
胰岛素诱导因子2基因rs7566605位点对中国老年人群肥胖及血脂影响
2014-04-11 10:46:18杨景慧刘建伟李小鹰
解放军医学院学报 2014年5期
杨景慧,刘建伟,刘 静,李小鹰,何 耀,叶 玲
解放军总医院,北京 100853 1老年医学研究所;2衰老及相关疾病研究北京市重点实验室;3解放军总医院老年心内科
高脂血症(Hyperlipidemia)作为一种脂质代谢的紊乱综合征,包括血清总胆固醇(total cholesterol,TC),甘油三酯(total triglycerides,TG)和高密度脂蛋白胆固醇(high-density lipoprotein cholesterol,HDL-c)水平的异常。以往流行病学和临床研究表明,高脂血症与冠状动脉疾病(Coronary Artery Disease,CAD)的风险增加显著相关[1-3]。高脂血症是一种由遗传因素和多种环境危险因素共同影响的复杂性疾病[4-6]。在欧洲,美国及日本等国家的大样本的全基因组关联(GWA)研究中[7-10],位于胰岛素诱导基因2(Insulin-induced gene 2,INSIG2)上游的单核苷酸多态性(Single nucleotide polymorphisms,SNP)位点(rs7566605)被确认与肥胖相关。然而,也有研究表明,该位点在东亚人群中与肥胖不相关,而对脂类代谢有一定作用[11]。细胞功能实验也表明,该基因的过度表达会导致血浆中甘油三酯水平的降低[12]。由于关于在中国人群该位点与血脂代谢的关系尚不明确,且老年人群有较高的血脂异常患病率,因此,本文旨在探讨在北京老年社区人群中该位点与肥胖及血脂代谢的关系。
研究对象和方法
1 研究对象 本研究对象来自2009年9月- 2010年6月间参加北京市海淀区万寿路地区老年人群健康情况调查并完成问卷填写、体格检查及基因分型的年龄≥60岁的老年人,共2 014例。本项研究已通过中国人民解放军总医院医学伦理委员会审查,所有研究对象均已签署知情同意书。
2 抽样方法 二阶段分层整群抽样。第一阶段,在万寿路社区全部街道或社区中(N≈300 ~ 600),随机抽取五个社区。第二阶段,抽取五个社区内全部符合纳入标准的研究对象,即能独立配合调查员调查,愿意提供血液样本且年龄≥60岁的老年人。对符合标准的研究对象,全部采用面对面问卷访谈方式,问卷设计合理,由调查员填写问卷内容,现场由流行病学专家进行严格质控。
3 调查内容 1)身体测量指标:身高、体重、腰围、臀围及血压,测量过程采用标准测量工具及方法;2)血液生化指标:空腹血糖、总胆固醇、甘油三酯、高密度脂蛋白胆固醇、餐后2 h血糖等,使用标准试剂盒进行检测(Boehringer Mannheim公司GMBH);3)环境因素:受教育程度,运动情况,烟草和酒精使用等。饮酒定义为在过去的5年内,平均每日饮酒量超过10g且持续时间>1年;吸烟定义为连续或累积吸烟≥6个月。
