
嗜冷黄杆菌酰基-Acp去饱和酶(AAD)的序列分析和结构建模
2014-03-23 02:20茆灿泉
生物学杂志 2014年4期
张 远, 茆灿泉
(西南交通大学 生命科学与工程学院, 成都 610031)
不饱和脂肪酸是生物膜的重要组成成分,同时也是生物体内信号传递、能量储备的前体物质,对于维持生物体的正常运行起到不可替代的作用。脂肪酸去饱和酶可以催化生物体内饱和脂肪酸的去饱和,根据作用底物的不同可以分为3类:分别为酰基载体蛋白去饱和酶、酰基-脂去饱和酶和酰基辅酶A去饱和酶[1]。酰基-Acp(酰基载体蛋白)去饱和酶(acyl-acyl carry protein desaturase, AAD)催化饱和脂肪酸链9和10位碳原子间碳碳单键形成双键,催化形成单不饱和键的脂肪酸是多不饱和脂肪酸的前体物质,很大程度上决定了多不饱和脂肪酸在生物体内的相对含量,从而影响整个生物体的生命状态。
细胞膜中膜脂的含量决定着细胞膜的流动性,低温条件下,多数微生物通过改变膜脂中的多不饱和脂肪酸含量来维持细胞膜的流动性。Weinstein[2]等研究表明,低温条件下培养部分真菌菌株,其细胞膜中多不饱和脂肪酸(PUFAs) 的含量会增加,其中长孢被孢霉(mortierellaelongata)会合成十八碳四烯酸,后者主要存在于低温环境下的微生物中。Wada[3]利用点突变使蓝细菌Δ12-脂肪酸脱氢酶失活,改变了细胞膜磷脂组成,使细胞在22℃培养时生长明显减慢,进一步把蓝细菌的Δ12-脂肪酸脱氢酶基因导入到对低温敏感的蓝细菌中表达,结果该蓝细菌中亚油酸含量明显增加,对5℃低温的抗性明显增强,进一步证明了脂肪酸去饱和酶在维持细菌低温耐受的关键作用。陆合[4]等通过低温胁迫根霉R31.6,发现其抵抗低温胁迫的主要机制是通过增加膜上不饱和脂肪酸的含量,以此来保证低温下膜的流动性。于爱群[5]等研究低温对于高山被孢霉的3种脂肪酸脱氢酶的影响,结果发现低温对于高山被孢霉的3种脂肪酸脱氢酶的基因有转录激活作用。
嗜冷黄杆菌(Flavobacteriumpsychrophilum)是一种在低温环境下生存的极端环境微生物,其温度生长范围在4℃~25℃,最适生长温度在21℃。不饱和脂肪酸的相对含量对于细胞膜的流动性起到重要作用,而AAD又是多不饱和脂肪酸合成的关键酶,因此AAD在低温环境下的活性状况对于嗜冷黄杆菌的新陈代谢将非常关键。虽有不同物种脂肪酸去饱和酶研究的多个报道[6],但对于嗜冷黄杆菌的脂肪酸去饱和酶的三维结构解析以及蛋白序列结构中的活性位点分析却未见报道。本研究利用生物信息学的技术与方法对嗜冷黄杆菌AAD的序列组成与理化特性、跨膜和功能结构区、系统发育、保守基序、三维结构特征和活性位点等进行分析,为后续深入研究细菌耐低温机制及应用奠定基础。
1 材料与方法
