的最重要指标,净光合速率越大,植物光合作用的水平越高,反之亦然。Cd胁迫下毛葱叶片净光合速率受到明显的影响,随Cd处理浓度的增加而明显降低(图4)。Cd胁迫毛葱12 d后,10、100 μmol/L和300 μmol/L Cd处理组毛葱其叶片净光合速率比对照组分别下降了24.2%、75.8% 和85.4%。外源Ca对其下降幅度有明显的影响。具体表现为,0.1 mmol/L Ca对Cd的毒害无明显缓解作用;1 mmol/L Ca显著缓解300 μmol/L Cd的毒害(P<0.05);10 mmol/L外源Ca比1 mmol/L Ca缓解效果更明显,对100 μmol/L和300 μmol/L Cd毒害均有明显缓解作用(图4)。
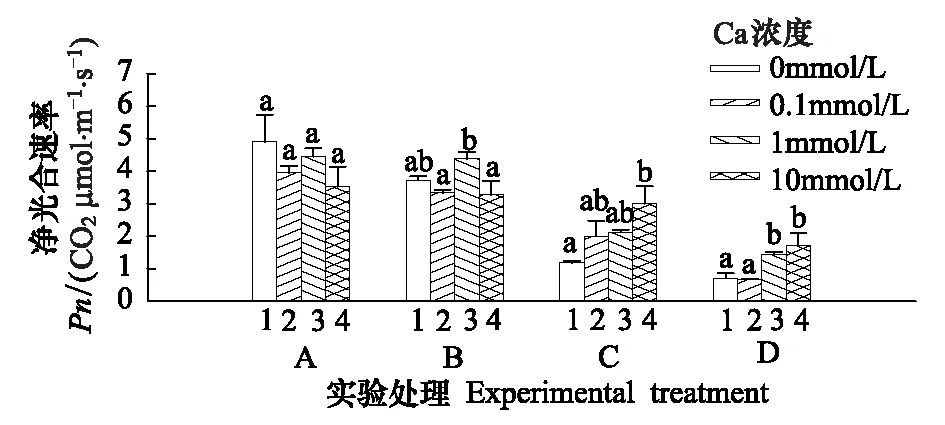
图4 不同浓度Ca对Cd胁迫下毛葱处理12 d后净光合速率(Pn)的影响Fig.4 Effects of different concentrations of Ca on the net photosynthetic rate (Pn) in Allium cepa var. agrogarum L. exposed to Cd stress over 12 d同一实验组中字母完全不同表示差异显著(n=15,P<0.05)
2.3.2 外源Ca对Cd胁迫下毛葱叶片气孔导度的影响
气孔导度(Gs,单位为mmol H2O·m-2·s-1),影响植物的光合作用和蒸腾作用。不同浓度的Cd对毛葱叶片的气孔导度有不同程度的抑制作用(图5)。10、100 μmol/L和300 μmol/L Cd处理毛葱12 d后,其叶片的气孔导度比对照组分别下降了19.8%、42.7% 和81.3%。施加外源Ca显著降低了A组叶片的气孔导度(P<0.05),但对B、C、D组叶片的气孔导度没有明显的影响(图5)。

图5 不同浓度Ca对Cd胁迫下毛葱处理12 d后气孔导度(Gs)的影响Fig.5 Effects of different concentrations of Ca on the stomatal conductance (Gs) in Allium cepa var. agrogarum L. exposed to Cd stress over 12 days同一实验组中字母完全不同表示差异显著(n=15,P<0.05)
2.3.3 外源Ca对Cd胁迫下毛葱叶片蒸腾速率的影响
蒸腾作用是植物吸收水分和转运水分的主要动力,对维持植物各部分的水分饱和,保持细胞组织的形态,促进无机盐类在植物体内的分布具有重要的作用。对比图6和图5可以看出,蒸腾速率(Tr)和气孔导度的变化趋势一致。不同浓度Cd胁迫毛葱12 d后,10、100 μmol/L和300 μmol/L Cd处理后毛葱叶片的蒸腾速率分别比对照下降了14.2%、43.1%和78.0%。施加外源Ca显著降低了A组叶片的蒸腾速率(P<0.05);外源Ca显著缓解了100 μmol/L Cd对毛葱的毒害(P<0.05),提高了毛葱叶片的蒸腾速率,尤其当Ca浓度大于1 mmol/L时,缓解效果尤为明显;外源Ca对B、C、D组叶片的蒸腾速率没有明显的影响(图6)。

图6 不同浓度Ca对Cd胁迫下毛葱处理12 d后蒸腾速率(Tr)的影响Fig.6 Effects of different concentrations of Ca on the transpiration rate (Tr) in Allium cepa var. agrogarum L. exposed to Cd stress over 12 days同一实验组中字母完全不同表示差异显著(n=15,P<0.05)
2.4 外源Ca对Cd胁迫下毛葱Cd含量及营养元素吸收的影响
2.4.1 外源Ca对Cd胁迫下毛葱含水量的影响
Cd显著降低毛葱体内的含水量,施加外源Ca能明显的减缓其降低幅度(图7)。本研究中毛葱幼苗根系的鲜干重比与Cd 浓度呈负相关,B1、C1、D1组幼苗(10—300 μmol/L)根系的鲜干重比始终显著低于A1组(对照组)(P<0.05)且各处理组间差异显著(P<0.05)。施加外源Ca后各实验组毛葱根系的鲜干重比均先升高后随着Ca浓度的提高(>0.1 mmol/L)而逐渐降低,在Ca浓度为10 mmol/L 时最低(图7)。
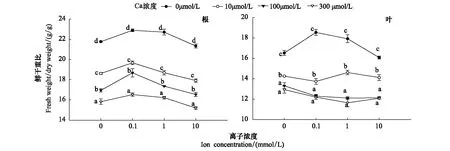
