
藻体部位、藻质量、碳源、水流对铁钉菜和羊栖菜营养盐吸收的影响*
2014-03-09 03:40:49孟伟杰桂福坤吴常文
海洋与湖沼 2014年3期
孟伟杰 王 萍① 桂福坤 吴常文 梁 君
(1.浙江海洋学院海洋科学学院 国家海洋设施养殖工程技术研究中心 舟山 316022;2.浙江省海洋水产研究所 舟山 316004)
大型藻类在人类活动中起着非常重要的作用,它们是食物、饲料、肥料、医药等的来源(Gregory,2005)。大型藻类作为生物过滤器可以有效地吸收、利用养殖环境中多余的营养物质,从而减轻养殖废水对环境的影响,是控制水体富营养化、增进食品安全和对污染水体进行生物修复的有效措施之一(毛玉泽等,2005)。近几年这方面的研究方兴未艾(Day,2003;Sandneset al;2005;李恒等,2012;陈聚法等,2012)
铁钉菜(Ishige foliaceaokamurai)在中国东海、南海分布广泛,民间用于喉炎、淋巴结炎、甲状腺肿的治疗,具有清热解毒、软坚散结的作用,可食用(张金鼎,1998);羊栖菜(Sargassum fusiforme)系暖温带海藻,在民间早已入药,具有“软坚散结、消痰、清凉解毒、破血祛淤、利水消肿”之功能,用于治疗瘿瘤,痰饮水肿等症(王勤等,2007)。目前针对铁钉菜的研究主要是化学成分分析和提取物生物活性方面的报道(冯美霞等,2011;林雄平等,2012;汤海峰等,2003;张迪等,2009);而对羊栖菜的研究主要集中在育种、营养与药用价值方面(李再亮等,2011)。鉴此,文章主要研究了藻体部位、藻质量、碳源浓度和水流流速等因素对这2种大型海藻氮磷营养盐吸收的影响,以期为生态环境修复提供科学参考依据。
1 材料与方法
1.1 海藻的来源与预培养
铁钉菜、羊栖菜采自舟山朱家尖水域,均是成体藻体。选择健康藻体,除去表面附着物,用过滤海水冲洗干净,移到有光照的水族箱中暂养,水族箱的容积为 10L,在 12L︰12D,1500µE/(m2*s),20°C 条件下,放入沉淀自然加富消毒海水[c(N)100µmol/L,c(P) 8µmol/L,f/2微量元素及维生素]中暂养1周,每隔2 d添加微量元素1次(包杰等,2008)。
1.2 试验项目
1.2.1 藻体部位对营养盐吸收的影响 铁钉菜藻体部位设为藻体顶端、中部、下部、固着器和全株藻体;羊栖菜藻体部位设为藻体顶端、中部、下部、固着器、气囊和全株藻体(陈丽梅等,2008)。试验环境因子设为 20°C、1500µE/(m2s)。培养液硝态氮(NO3-N)、铵态氮(NH4-N)和无机磷(PO4-P)的浓度分别设为200、120和100µmol/L。