嗜冷黄杆菌AAD序列来自于NCBI中protein数据库(登录号:YP_001294991.1),模板序列来自于PDB数据库(登录号:1oq9),用于系统发育及保守基序分析的序列均来自于NCBI中protein数据库。使用ProtParam(http://web.expasy.org/protparam/)、Protscal(http://web.expasy.org/protscale/)、PSORT(http://psort.hgc.jp/)、Tmpred(http://www.ch.embnet.org/software/TMPRED_form.html)、Interproscan 4(http://www.ebi.ac.uk/Tools/pfa/iprscan/)对目标蛋白序列AAD进行氨基酸序列分析;应用ClustalX软件进行蛋白序列的多序列比对,MEGA4.1构件系统发育树;利用weblogo(http://weblogo.berkeley.edu/logo.cgi)分析不同物种中脂酰去饱和酶的保守基序;AAD的三维结构由swiss-model(http://swissmodel.expasy.org/)同源建模完成,使用procheck、Verify 3D(http://nihserver.mbi.ucla.edu/SAVES/)对构建的模型进行评价;Swiss-Pdb Viewer用于显示和分析三维结构模型。
2 结果与分析
2.1 氨基酸序列分析
Expasy ProtParam tool分析显示,嗜冷黄杆菌AAD序列全长329个氨基酸,理论等电点pI=5.08,分子量38.42 kD,含酸性氨基酸asp和glu共51个,碱性氨基酸arg和lys共38个,为偏酸性稳定蛋白质。Protscal分析表明AAD为亲水性蛋白[7]。PSORT分析AAD定位于细胞质中,这与Tmpred分析的AAD为非跨膜结构蛋白一致。Interproscan 4分析AAD表明其具备功能delt9-acylAcp-des区以及Fe离子结合区,推断AAD合成后直接在细胞质中与Fe离子结合形成反应复合物,催化9与10位上的碳碳单键形成双键。
2.2 系统发育分析
在NCBI蛋白数据库中检索出不同物种的AAD蛋白序列24个,分别为黄杆菌属(Flavobacteriumfrigoris,WP_007136780.1)、黄杆菌属(FlavobacteriumpsychrophilumJIP02/86,YP_001294991.1)、鲍氏不动杆菌属(AcinetobacterbaumanniiOIFC111,EKP44864.1)、互生平胞杆菌(Alteromonasmacleodiistr. ′BlackSea11′,YP_006826131.1)、淀粉丝菌(Amylomycesrouxii,AAB82294.1)、拟南芥(Arabidopsisthaliana,AEE27937.1)、PolysphondyliumpallidumPN500 (PolysphondyliumpallidumPN500,EFA75230.1)、CapsasporaowczarzakiATCC 