图7 不同浓度Ca对Cd胁迫毛葱12 d后各器官鲜干重比值的影响Fig.7 Effects of different concentrations of Ca on the fresh and dry weights ratio in Allium cepa var. agrogarum L. exposed to Cd stress over 12 days 同列字母完全不同表示差异显著(n=15,P<0.05)
毛葱叶片鲜干重比的变化趋势与根系不完全一致(图7)。毛葱叶片的的鲜干重比与Cd浓度呈现负相关,除C1和D1组外其它各实验组间差异显著(P<0.05)。施加外源Ca后,A组中毛葱叶片鲜干重比的变化趋势与根系保持一致。B、C、D组在Ca浓度为0.1 mmol/L 时比值均下降,与根系的变化趋势相反;当Ca浓度为1 mmol/L 时,B组比值上升,C、D组均下降;当Ca浓度至10 mmol/L时,B、C、D各组叶片的变化趋势正好与1 mmol/L Ca处理组相反(图7)。
2.4.2 外源Ca对Cd 胁迫下毛葱体内 Cd 含量的影响
Cd处理后毛葱体内Cd含量随Cd浓度上升而增加,但Cd在毛葱根系中的积累量远远高于叶片中的含量(图8)。施加外源Ca显著降低了B、C、D组毛葱根系和叶片中Cd的积累量(P<0.05)。
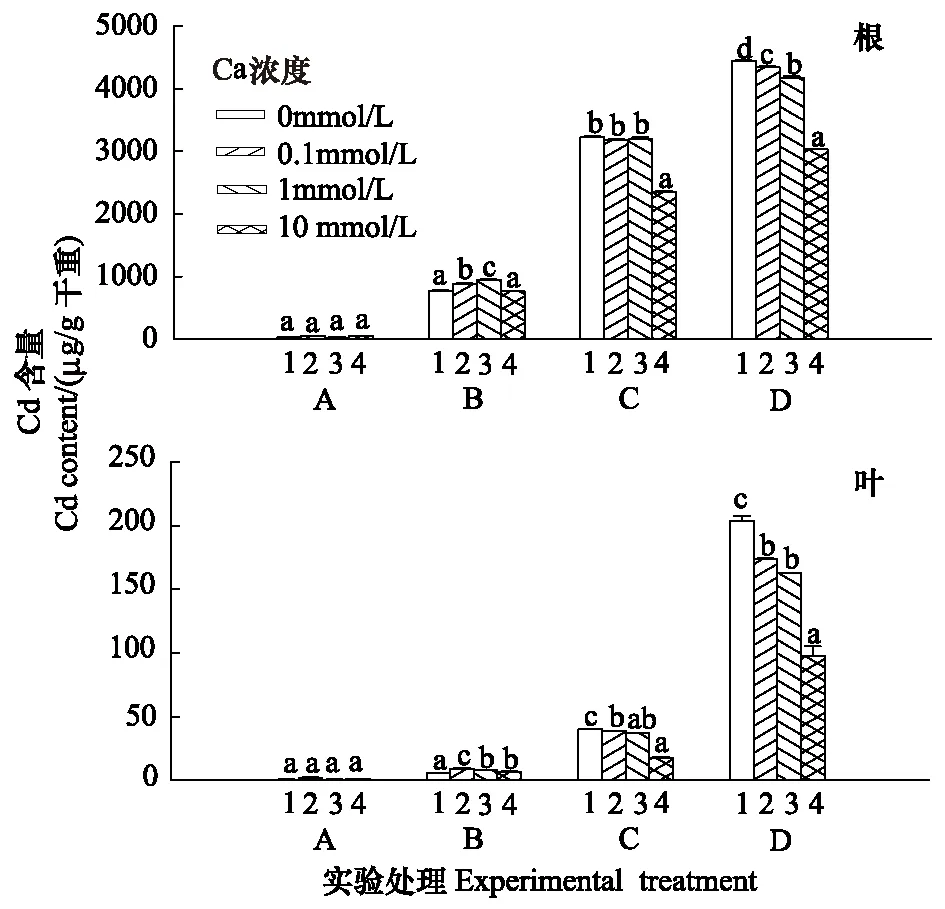
图8 不同浓度Ca对Cd胁迫下毛葱12 d后,Cd在毛葱体内不同器官中的积累和分布Fig.8 Cadmium uptake and accumulation by Allium cepa var. agrogarum L. under cadmium stress, after 12 days cultivation with different concentration of Ca 同一实验组中字母完全不同表示差异显著(n=5,P<0.05)
外源Ca对毛葱根系中Cd的积累量有明显的影响(图8)。B组中,1 mmol/L外源Ca显著促进Cd在毛葱根系中的积累(P<0.05),10 mmol/L Ca对其积累量无明显影响。在C、D组中,10 mmol/L外源Ca能显著降低Cd在根系的积累(P<0.05);D组中,0.1 mmol/L外源Ca能显著降低Cd在毛葱根系中的积累(P<0.05),随着Ca浓度的升高,降低效果更为显著(图8)。
与外源Ca对毛葱根系中Cd的积累的影响一致,毛葱叶片中Cd的积累量随着施加外源Ca而明显降低(图8)。与根不同的是,0.1 mmol/L的Ca就能显著降低C组叶片中Cd的积累量;而在D组中,0.1 mmol/L和1 mmol/L 外源Ca对毛葱叶片中Cd的积累量没有产生明显的影响(P> 0.05)(图8)。
2.4.3 外源Ca对Cd 胁迫下毛葱营养元素吸收的影响
Cd胁迫造成毛葱体内明显的矿质营养失衡,主要表现为显著降低毛葱根中Mg、Mn,叶中Ca、Mg、Mn、Zn等元素的含量;显著增加毛葱根中Ca、Fe、Zn,叶中Fe元素的含量,扰乱毛葱体内矿质营养的内稳态;外源Ca削弱了Cd 胁迫造成的矿质元素失衡,稳定了毛葱体内矿质营养的内稳态(表2)。
Cd造成毛葱根系中钙(Ca)含量显著高于对照组(A1)(P<0.05)并随Cd浓度的升高而增加;不同浓度的外源Ca对Cd胁迫下毛葱体内钙(Ca)的代谢与利用有不同的影响(表2)。A组中,外源Ca浓度低于10 mmol/L时显著降低毛葱根系中Ca的含量(P<0.05);当Ca浓度升至10 mmol/L时,根系中Ca的积累量显著高于对照组(P<0.05)。B、C、D组中,外源Ca显著降低毛葱根系Ca含量。Ca在毛葱叶片中的吸收和积累情况与根系中明显不同(表2)。Cd显著降低毛葱叶片中的Ca含量。A组中,外源Ca浓度为0.1、1和10 mmol/L时,叶片中Ca的含量比对照组分别增加了17.3%、37.8% 和44.7%;B、C、D组中,Ca在毛葱叶片的积累随外源Ca浓度的变化趋势与根系保持一致。A组施加外源Ca后,毛葱体内Ca含量较对照组显著增加(P<0.05);而B、C、D组中毛葱体内Ca含量却随着外源Ca的施加而减少。