用1000 mL的三角烧瓶,内装 1000mL培养液(培养液由过滤消毒海水添加营养盐配制),每个三角烧瓶中装入海藻(用经蒸馏水浸泡去除硝态氮和亚硝态氮的滤纸擦干张永正等,2012)1g,加盖盖玻片,放在有光照的恒温培养箱中培养。试验前,将海藻置于试验设定的条件下先适应4 h。试验前和试验后分别测定藻体培养液中的营养盐浓度,试验时间为8 h。每个处理3个重复和1个空白对照(不加海藻,用来测定海水中的营养盐)(包杰等,2008)。
吸收速率的计算公式(Gao,1993):
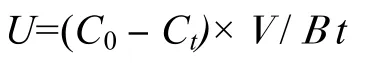
式中,U为营养盐吸收速率[µmol/(g h)];C0是空白对照的营养盐浓度(µmol/L);Ct是最终的营养盐浓度(µmol/L);V是培养液体积(L);B是海藻鲜质量(g);t是试验时间(h)。
1.2.2 藻质量对营养盐吸收的影响 2种海藻的藻体均被切割成1cm大小,藻质量设0.5、1.0、1.5、2.0、2.5g 5个梯度。试验条件同1.2.1。
1.2.3 碳源NaHCO3对营养盐吸收的影响 2种海藻的藻体均被切割成1cm大小,碳源NaHCO3浓度设0、100、400、800、1200、1600mg/L6个梯度。试验条件同1.2.1。
1.2.4 水流对营养盐吸收的影响 2种海藻的藻体均被切割成 1cm 大小,水流流速设 0、50、100、150cm/s四个梯度,水流流速通过水泵的电机功率控制,使用便携式流速测定仪测量流速。试验条件同1.2.1。
1.3 数据处理与分析
测样仪器为:Skalar San plus微量自动分析系统,NO3-N采用Cd-Cu还原后重氮偶氮法,NH4-N采用酚-次氯酸钠法,PO4-P采用抗坏血酸-磷钼蓝法(张永正等,2012)。
统计分析在 SPSS 软件包下进行。逐步回归变量筛选按概率设置阈值,P<0.05引入,P≥0.10剔除。
2 结果与分析
2.1 藻体部位对营养盐吸收的影响
把铁钉菜藻体分成顶端、中部、下部、固着器几个部分,研究其对营养盐的吸收速率。铁钉菜下部对氮磷营养盐的吸收速率最大,NO3-N、NH4-N和PO4-P的吸收速率分别为 26.0、15.8和 12.3µmol/(L·h);其次是固着器(图1)。把羊栖菜藻体分成顶端、中部、下部、固着器、气囊几个部分,研究其对营养盐的吸收速率。羊栖菜顶部对氮磷营养盐的吸收速率最大,NO3-N、NH4-N和PO4-P的吸收速率分别为26.2、15.7和 12.1µmol/(L·h);其次是中部(图2)。
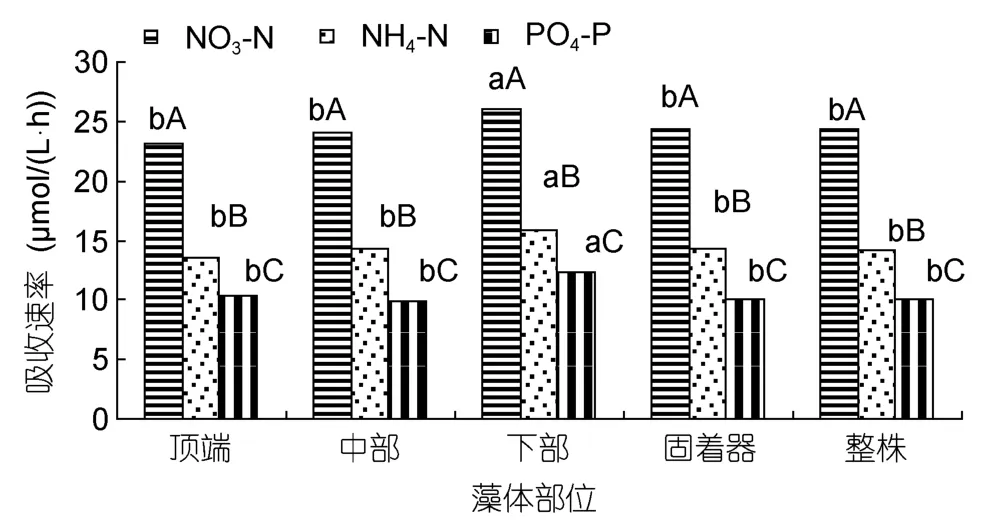
图1 铁钉菜不同藻体部位对氮磷营养盐的吸收Fig.1 The uptaking rates in different parts of Ishige foliaceaokamurai