30864 (XP_004364396.1)、网柱菌 (DictyosteliumdiscoideumAX4,XP_636528.1)、播娘蒿属(Descurainiasophia,ABS86965.1) 、弗朗西斯菌属(Francisellacf.novicida3523,YP_005824683.1)、伽玛变形菌属(gammaproteobacteriumIMCC3088,WP_009574453.1)、柯柯豆毛色二孢(Lasiodiplodiatheobromae,ABQ00180.1)、被孢霉菌(Mortierellaalpina, ADE06659.1)、 亚硝化螺菌属(Nitrosospirasp.APG3,CCU63342.1)、糙皮侧耳(Pleurotusostreatus,ACQ73330.1)、核腔菌属(Pyrenophoratritici-repentisPt-1C-BFP,EDU49212.1)、蓖麻(Ricinuscommunis,EEF44805.1)、蔷薇属(Rosahybridcultivar,BAA23136.1)、管圆线虫(Angiostrongyluscantonensis,AEB96387.1)、土拉热弗朗西斯菌(Francisellatularensissubsp.tularensisTI0902,YP_005317387.1)、梭孢壳属(ThielaviaterrestrisNRRL8126,AEO66985.1)、油桐属(Verniciafordii,ADC80920.1)、unculturedmarinegroupIIeuryarchaeote(EHR76326.1)。
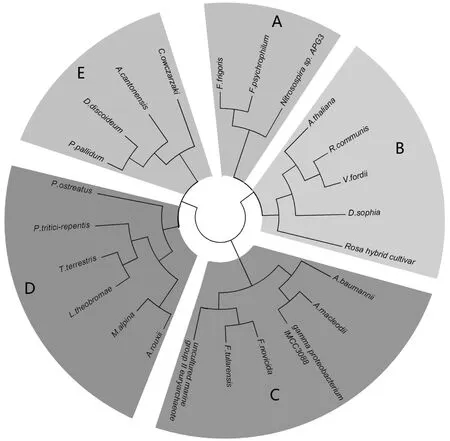
图1 不同物种中脂酰去饱和酶的系统进化树
由图1分析可见,此系统发育树分为5个分支[8],嗜冷菌F.frigoris、F.psychrophilumJIP02/86和细菌Nitrosospirasp.APG3同源关系较近并聚为一个分支A、B为植物类分支,C为细菌类分支,D为真菌类分支,E为动物类分支,其中与C类分支较近的unculturedmarinegroupIIeuryarchaeote为古细菌。
2.3 保守基序分析
利用weblogo分析不同物种中AAD的一致或相似区[9]。weblogo是将多序列比对的结果以一种比较直观的方式表现出来的软件,如图2所示,字母越大表示此位点该氨基酸一致性越高;反之一致性越低,保守性也越差。从图2可以看出,His102、His107和his139、his142、his143以及his303、his306、his307组成HxxxxH、HxxHH、HxxHH(图2箭头表示)3个组氨酸保守区,这3个区域是铁离子结合区,8个组氨酸任何一个的突变都可能导致整个蛋白酶失去活性[10]。