Cd显著地抑制了毛葱对镁(Mg)的吸收和积累;不同浓度外源Ca对Cd胁迫下毛葱体内Mg的代谢与利用有不同的影响(表2)。毛葱根系Mg含量显著受到外源Ca的影响。A组中,施加外源Ca后,毛葱根系中Mg含量较对照组显著降低(P<0.05),0.1 mmol/L Ca对其降低效果最明显仅为对照组(A1)的83.3%;B、C、D组中0.1 mmol/L 外源Ca施加组(B2、C2、D2),毛葱根系中Mg含量仅为其无Ca组(B1、C1、D1)含量的71.0%、96.0%和63.9%(表2)。毛葱叶片中Mg含量也明显受到外源Ca的影响(表2)。与根系中Mg含量不同的是,A组中,0.1 mmol/L 外源Ca处理下毛葱叶片中Mg含量高于对照组及其它Ca处理组(P<0.05)。随着Ca浓度的升高,叶片中Mg的积累量逐渐下降,10 mmol/L时达最低且显著低于对照组(P<0.05);B、C、D组与A组变化趋势一致。0.1 mmol/L外源Ca能显著降低毛葱根系中Mg含量并随外源Ca浓度的增加降低幅度减小;叶片中Mg含量在0.1 mmol/L外源Ca组中显著增加,但其增加幅度随外源Ca浓度的升高而减小(表2)。

Cd显著促进了毛葱对铁(Fe)的吸收(P<0.05);不同浓度外源Ca对Cd胁迫下毛葱体内Fe的代谢与利用有不同的影响(表2)。根系中:A、B组Fe含量随外源Ca浓度的升高而增加,其中A4、B4组Fe含量分别为A1、B1组的143.7% 和130.6%;C、D组Fe含量在外源Ca浓度为0.1 mmol/L(C2、D2组)时最大,其值分别为C1组、D1组的107.2% 和119.1%,当外源Ca浓度大于0.1 mmol/L时,毛葱根系中Fe含量逐渐减少(表2)。叶片中:A、C和D实验组均在外源Ca浓度为0.1 mmol/L(A2、C2、D2组)时叶片中Fe含量最大,其值分别比A1、C1和D1组增加了111.3%、61.5%和44.0%,外源Ca浓度大于0.1 mmol/L时,毛葱叶片中Fe含量逐渐减少;B组中,毛葱叶片中Fe含量在Ca浓度为1 mmol/L最大,为B1组的124.7%。因此,施加外源Ca能显著增加毛葱体内Fe的积累量,0.1 mmol/L Ca效果最明显。
Cd显著抑制了毛葱对锰(Mn)的积累(P<0.05);不同浓度外源Ca对Cd胁迫下毛葱体内Mn的代谢与利用有不同的影响(表2)。A实验组中,外源Ca会显著降低毛葱根系中Mn的积累量(P<0.05),Ca浓度为0.1 mmol/L时含量达最低,为对照组的68.7%;B、C、D实验组中,外源Ca也会显著降低毛葱根系中Mn的积累量(P<0.05),但只有当Ca浓度升至10 mmol/L时才会使毛葱根中Mn的积累量达到各组的最低含量(表2)。与根系不同的是,外源Ca会显著增加A组中毛葱叶片中Mn的含量(P<0.05),在Ca浓度为0.1 mmol/L时叶片中Mn含量达最高为A1组的157.5%;B、C、D实验组中,毛葱叶片中Mn含量随外源Ca浓度的施加而降低(表2)。因此,外源Ca能显著降低Cd胁迫下毛葱体内Mn的积累量。
Cd显著促进了毛葱根系对锌(Zn)的积累却显著抑制了叶片对Zn的积累(P<0.05);不同浓度外源Ca对Cd胁迫下毛葱体内Zn的代谢与利用的影响不同(表2)。A组中,外源Ca能显著促进毛葱根系对Zn的吸收(P<0.05),0.1 mmol/L Ca处理组比对照组增加了29.8%;B组中,外源Ca显著增加毛葱根系中Zn的含量(P<0.05),当Ca浓度为1 mmol/L时达到Zn积累量的最大值,为B1组的122.1%;C组中10 mmol/L Ca对根中Zn的含量产生显著抑制作用(P<0.05),低于此浓度的Ca对其均无明显影响;D组中,外源Ca能显著抑制毛葱根中Mn含量(P<0.05),Ca浓度为1 mmol/L时含量最低,比D1组下降36.8%(表2)。毛葱叶片中Zn的积累与根系的变化趋势不同(表2)。A组中,当外源Ca浓度为0.1 mmol/L时可显著促进Zn在毛葱叶片的积累(P<0.05),而10 mmol/L的Ca显著抑制Zn在毛葱叶片的含量(P<0.05),1 mmol/L Ca对其无明显影响(P>0.05);B实验组,0.1mmol/L Ca显著地促进Zn在毛葱叶片中的积累(P<0.05),但当Ca浓度上升时,其对Zn在毛葱叶片的积累量与B1组无显著差异(P> 0.05);C实验组中外源Ca未对Zn在毛葱叶片的积累产生明显影响;D实验组中,外源Ca能显著增加毛葱根中Mn的含量(P<0.05)。
3 讨论
3.1 外源Ca对 Cd 胁迫下毛葱生长的影响
株高根长是水培模拟试验中最易观测的植株外观指标,并能反映植物所受胁迫的强度[14]。Cd胁迫严重影响了毛葱的形态特征,使其株高、根长也受到明显的抑制,外源Ca能有效缓解Cd对毛葱幼苗产生的毒害,使其形态得到有效恢复、对其生长的抑制也明显减弱(图1,图2)。因为Ca作为植物必需的营养元素之一,参与植物的多种生命活动与多种酶的代谢过程[15],可在一定程度上缓解Cd的毒害,促进植物体的生长。