4 诊断标准 1)高脂血症:高胆固醇血症,血清总胆固醇≥5.72vmmol/L(220 mmg/dL);高甘油三酯血症,血清甘油三酯≥1.7 mmol/L(150 mmg/dL);或低HDL-c血症,高密度脂蛋白胆固醇,男性血清HDL-c<1.03 mmol/L(40 mmg/dL),女性血清HDL-c<1.3 mmol/L(50 mmg/dL)或已接受降脂治疗。满足以上任何一种即诊断为高脂血症。2)肥胖:体重指数(BMI)≥25 kg/m2[13]。
5 基因提取及分型 DNA标本通过标准蛋白酶K法从外周静脉血样品中提取。使用美国Sequenom公司的MassARRAY系统进行基因分型。
6 统计学方法 采用SPSS16.0版软件包进行统计分析。正态连续性资料以±s表示。正态计量资料的组间差异比较采用独立样本t-检验或方差分析。计数资料采用Pearsonχ2或Fisher确切概率法,利用多因素Logistic回归模型评估疾病影响因素的比值比(Odds ratio,OR)和95%置信区间(95% Confidence Interval,95% CI)。采用Haploview 4.2软件(http://www.broadinstitute.org/haploview)对基因分型数据进行Hardy-Weinberg平衡(HWE)检验,α=0.05。
结 果
1 一般资料 本研究最终完成问卷并分型成功者共2 014例,rs7566605的基因型在本人群中的频率分别为GG 42.9%,GC 45.4%和CC 11.8%。基因分型结果符合Hardy-Weinberg平衡。女性腰围,收缩压值均高于男性(P<0.001),见表1。
2 基因与空腹血糖及血脂水平的关系 随着等位基因位点C数量的增加,男性甘油三酯血清水平显著升高,男女总人群的HDL-c水平逐渐降低,而在女性人群中,虽然也可以观察到相同的趋势,但差异无统计学意义,见表2。
3 基因与肥胖及血脂代谢异常疾病关系 采用多因素logistic回归对该基因对肥胖、高脂血症及其组分的影响进行分析,遗传模型选择加性模型,BMI、年龄、性别及环境因素作为调整因素。结果显示,在该老年人群中,任何基因型个体的高脂血症患病率均较高(GG 73.3%,GC 78.4%,CC 73.4%),差异无统计学意义。SNP rs7566605的C位点变异可以显著增加高甘油三酯血症及低HDL-c血症的患病风险(P=0.019,OR=1.175和P=0.020,OR=1.178)。然而,在本人群研究中,该位点未发现与肥胖有任何统计学关联,见表3。
讨 论
位于INSIG2基因上游的SNP rs7566605最初由Herbert等[7]研究发现与体重指数存在关联,并在四个其他人群队列中对结果进行了重复性验证。此外,Lyon等[14]也在五个人群队列中证实该SNP与肥胖之间显著关联,但在另外三个队列中未获得相同结论。中国儿童相关研究中也未发现该位点对BMI升高或体脂含量的增加存在显著作用[15]。我们将人群根据WHO推荐的亚洲人群BMI分类标准进行进一步分层分析,发现在肥胖个体(BMI>28 kg/m2)中rs7566605与肥胖显著相关。以往其他研究也曾获得过类似的结果,推测该位点对体重指数变化的影响在肥胖人群中可能更加显著[11,16-17],由此可以一定程度上解释体型偏瘦的亚洲人群无法获得关联结果的原因。