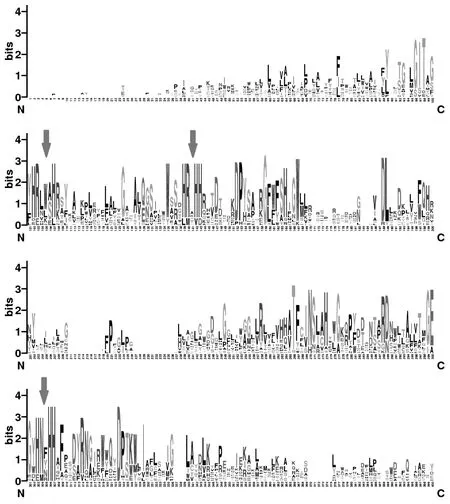
图2 不同物种中脂酰去饱和酶的保守基序分析

图3 SAD与AAD的序列比对及二级结构分布
三角符号表示与二铁离子相互作用的活性位点区。
2.4 分子建模
将AAD蛋白序列在NCBI中进行BLAST分析,结果显示,在所有的三维结构中,嗜冷黄杆菌AAD与蓖麻子硬脂酰-ACP去饱和酶(Stearoyl Acyl Carrier Protein Desaturase, SAD)的同源一致性为49%,序列覆盖为96%,而且与SAD与AAD的分子进化关系较近,因此选定蓖麻子SAD作为同源建模模板。其与嗜冷黄杆菌AAD的序列比对结果见图3。利用swiss-model[11]进行同源模建得到嗜冷黄杆菌AAD的三维结构,其三维结构含18个ɑ-螺旋,2个β-折叠。SAD与同源建模得到的AAD三维结构重叠如图4所示,二者呈现较高的相似性,其重叠的均方根偏差(RMSD)为1.5 Å。

图4 SAD与AAD三维结构重叠
2.5 分子建模评估
采用PROCHECK和Verify 3D对建立的嗜冷黄杆菌AAD三维结构模型进行评估。PEOCHECK通过分析残基与残基间的几何结构以及整体的几何结构来评估蛋白结构的立体质量,Verify 3D依据原子模型所处的位置和环境(α-螺旋,β-折叠,无规则卷曲,极性,非极性等)以及与适当结构的比较从而决定原子模型与自身的氨基酸序列的兼容性。PEOCHECK得到的拉式图(见图5)里所示,99.3%的氨基酸的结构处于合理区,仅有0.7%的氨基酸位于合理区外,说明同源建模得到的AAD三维结构模型立体几何结构较为合理。同样地,通过Verify 3D评价[12],超过80%的氨基酸评分大于0,也证明了同源建模得到的AAD结构模型其三维结构构象的合理性评价良好。
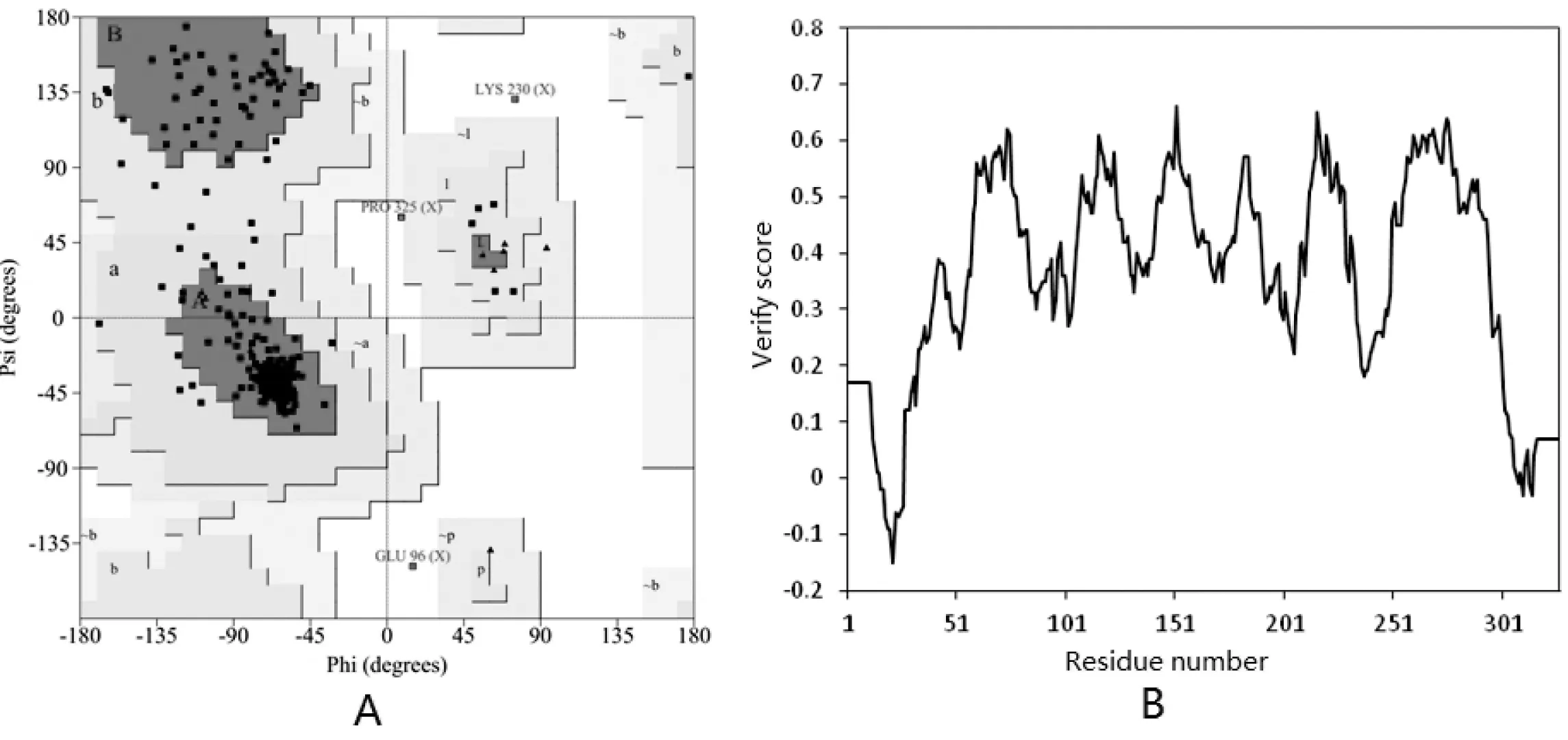
图 5 AAD三维结构模型的评估
Fig 5 Evaluation of the three-dimensional structure model of AAD
A—AAD三维结构模型的拉氏图,其中“A, B, L”为核心区,“a, b, l, p”为允许区,“~a, ~b, ~l, ~p”为额外接受区域,空白区为不合理区域;B—AAD三维结构模型的Verify-3D图,分值大于0的为合理构象。
2.6 活性位点分析
将AAD与SAD三维结构重叠,从AAD上找到了与SAD一致的活性位点,主要有His114、Glu111、Glu76、His200、Glu197、Glu164,与SAD上面的活性位点高度保守(图3),其中包含两个螯合铁离子Fe1、Fe2,Fe1与His114 、Glu111 、Glu76相互作用[13],Fe2与His200、Glu197、Glu164作用。铁离子对于维持整个蛋白活性起到重要作用,铁离子的缺失将会造成活性位点碳侧链的构象紊乱,从而导致AAD活性丧失。