植株的生理代谢活性能够反映环境对植物的适宜程度,含水量的高低是衡量植物生理活性的一个重要指标。采用植株鲜重(FW)与干重(DW)的比值代表植株的含水量。Cd造成毛葱体内含水量的显著下降,造成体内水分失衡(图7)。植物对这一失衡关系的调节主要是通过相应有机物的合成和无机离子的积累两条途径来实现[16- 19]。外源Ca促进了毛葱根系的生长,保持了毛葱体内水分的相对平衡(图7),可能是由于外源Ca对根尖部位离子关系有明显的改善,从而有利于根系生长[18,20](图1)。Ca作为植物必需的营养元素之一,是多种酶的组成成分和激活剂,能调节细胞的充水度及渗透性,维持细胞正常的生理状态,从而维持植物体的正常生理代谢,因此,Ca对于降低植物对Cd的吸收,及减轻Cd对植物的毒害具有重要作用[21]。
3.2 外源Ca对Cd 胁迫下毛葱生理特性的影响
Cd 毒害造成植物的生理代谢紊乱,加速植物的衰老。有研究表明,叶片衰老过程中伴随着叶绿素酶活性提高,叶绿素降解加剧。唐蕾等[22]采用反相高效液相色谱,结合光电二极管阵列和荧光检测技术,发现在衰老的银杏叶中主要存在叶绿素a、叶绿素b 和脱镁叶绿素(暗橄榄褐色),脱植基叶绿素(鲜绿色)。在液泡膜上也发现了一种依赖ATP(Adenosine Triphosphate, ATP)的特异运载蛋白,负责叶绿素代谢物在液泡中转运[23]。由上可知,叶绿素的许多卟啉衍生物在叶片衰老过程中从衰老叶绿体转运到液泡中贮存起来,这些叶绿素代谢物如脱植基叶绿素等的光谱性质和叶绿素原体基本相同,可在同一波长下被检测到,但这些叶绿素的代谢产物均不能行使正常叶绿素的功能,从而降低了叶片的光合特性。毛葱受Cd毒害后其色素含量上升但其光合作用却严重受到了抑制。叶绿素降解除了影响光合作用外还具有其它的意义。例如,衰老时叶绿素降解启动蛋白质的水解,利于氮的释放利用;一些叶绿素降解产物或衍生物可以提高细胞对重金属毒害的耐受力[24]。外源Ca对毛葱幼苗色素含量产生较大影响。毛葱叶片色素含量受外源Ca水平的限制,当外源Ca浓度为0.1 mmol/L时毛葱叶片叶绿素和类胡萝卜素含量与Ca对照组相比均显著下降,Ca浓度升至1 mmol/L时,色素含量继续下降,但当Ca浓度为10 mmol/L时,其含量反而上升(低于无施加Ca组)。这可能与外源Ca促进了毛葱的快速生长,其生长量增大导致的“稀释效应”有关[25],但高浓度外源Ca(10 mmol/L)对毛葱幼苗来说可能是一种营养胁迫,在这种营养胁迫下,Ca可能会造成强光胁迫,这与邱栋梁[26]的研究结果是一致的,即Ca能减少叶绿素a、b和类胡萝卜素含量的下降。类胡萝卜素是叶绿体内的天线色素,与光合作用中的光系统Ⅰ(PSⅠ)和光系统Ⅱ(PSⅡ)相连结,一方面阻止激发态叶绿素分子的激发能从反应中心向外传递,另一方面又能保护叶绿素分子的激发能免遭光氧化损伤,是单线态氧重要的猝灭剂[27],能够在一定程度上保护光合机构,调节过剩光能的耗散,减轻过剩光能对光合机构的破坏。 因此,毛葱叶片在10 mmol/L外源Ca处理下保持了较高的类胡萝卜素含量[25]。这也是其耐Cd的一种有效措施。
Cd能使毛葱叶片的净光合速率(Pn)、气孔导度(Gs)和蒸腾速率(Tr)持续降低,外源Ca明显减小了其降低幅度,使Pn维持在较高的水平(图4)。这可能是由于Cd造成毛葱体内矿质营养失衡所致。Mn和Ca都是光合放氧复合物中的重要组成成分。Mn是植物必需营养元素,与其体内许多酶的活性和氧化还原体系有关,并直接参与光合作用[28]。Mn含量的下降而使叶绿体的放氧活性下降,从而抑制了毛葱的光合效率[26]。而外源Ca能适当提高毛葱的净光合速率主要是因为Ca能提高叶绿体二氯酚吲哚酚(Dichlorophenol Indophenol, DCPIP)光还原活性。在PSⅡ膜中,每个PSⅡ含有一个高亲合性及两个低亲合性的Ca的结合位点;这些位点上含有2—3个结合态Ca,并且非常靠近Mn复合物[29],Ca能提高叶绿体DCPIP光还原活性可能是因为Ca促进Mn簇电子向原初氧化剂传递。已有研究证明Ca的作用位于Mn的S1—S4(不同氧化还原状态的放氧复合体)状态之后,接近原初氧化剂Y+,且促进Y+的还原[30]。Handreck[31]认为,Ca供应过多会引起植物出现缺Mg的症状,这与本文的研究结果是一致的(表2)。肖细元等[27]也指出Ca处理显著限制Mg的吸收,高钙处理使蜈蚣草羽片中镁浓度含量极低,几乎为对照处理的一半。Mg和Mn一样在植物的光合作用中发挥着重要的作用,Mg是叶绿素的重要组成成分,同时Mg也是催化磷酸化过程酶的关键辅助因子之一,在磷酸盐的代谢过程中起着重要的作用[32]。因此,Mg的缺乏也可造成植物净光合速率的显著下降。