INSIG基因功能已经在小鼠和细胞水平上得以证实[12,18],该基因通过作用于固醇调节元件结合 蛋 白(sterol regulatory element-binding proteins,SREBPs)调节胆固醇运输,SREBPs作为一种合成转录因子可以激活总胆固醇和甘油三酯在肝脏及其他器官内合成。由此推测该基因可能通过参与机体内脂类代谢,导致肥胖或血脂水平的改变。然而,关于该位点与人类脂类代谢的关系研究尚少,中国人群中尚未发现相似关联。研究显示从总体趋势看,等位基因C与血清TG和FPG水平的升高及TC和HDL-c的减少可能相关。但仅男性TG及联合样本的HDL-c水平基因型间差异有统计学意义。女性中未发现显著性结果,可能由于男性接触更多的环境因素,如吸烟、饮酒等。为排除性别及环境因素的干扰,我们利用多因素调整的logistic回归模型分析rs7566605与血脂异常疾病的关系。如表3所示,C等位基因与高脂血症,高甘油三酯血症和低高密度脂蛋白血症患病增加显著相关。而未发现与高胆固醇血症关联有统计学意义。此外,我们观察到,相对于CC及GG纯合子,杂合子CG似乎对血脂水平影响更明显。值得注意的是,在韩国女性人群相同研究中,C位点被认为是与血清胆固醇升高相关,并也可观察到杂合子个体血清总胆固醇的平均水平要高于另外两种纯合体,结果与本研究相反[19]。基于以上研究结论,我们推测该基因对脂类代谢性疾病的影响可能遵循杂合共显性模型。
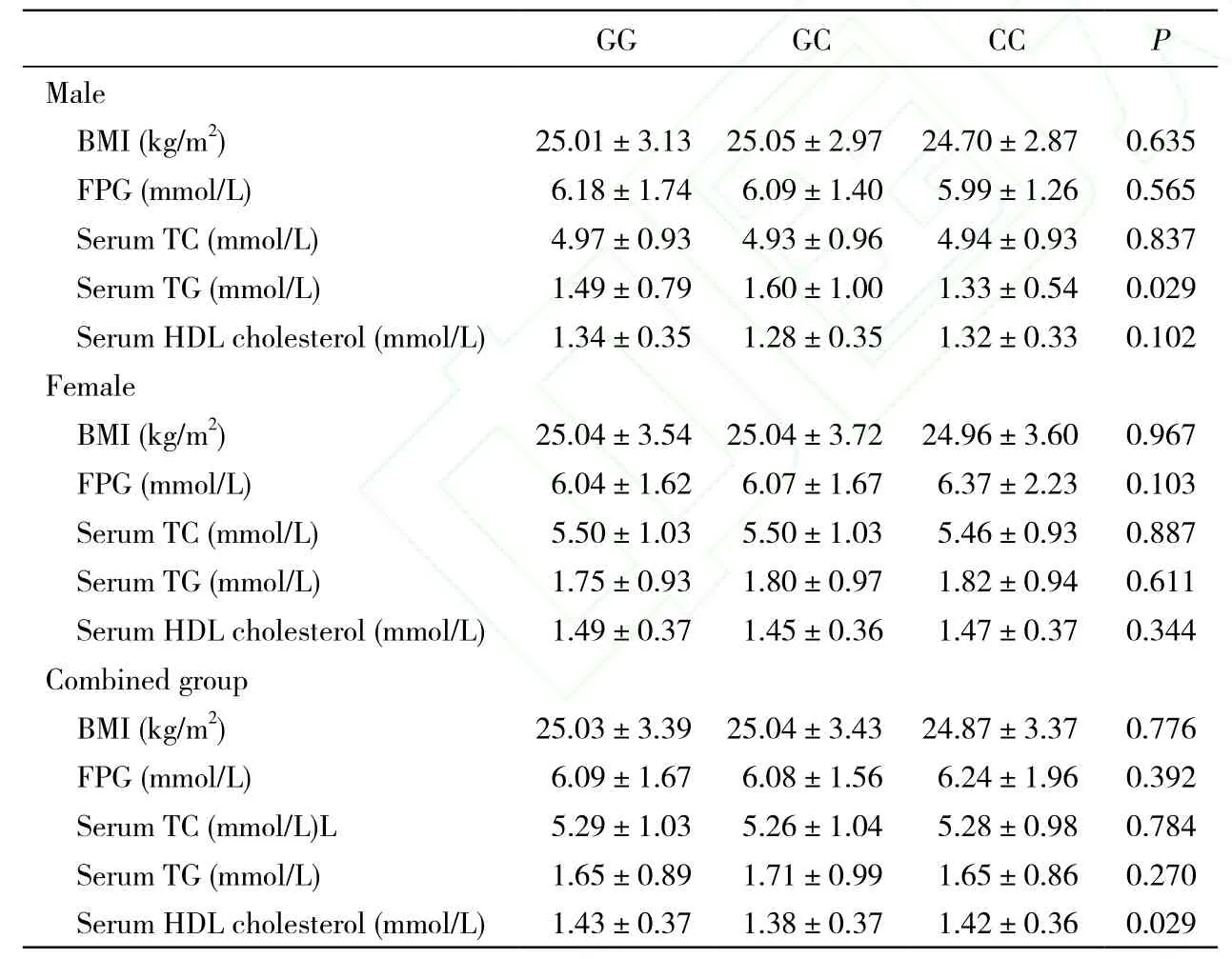
表2 rs7566605位点不同基因型血清脂类及空腹血糖水平差异Tab. 2 Serum lipid and fasting blood glucose levels in different genotypes of rs7566605(n=2 014)
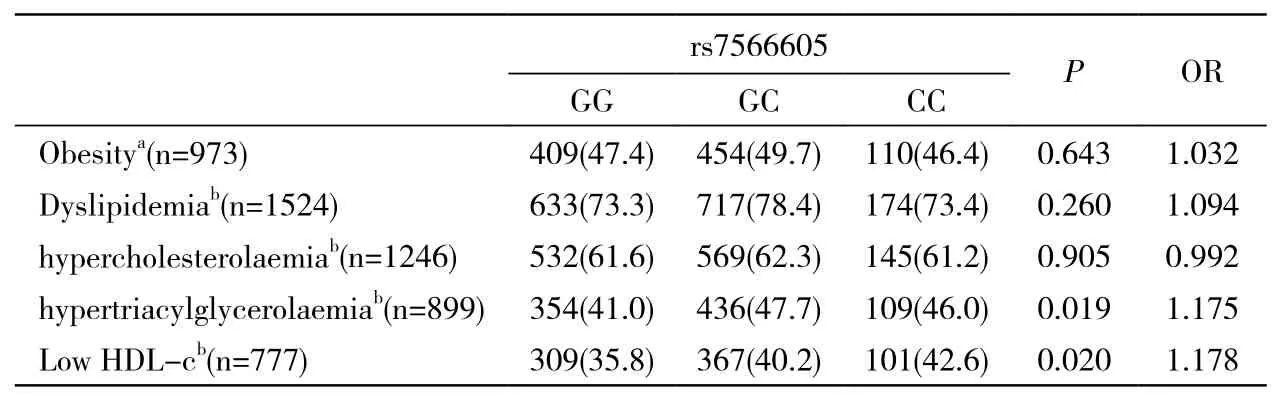
表3 rs7566605位点与肥胖及血脂代谢异常疾病关联性分析Tab. 3 Association of rs7566605 genotypes with obesity and abnormal lipid metabolism-related diseases
以往研究观察到,CC基因型是对日裔美国女性高胆固醇血症的发病具有保护作用,而在日本本土人群样本中未发现相同结果[11]。然而,同作为东亚人群的韩国女性样本则表现出C位点可以导致胆固醇水平的升高[19]。在一项以黑人及白人作为研究对象的队列研究证实,INSIG2基因上多态性rs1352083和rs10185316对HDL-c水平的降低程度随研究对象年龄的升高而更加明显[20]。观察以上几组样本特征,可以发现日裔美国人和本研究对象的年龄要明显高于日本本土及韩国的样本,这表明年龄可能作为一个不可忽略的影响因素调节基因与脂类代谢的关系,从而导致结果的不一致性。然而,年龄的影响作用机制还需要进一步研究验证。