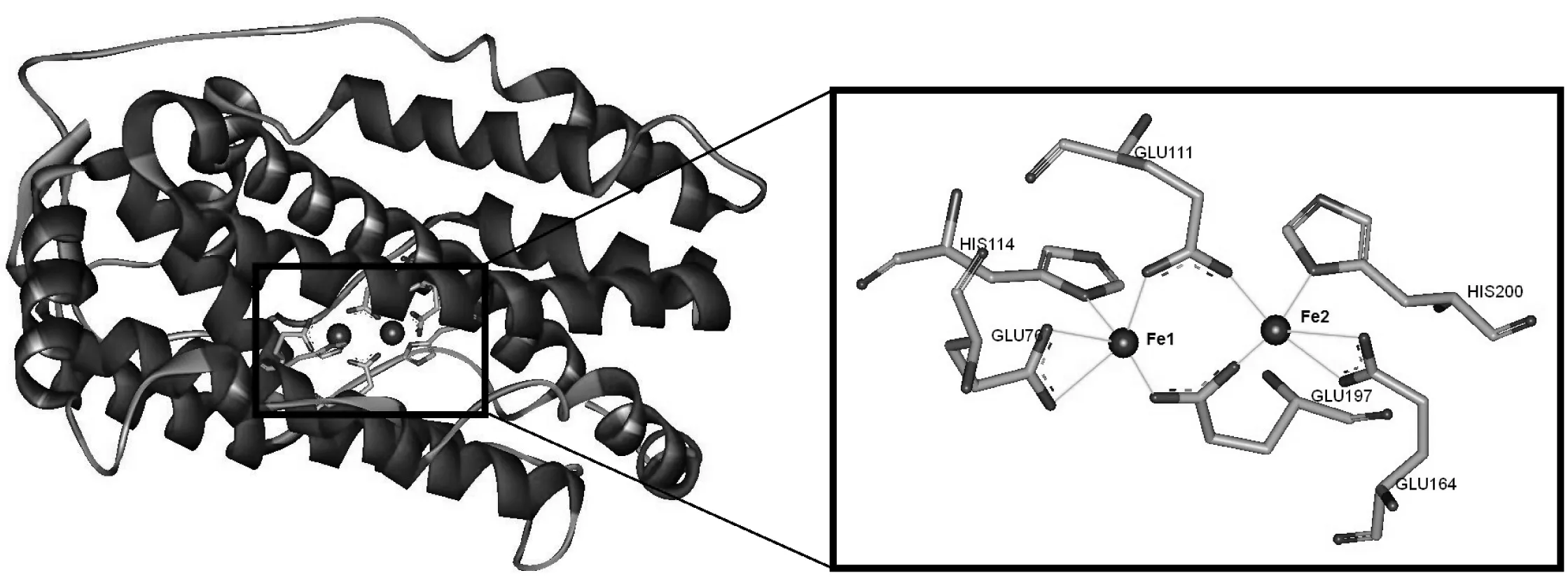
图6 活性位点与铁离子的相互作用及立体构象
3 讨论
微生物在低温条件下的耐受与活性机制一直都是嗜冷微生物研究的热点,不饱和脂肪酸对于低温情况下维持微生物细胞的流动性起到非常重要的作用。本研究以低温微生物嗜冷黄杆菌中与多不饱和脂肪酸代谢重要相关的蛋白AAD为研究对象,就其序列组成与基序特征、系统进化与重要活性位点中心,三维结构模建及评价等展开了广泛和深入的研究。系统发育分析发现,不同物种的脂肪酸去饱和酶差异很大,嗜冷黄杆菌等低温耐受菌与常温细菌之间的AAD分子进化关系较远,反而与植物类物种的SAD分子进化关系较近。之后的保守基序分析表明大多物种的脂肪酸脱氢酶都具备HxxxxH、HxxHH、HxxHH 3个组氨酸保守区,这与该保守区能与二铁离子结合[14],对酶的活性产生重要影响。将AAD蛋白序列在NCBI中用BLAST分析,结果显示在所有的三维结构中,AAD与SAD的同源一致性为49%,序列覆盖度为96%,而其它解析结构的序列覆盖度都不超过50%,而且SAD与AAD的分子进化关系较近,因此选定蓖麻子SAD作为同源建模模板。之后的三维结构建模及活性位点分析发现,AAD与SAD的同源一致性非常高,两者之间的序列比对分析发现,SAD的二铁离子结合区[13],在AAD中也同样存在。结合嗜冷黄杆菌AAD与常温细菌的分子进化关系较远,而与植物SAD的分子进化关系较近,并且二铁离子结合区的活性位点更接近于植物SAD的活性位点,可以推测相对于常温菌而言,低温菌中的低温耐受相关的基因可能发生了一定的变异。
本实验室目前已从环境污水中筛选获得一株较强低温耐受性的微小杆菌(Exiguobacteriumsibiricum)并从中克隆和获得了脂肪酸去饱和酶(fatty acid desaturase)基因,而对AAD的生物信息分析发现,大多物种由His102、His107、His139、His142、His143、His303、His306和His307组成的二铁离子结合区与嗜冷黄杆菌AAD由His114、Glu111、Glu76、His200、Glu197、Glu164组成的二铁离子结合区存在着氨基酸差异,而二铁离子结合区对于脂肪酸去饱和酶的活性影响很大。这为深入开展脂肪酸脱氢酶的结构改造与功能研究提供了方向,也为了解低温菌低温耐受机制奠定了重要的前期工作基础。
参考文献:
[1]Pereira S L, Leonard A E, Mukerji P. Recent advances in the study of fatty acid desaturases from animals and lower eukaryotes[J]. Prostaglandins, Leukotrienes and Essential Fatty Acids, 2003, 68(2): 97-106.