在Cd胁迫下毛葱体内Zn、Fe和Ca等半径与Cd相似的离子含量均有所上升,因为Fe是植物体某些酶的组成成分,在氧化还原过程中起着重要的电子传递作用,而Zn与植物蒸腾作用有关,Zn还参与植物生长素的形成,促进光合作用和蛋白质合成[27]。这些离子与Cd竞争毛葱细胞表面有限的离子通道,进而减少毛葱对Cd的吸收,从而减轻Cd毒害,另一方面,Fe、Zn含量的增加进一步保证了毛葱的正常生理,从而有利于毛葱光合作用的提高。另外有研究报道,Zn含量的上升还可能是因为Zn与Ca之间存在共积累关系[33- 34]。Cd 胁迫所造成的矿质营养失衡,影响植物正常的生理代谢[35- 36],导致植物生物量下降[37]。外源Ca能缓解Cd的毒害,因为Ca与Cd具有相似的化学性质,是Cd吸附位点的主要竞争者,当溶液中Ca和Cd同时存在时,Ca可显著地降低植物对Cd的吸收和积累,从而减轻Cd的毒害。植物所处环境中的离子的物理性质(化学价、离子半径)的相似性会影响植物对离子的吸收及分配,如Cd能够利用Ca2+通道被植物运输,运Fe载体也参与Cd的运输[38- 40]。另有研究指出,在Cd胁迫下,外源Ca使细胞内游离Ca常常显著增加,以便于启动特定基因的表达,激活一系列生化反应,从而使植物适应环境中的胁迫因子[39- 48]。
4 结论
(1)不同浓度外源Ca 对Cd胁迫下的毛葱处理12 d后,对Cd胁迫所造成的伤害具有不同程度的缓解作用。其中,10 mmol/L的外源Ca对毛葱生长的缓解作用最为显著。
(2)外源Ca对 Cd 胁迫下毛葱叶片光合色素含量的影响。Cd胁迫毛葱12 d后能导致其叶绿素a含量的显著增加,处理组与对照组之间差异显著(P<0.05)。不同浓度外源Ca 对叶绿素a的增加幅度有明显的影响。1 mmol/L Ca有效的缓解了300 μmol/L Cd处理组毛葱叶片叶绿素a含量的骤升;10 mmol/L Ca使10 μmol/L Cd处理毛葱叶片叶绿素a含量恢复到正常水平。
(3)外源Ca 对Cd胁迫下毛葱叶片光合特性的影响。Cd胁迫毛葱12 d后,10、100 μmol/L和300 μmol/L Cd处理毛葱其叶片净光和速率比对照组分别下降了24.2%、75.8% 和85.4%。外源Ca 对其下降幅度有明显的影响。具体表现为,Ca 浓度为0.1 mmol/L时对Cd的毒害无明显缓解作用;当Ca 浓度达1 mmol/L时可显著缓解300 μmol/L Cd的毒害(P< 0.05);当外源Ca浓度高达10 mmol/L时,比1 mmol/L有更好的缓解作用,对100 μmol/L和300 μmol/L Cd毒害均有明显缓解作用。
(4)Cd处理后毛葱体内Cd含量显著增加且与Cd浓度呈正相关性,但Cd在毛葱根系中的积累量远远高于叶片中的含量。施加外源Ca显著降低了Cd处理组中毛葱根系和叶片中Cd的积累量(P<0.05)。与外源Ca 对毛葱根系中Cd的积累的影响一致,毛葱叶片中Cd的积累量随着施加外源Ca 而明显降低。
(5)Cd胁迫造成毛葱体内明显的矿质营养失衡,显著降低毛葱根中Mg、Mn,叶中Ca、Mg、Mn、Zn等元素的含量;却显著增加了毛葱根中Ca、Fe、Zn,叶中Fe元素的含量,扰乱毛葱体内矿质营养的内稳态;外源Ca削弱了Cd 胁迫所造成的矿质元素失衡,稳定了毛葱体内矿质营养的内稳态。
[1] Das P, Samantaray S, Rout G R. Studies on cadmium toxicity in plants: a review. Environmental Pollution, 1997, 98(1): 29- 36.
[2] Xu Q S, Shi G X, Zhou Y M, Wu G R, Wang X. Distribution and toxicity of cadmium inHydrillaverticillata(L. F.) Royle. Acta Biologiae Experimentalis Sinica, 2004, 37(6): 461- 468.