遗憾的是,我们没有收集相应的饮食资料,也未在研究过程中进行严格的饮食控制,导致的结论可靠性降低。此外,INSIG2基因功能受血清胰岛素水平的影响,而我们的研究未能获得这方面资料,无法观察其对结果的影响。
综上所述,作为中国人群中获得INSIG2基因多态性rs7566605与血脂异常,包括高胆固醇血症,高甘油三酯血症及低HDL-c血症关联的首个研究,我们发现SNP rs7566605的C等位基因与高甘油三酯血症和低高密度脂蛋白血症患病风险呈正相关,同时表现出对高胆固醇血症的保护性趋势。杂合共显性模型可能是该基因遗传的主要模式。在本人群中,多态性rs7566605与肥胖之间也未发现统计学关联。不过,仍需要更大样本的、具有严格饮食资料的纵向研究对本研究所获得结论进行进一步的验证,以期获得更加准确结论。
1 Achari V, Thakur AK. Association of major modifiable risk factors among patients with coronary artery disease--a retrospective analysis[J]. J Assoc Physicians India, 2004, 52: 103-108.
2 Holewijn S, den Heijer M, Swinkels DW, et al. Apolipoprotein B,non-HDL cholesterol and LDL cholesterol for identifying individuals at increased cardiovascular risk[J]. J Intern Med, 2010, 268(6):567-577.
3 Austin MA, Hokanson JE, Edwards KL. Hypertriglyceridemia as a cardiovascular risk factor[J]. Am J Cardiol, 1998, 81(4A):7B-12B.
4 Ruixing Y, Jinzhen W, Weixiong L, et al. The environmental and genetic evidence for the association of hyperlipidemia and hypertension[J]. J Hypertens, 2009, 27(2): 251-258.
5 Ruixing Y, Qiming F, Dezhai Y, et al. Comparison of demography,diet, lifestyle, and serum lipid levels between the Guangxi Bai Ku Yao and Han populations[J]. J Lipid Res, 2007, 48(12):2673-2681.