[2]Weinstein R N, Montiel P O, Johnstone K. Influence of growth temperature on lipid and soluble carbohydrate synthesis by fungi isolated from fell field soil in the maritime Antarctic[J]. Mycologia, 2000: 222-229.
[3]Wada H, Combos Z, Murata N. Enhancement of chilling tolerance of a cyanobacterium by genetic manipulation of fatty acid desaturation[J]. Nature, 1990, 13, 347(6289):200-203.
[4]陆 合, 张学昆, 李加纳, 等. 逆境对真菌膜脂肪酸成分的影响[J]. 微生物学杂志, 2005, 25(2): 1-3.
[5]于爱群, 石桐磊, 张 飙, 等. 低温和外源不饱和脂肪酸对高山被孢霉脂肪酸脱氢酶基因表达的影响[J]. 微生物学报, 2012, 52(11): 1369-1377.
[6]曹英萍, 石金磊, 李 钟, 等. 水稻 的克隆及其家族成员对非生物胁迫的响应[J]. 遗传, 2010, 32(8): 839-847.
[7]Zhang H W, Sun X F, He Y N, et al. Bioinformatics analysis of breast cancer bone metastasis related gene-CXCR4[J]. Asian Pacific Journal of Tropical Medicine, 2013, 6(9): 732-738.
[8]Muto M, Kubota C, Tanaka M, et al. Identification and functional analysis of delta-9 desaturase, a key enzyme in PUFA synthesis, isolated from the oleaginous diatom Fistulifera[J]. PloS One, 2013, 8(9): e73507.
[9]Chakraborty C, Tomar J, Kumar Gera V. Conserved domains, residues, webLogo and active sites of caspase-cascades related to apoptotic signaling pathway[J]. Current Bioinformatics, 2012, 7(4): 392-401.
[10]Bordoli L, Schwede T. Automated protein structure modeling with SWISS-MODEL Workspace and the Protein Model Portal[M]//Homology Modeling. Humana Press, 2012: 107-136.
[11]Kiefer F, Arnold K, Knzli M, et al. The SWISS-MODEL Repository and associated resources[J]. Nucleic Acids Research, 2009, 37(suppl 1): D387-D392.
[12]Petrey D, Xiang Z, Tang C L, et al. Using multiple structure alignments, fast model building, and energetic analysis in fold recognition and homology modeling[J]. Proteins: Structure, Function, and Bioinformatics, 2003, 53(S6): 430-435.
[13]Moche M, Shanklin J, Ghoshal A, et al. Azide and acetate complexes plus two iron-depleted crystal structures of the di-iron enzyme Δ9 stearoyl-acyl carrier protein desaturase implications for oxygen activation and catalytic intermediates [J]. Journal of Biological Chemistry, 2003, 278(27): 25072-25080.
[14]Whitney H M, Michaelson L V, Sayanova O, et al. Functional characterisation of two cytochrome b 5-fusion desaturases fromAnemoneleveillei: the unexpected identification of a fatty acid Δ6-desaturase[J]. Planta, 2003, 217(6): 983-992.
猜你喜欢
上海金属(2021年6期)2021-12-02
昆明医科大学学报(2021年3期)2021-07-22
中国生殖健康(2020年5期)2021-01-18
生物学通报(2019年3期)2019-02-17
中国生殖健康(2018年5期)2018-11-06
天然产物研究与开发(2018年9期)2018-10-08
中学科技(2016年5期)2016-05-12
合成化学(2015年10期)2016-01-17
中国洗涤用品工业(2015年8期)2015-02-28
西安交通大学学报(医学版)(2015年2期)2015-02-28