[3] Xu Q S, Shi G X, Wang X, Wu G R. Generation of active oxygen and change of antioxidant enzyme activity inHydrillaverticillataunder Cd, Cu and Zn stress. Acta Hydrobiologica Sinica, 2006, 30(1): 107- 112.
[4] Li S, Yu J L, Zhu M J, Zhao F G, Luan S. Cadmium impairs ion homeostasis by altering K+and Ca2+channel activities in rice root hair cells. Plant, Cell & Environment, 2012, 35(11): 1998- 2013.
[5] Xu Q S, Ji W D, Yang H Y, Wang H X, Xu Y, Zhao J, Shi G X. Cadmium accumulation and phytotoxicity in an aquatic fern,Salvinianatans(Linn.). Acta Ecologica Sinica, 2009, 29(6): 3019- 3027.
[6] Cheval C, Aldon D, Galaud J P, Ranty B. Calcium/calmodulin-mediated regulation of plant immunity. Biochimica et Biophysica Acta (BBA)-Molecular Cell Research, 2013, 1833(7): 1766- 1771.
[7] Gong M, Li Y J, Chen S Z. Abscisic acid-induced thermotolerance inMaizeseedlings is mediated by calcium and associated with antioxidant systems. Journal of Plant Physiology, 1998, 153(3/4): 488- 496.
[8] Wang H, Zhou W, Lin B. Effects of Ca on growth and some physiological characteristics ofmaizeunder Cd stress. Plant Nutrition and Fertilizer Science, 2001, 7(1): 78- 87.
[9] Chen X L, Yu T Y, Qin H M, Liang X L, Wu P D, Lun J H, Zhou L Y. The effect of calcium on corn seedling growth and physiological characteristics under chromium stress. Journal of Maize Sciences, 2009, 17(4): 74- 78.
[10] Fiskesjö G. Nucleolar dissolution induced by aluminium in root cells ofAllium. Physiologia Plantarum, 1983, 59(3): 508- 511.
[11] Qin R, Jiao Y Q, Zhang S S, Jiang W S, Liu D H. Effects of aluminum on nucleoli in root tip cells and selected physiological and biochemical characters inAlliumcepavar.agrogarumL. BMC Plant Biology, 2010, 10: 225, doi: 10.1186/1471- 2229- 10- 225.
[12] Zhang Z L, Zhai W J. Guide of Plant Physiology Experiment. 3rd ed. Beijing: Higher Education Press, 2005.
[13] Lichtenthaler H K. Chlorophylls and carotenoids: pigments of photosynthetic biomembranes. Methods in Enzymology, 1987, 148: 350- 382.
[14] Zhang F, Luo C D, Zhang J. Alleviation effect of exogenous Ca, P and N on the growth ofChinesefirseedlings under Al stress. Chinese Journal of Applied Ecology, 2005, 16(2): 213- 217.
[15] Marschner H. Mineral Nutrition of Higher Plants. 2nd ed. New York: Academic Press, 1995.
[16] McNulty I B. Rapid osmotic adjustment by a succulent halophyte to saline shock. Plant Physiology, 1985, 78(1): 100- 103.
[17] Guerrier G. Fluxes of Na+, K+and Cl-and osmotic adjustment inLycopersiconpimpinellifoliumandL.esculentumduring short- and long-term exposures to NaCl. Physiologia Plantarum, 1996, 97(3): 583- 591.
[18] Zhong H, Lauchli A. Spatial distribution of solute, K, Na, Ca and their deposition rates in the growth zone of primary cotton roots: Effects of NaCl and CaCl2. Planta, 1994, 194(1): 34- 41.
[19] Talukdar D. Exogenous calcium alleviates the impact of cadmium-induced oxidative stress inLensculinarismedic. Seedlings through modulation of antioxidant enzyme activities. Journal of Crop Science and Biotechnology, 2012, 15(4): 325- 334.
[20] Colmer T D, Fan T W M, Higashi R M, Läuchli A. Interactive effects of Ca2+and NaCl salinity on the ionic relations and proline accumulation in the primary root tip ofSorghumbicolor. Physiologia Plantarum, 1996, 97(3): 421- 424.
[21] McLaughlin M J, Singh B R. Cadmium in Soils and Plants. Dordrecht: Kluwer Academic Publishers, 1999.
[22] Tang L, Chen Y, Wang W. Reversed phase high performance liquid chromatography analysis of chlorophyll derivatives inGinkgo(GinkgobilobaL.) leaves. Plant Physiology Communications, 2006, 42(2): 265- 267.
[23] Shen C G, Zhang F S, Mao D R. Advances in degradation metabolism of chlorophyll during plant leaf senescence. Chinese Bulletin of Botany, 1998, 15(S1): 41- 46.
[24] Ghosh A K, Sen S, Palit S, Ghosh A, Sharma A, Talukder G. Comparative efficacy of chlorophyllin in reducing cytotoxicity of some heavy metals. Biology of Metals, 1991, 4(3): 158- 161.
[25] Dong C X, Zhou J M, Zhao S J, Wang H Y. Effects of exogenous Ca on some physiological characteristics of tomato (Lycopersiconesculentum) seedlings with different Ca sensitivity. Chinese Journal of Applied Ecology, 2005, 16(2): 267- 272.
[26] Qiu D L, Liu X H, Guo S Z. Regulation function of calcium on photosynthesis ofDimocarpuslonganaLour. cv. Wulongling under simulated acid rain stress. Chinese Journal of Applied Ecology, 2002, 13(9): 1072- 1076.
[27] Wang A G. The oxygen metabolism in plant // Yu S W, Tang Z C, eds. Plant Physiology and Molecular Biology. Beijing: Science Press, 1998: 366- 389.