6 Ruixing Y, Jinzhen W, Yaoheng H, et al. Associations of diet and lifestyle with hyperlipidemia for middle-aged and elderly persons among the Guangxi Bai Ku Yao and Han populations[J]. J Am Diet Assoc, 2008, 108(6): 970-976.
7 Herbert A, Gerry NP, Mcqueen MB, et al. A common genetic variant is associated with adult and childhood obesity[J]. Science, 2006,312(5771): 279-283.
8 Yang Q, Lai CQ, Parnell L, et al. Genome-wide linkage analyses and candidate gene fine mapping for HDL3 cholesterol: the Framingham Study[J]. J Lipid Res, 2005, 46(7): 1416-1425.
9 Harrap SB, Wong ZY, Scurrah KJ, et al. Genome-wide linkage analysis of population variation in high-density lipoprotein cholesterol[J]. Hum Genet, 2006, 119(5): 541-546.
10 Kathiresan S, Melander O, Guiducci C, et al. Six new loci associated with blood low-density lipoprotein cholesterol, high-density lipoprotein cholesterol or triglycerides in humans[J]. Nat Genet,2008, 40(2): 189-197.
11 Oki K, Yamane K, Kamei N, et al. The single nucleotide polymorphism upstream of insulin-induced gene 2 ( INSIG2) is associated with the prevalence of hypercholesterolaemia, but not with obesity, in Japanese American women[J]. Br J Nutr, 2009, 101(3):322-327.
12 陈科,莫朝晖,邢晓为,等.INSIG2稳定表达细胞系的建立及该基因对脂肪代谢的影响[J].基础医学与临床,2008,28(7):713-718.
13 IASO International Association for the Study of Obesity. The Asia-Pacific perspective: Redefining obesity and its treatment[S/ OL]. http://www.wpro.who.int/nutrition/documents/docs/ Redefiningobesity.pdf
14 Lyon HN, Emilsson V, Hinney A, et al. The association of a SNP upstream of INSIG2 with body mass index is reproduced in several but not all cohorts[J]. PLoS Genet, 2007, 3(4): e61.
15 张衡,王海俊,马军,等.青少年肥胖及代谢异常与INSIG2基因多态性[J].中国公共卫生,2009,25(5):537-540.
16 Hall DH, Rahman T, Avery PJ, et al. INSIG-2 promoter polymorphism and obesity related phenotypes: association study in
1428 members of 248 families[J]. BMC Med Genet, 2006, 7: 83.
17 Smith AJ, Cooper JA, Li LK, et al. INSIG2 gene polymorphism is not associated with obesity in Caucasian, Afro-Caribbean and Indian subjects[J]. Int J Obes(Lond), 2007, 31(11):1753-1755.
18 Engelking LJ, Liang G, Hammer RE, et al. Schoenheimer effect explained--feedback regulation of cholesterol synthesis in mice mediated by Insig proteins[J]. J Clin Invest, 2005, 115(9):2489-2498.
19 Cha S, Koo I, Choi SM, et al. Association analyses of the INSIG2 polymorphism in the obesity and cholesterol levels of Korean populations[J]. BMC Med Genet, 2009, 10: 96.
20 Fornage M, Papanicolaou G, Lewis CE, et al. Common INSIG2 polymorphisms are associated with age-related changes in body size and high-density lipoprotein cholesterol from young adulthood to middle age[J]. Metabolism, 2010, 59(8): 1084-1091.
猜你喜欢
老年人(2022年8期)2022-04-29 00:44:03
锦州医科大学报(2021年8期)2021-11-18 09:11:53
中老年保健(2021年4期)2021-08-22 07:07:36
家庭医学(下半月)(2019年9期)2019-10-12 08:03:56
中医眼耳鼻喉杂志(2019年2期)2019-04-13 05:23:46
祝您健康(2018年12期)2018-11-27 02:30:34
中国卫生标准管理(2015年24期)2016-01-14 09:29:03
分子影像学杂志(2015年3期)2015-12-04 03:28:59
中国当代医药(2015年21期)2015-03-01 02:05:07
疑难病杂志(2014年12期)2014-04-16 05:19:33