[28] Xiao X Y, Liao X Y, Chen T B, Zhang Y Z. Effects of arsenic and calcium on metal accumulation and translocation inPterisvittataL.. Acta Ecologica Sinica, 2003, 23(8): 1477- 1487.
[29] Hall D O, Rao K K. Photosynthesis. 5th ed. London: Cambridge University Press, 1994: 76- 165.
[30] Crane F L, Bar R. Function of calcium in chloroplasts // Trewavas A J, ed. Molecular and Cellular Aspects of Calcium in Plant Development. New York: Plenum Press, 1986: 269- 276.
[31] Handreck K A. Growth of ferns in soil less media, as affected by pH, iron and calcium/magnesium ratio. Scientia Horticulturae, 1992, 50(1/2): 115- 126.
[32] Mengel K, Kirkby E A. Principles of Plant Nutrition. Worblaufen-Bern: International Potash Institute, 1978.
[33] Skórzyńska-Polit E, Tukendorf A, Selstam E, Baszyński T. Calcium modifies Cd effecton runnerbean plants. Environmental and Experimental Botany, 1998, 40(3): 275- 286.
[34] Sarret G, Harada E, Choi Y E, Isaure M P, Geoffroy N, Fakra S, Marcus M A, Birschwilks M, Clemens S, Manceau A. Trichomes of tobacco excrete zinc as zinc-substituted calcium carbonate and other zinc-containing compounds. Plant Physiology, 2006, 141(3): 1021- 1034.
[35] Grattan S R, Grieve C M. Mineral element acquisition and growth response of plants grown in saline environments. Agriculture, Ecosystems & Environment, 1992, 38(4): 275- 300.
[36] Sagi M, Dovrat A, Kipnis T, Lips H. Ionic balance, biomass production, and organic nitrogen as affected by salinity and nitrogen source in annual ryegrass. Journal of Plant Nutrition, 1997, 20(10): 1291- 1316.
[37] Ruiz D, Martinez V, Cerda A. Citrus response to salinity: Growth and nutrient uptake. Tree Physiology, 1997, 17(3): 141- 150.
[38] Cohen C K, Fox T C, Garvin D F, Kochian L V. The role of iron-deficiency stress responses in stimulating heavy-metal transport in plants. Plant Physiology, 1998, 116(3): 1063- 1072.
[39] Gong M, Chen S N, Song Y Q, Li Z G. Effect of calcium and calmodulin on intrinsic heat tolerance in relation to antioxidant systems in maize seedlings. Australian Journal of Plant Physiology, 1997, 24(3): 371- 379.
[40] Gong M, van der Luit A H, Kinght M R, Trewavas A J. Heat shock induced changes in intracellular Ca2+level in tobacco seedlings in relation to thermo tolerance. Plant Physiology, 1998, 116(1): 429- 437.
[41] Hanson J B. The functions of calciurn in plant nutrition. Advanced Plant Nutrition, 1984, 1: 149- 208.
[42] McLaughlin M J, Singh B R. Cadmium in Soils and Plants. Dordrecht: Kluwer Academic Publishers, 1999.
[43] Cheng S H, Willmann M R, Chen H C, Sheen J. Calcium signaling through protein Kinases. The Arabidopsis calcium-dependent protein kinase gene family. Plant Physiology, 2002, 129(2): 469- 485.
[44] McLanughlin S B, Wimmer R. Calcium physiology and terrestrial ecosystem processes. New Phytology, 1999, 142(3): 373- 417.
[45] Sander D, Pelloux J, Brownlee C, Harper J F. Calcium at the crossroads of signaling. Plant Cell, 2002, 14(S1): S401- S417.
[46] Trewavas A. Le calcium, C′est la vie: Calcium makes waves. Plant Physiology, 1999, 120(1): 1- 6.
[47] Kinoshita T, Nishimura M, Shimazaki K. Cytosolic concentration of Ca2+regulates the plasma membrane H+-ATPase in guard cells of fava bean. Plant Cell, 1995, 7(8): 1333- 1342.
[48] Min H L, Cai S J, Xu Q S, Shi G X. Effects of exogenous calcium on resistance ofHydrillaverticillata(L. f.) Royle to cadmium stress. Acta Ecologica Sinica, 2012, 32(1): 256- 264.
参考文献:
[2] 徐勤松, 施国新, 周耀明, 吴国荣, 王学. 镉在黑藻叶细胞中的亚显微定位分布及毒害效应分析. 实验生物学报, 2004, 37(6): 461- 468.
[3] 徐勤松, 施国新, 王学, 吴国荣. 镉、铜和锌胁迫下黑藻活性氧的产生及抗氧化酶活性的变化研究. 水生生物学报, 2006, 30(1): 107- 112.
[5] 徐勤松, 计汪栋, 杨海燕, 王红霞, 许晔, 赵娟, 施国新. 镉在槐叶苹叶片中的蓄积及其生态毒理学分析. 生态学报, 2009, 29(6): 3019- 3027.
[8] 汪洪, 周卫, 林葆. 钙对镉胁迫下玉米生长及生理特性的影响. 植物营养与肥料学报 2001, 7(1): 78- 87.
[9] 陈晓玲, 余土元, 秦华明, 梁雪莲, 吴培钿, 伦锦华, 周玲艳. 钙对铬胁迫下玉米幼苗生长及生理特性的影响. 玉米科学 2009, 17(4): 74- 78.
[12] 张志良, 翟伟菁. 植物生理学实验指导 (第三版). 北京: 高等教育出版社, 2005.
[14] 张帆, 罗承德, 张健. 外源钙、磷、氮对铝胁迫下杉木幼苗生长影响的调控研究. 应用生态学报, 2005, 16(2): 213- 217.
[22] 唐蕾, 陈蕴, 王武. 银杏叶中叶绿素衍生物的反相高效液相色谱分析. 植物生理学通讯, 2006, 42(2): 265- 267.
[23] 沈成国, 张福锁, 毛达如. 植物叶片衰老过程中叶绿素降解代谢研究进展. 植物学通报, 1998, 15(增刊): 41- 46.
[25] 董彩霞, 周健民, 赵世杰, 王火焰. 外源钙对不同钙敏感型番茄幼苗生理特性的影响. 应用生态学报2005, 16(2): 267- 272.
[26] 邱栋梁, 刘星辉, 郭素枝. 模拟酸雨胁迫下钙对龙眼光合功能的调节作用. 应用生态学报, 2002, 13(9): 1072- 1076.
[27] 王爱国. 植物的氧代谢 // 于叔文, 汤章成主编.植物生理与分子生物学.北京:科学出版社,1988:366- 389.
[28] 肖细元, 廖晓勇, 陈同斌, 张杨珠. 砷、钙对蜈蚣草中金属元素吸收和转运的影响. 生态学报, 2003, 23(8): 1477- 1487.
[48] 闵海丽, 蔡三娟, 徐勤松, 施国新. 外源钙对黑藻抗镉胁迫能力的影响. 生态学报, 2012, 32(1): 256- 264.
Effects of exogenous calcium (Ca) on tolerance ofAlliumcepavar.agrogarumL. to cadmium (Cd) stress
WANG Qiaoling,ZOU Jinhua,LIU Donghua,YUE Jieyu*
TianjinKeyLaboratoryofAnimalandPlantRegulation,TianjinNormalUniversity,Tianjin300387,China
Cadmium (Cd) pollution is one of the most extensive and harmful environmental problems worldwide. Recently, coastal water pollution by Cd and other heavy metals have been becoming more serious because of continuous economic development, including mining, urban traffic, burning of fossil fuels, and production of phosphate fertilizers. Calcium (Ca) is an essential macroelement that plays a crucial role in regulating plant growth and development. Moreover, Ca has been shown to alleviate heavy metal toxicity in many plants.Alliumcepais well known and commonly used in many laboratories primarily due to its many advantages, such as low cost, short test time, ease of storage and handling, large number of roots, and large cells with excellent chromosome conditions. In the present study, this plant was chosen as the experimental material to investigate whether the application of exogenous Ca can enhance the tolerance ofA.cepavar.agrogarumL. to Cd stress (10,100 μmol/L and 300 μmol/L). In addition, the effects of various concentrations of exogenous Ca application (0, 0.1, 1, and 10 mmol/L) on the vegetative growth, photosynthesis characteristics, Cd accumulation, and nutrient minerals ofA.cepavar.agrogarumL. under various concentrations of Cd stress (10 μmol/L to 300 μmol/L) were analyzed. This study showed the following results: (1) Excess Cd significantly inhibited the growth ofA.cepavar.agrogarumL., resulting in changes in morphological characteristics, such as stunted and bent root tips and darkened green leaves, whereas exogenous Ca significantly decreased the negative effects of Cd and alleviated growth inhibition inA.cepavar.agrogarumL. (2) Cd treatment greatly increased the chlorophyll contents and significantly decreased net photosynthetic rate, stomatal conductance, and transpiration rate, whereas exogenous application of Ca markedly alleviated the increase in photosynthetic pigments and significantly increased the net photosynthetic rate, stomatal conductance, and transpiration rate. (3) Cd stress induced substantial Cd accumulation, resulting in serious nutrient imbalance, such as the significant inhibition of absorption of Mg ( Magnesium ) and Mn (Manganese ) in roots and Ca, Mg, Mn, and Zn ( Zinc ) in leaves and a massive accumulation of Ca, Fe (Iron) and Zn in roots and Fe in leaves, whereas the exogenous addition of Ca significantly decreased Cd accumulation and effectively reversed the Cd-induced imbalance of nutrients inA.cepavar.agrogarumL. Therefore, supplementing exogenous Ca with an optimal concentration can effectively relieve the phytotoxicity of Cd stress toA.cepavar.agrogarumL. by decreasing excessive Cd accumulation, maintaining high levels of photosynthesis characteristics, and maintaining the balance of water content, photosynthetic pigments, and nutrient elements, among others.
Alliumcepavar.agrogarumL.; Cd; Ca; stress; tolerance
天津市高等学校科技发展计划项目(20100606,20110603);天津市应用基础计划项目青年项目(12JCQNJC09700);天津师范大学校级基金(52XB1105,52XS1210)
2013- 05- 06;
2013- 09- 18
10.5846/stxb201305060949
*通讯作者Corresponding author.E-mail: yueshan1982@163.com
王巧玲,邹金华,刘东华,岳洁瑜.外源钙(Ca)对毛葱耐镉(Cd)胁迫能力的影响.生态学报,2014,34(5):1165- 1177.
Wang Q L,Zou J H,Liu D H,Yue J Y.Effects of exogenous calcium (Ca) on tolerance ofAlliumcepavar.agrogarumL. to cadmium (Cd) stress.Acta Ecologica Sinica,2014,34(5):1165- 1177.
猜你喜欢
舰船科学技术(2022年11期)2022-07-15
阅读(科学探秘)(2020年8期)2020-11-06
科学导报·学术(2020年19期)2020-07-09
西藏农业科技(2019年3期)2019-11-04
中国果业信息(2019年1期)2019-01-05
现代园艺(2018年3期)2018-02-10
生物学教学(2017年9期)2017-08-20
上海农业学报(2017年3期)2017-04-10
汉语世界(The World of Chinese)(2016年3期)2016-01-10
食品工业科技(2014年6期)2014-05-10