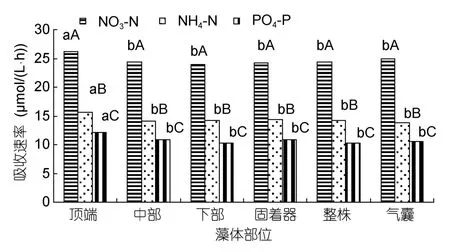
图2 羊栖菜不同藻体部位对氮磷营养盐的吸收Fig.2 The uptaking rates in different parts of Sargassum fusiforme
2.2 藻质量对营养盐吸收的影响
把铁钉菜和羊栖菜藻体分成 0.5、1.0、1.5、2.0和2.5g共5个质量梯度,研究其对营养盐的吸收。铁钉菜藻质量 1.0g对氮磷营养盐的吸收速率最大,NO3-N、NH4-N和PO4-P的吸收速率分别为27.7、16.1和12.0µmol/(L·h);其次是 1.5g;2.0和2.5g吸收速率相差不多(图3)。羊栖菜藻质量0.5g对氮磷营养盐的吸收速率最大,NO3-N、NH4-N和PO4-P的吸收速率分别为 26.8、16.9 和 13.0µmol/(L·h);其次是 1.0g;2.0和2.5g吸收速率相差不多(图4)。
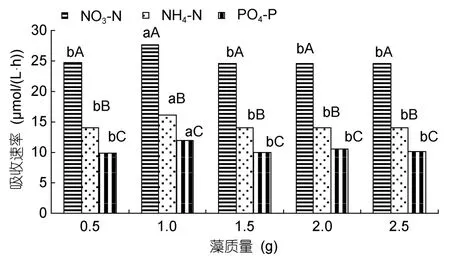
图3 铁钉菜不同藻质量对氮磷营养盐的吸收Fig.3 The uptaking rates of Ishige foliaceaokamurai in different weights
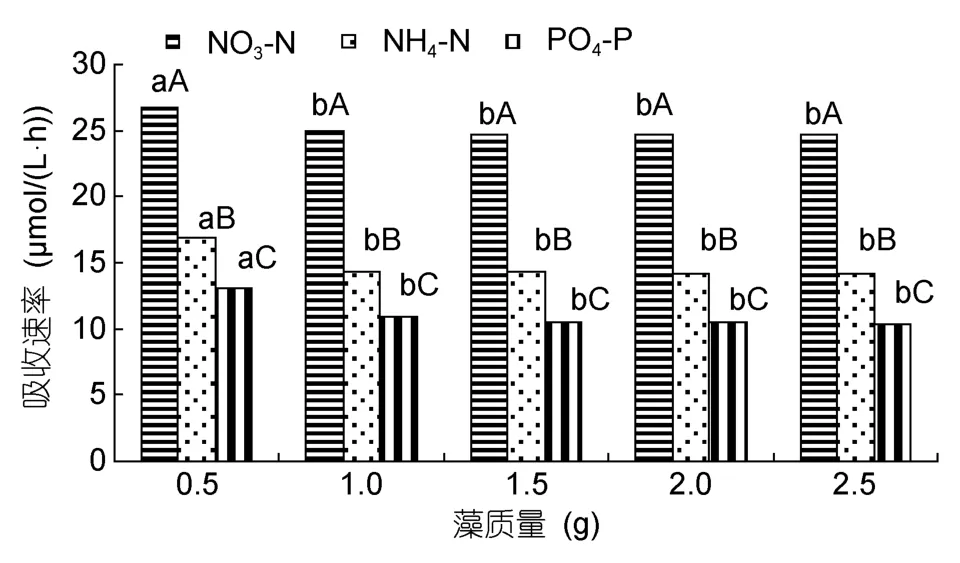
图4 羊栖菜不同藻质量对氮磷营养盐的吸收Fig.4 The uptaking rates of Sargassum fusiforme in different weights
2.3 碳源对营养盐吸收的影响
碳源质量浓度为 1200mg/L时,铁钉菜对氮磷营养盐的吸收速率最大。NO3-N、NH4-N和PO4-P的吸收速率分别为 25.9、15.3和 13.0µmol/(L·h);其次是碳源800mg/L;对照组的吸收速率相对较低,NO3-N、NH4-N和 PO4-P的吸收速率分别为 24.0、12.6和10.1µmol/(L·h)(图5)。碳源质量浓度为 800mg/L时,铁钉菜对氮磷营养盐的吸收速率最大。NO3-N、NH4-N和 PO4-P的吸收速率分别为 26.9、16.3和14.1µmol/(L·h);其次是碳源 400mg/L;对照组的吸收速率相对较低,NO3-N、NH4-N和PO4-P的吸收速率分别为 24.1、14.3 和 10.0µmol/(L·h)(图6)。

图5 碳源质量浓度对铁钉菜氮磷营养盐吸收的影响Fig.5 The uptaking rates of Ishige foliaceaokamurai under different carbon concentrations

图6 碳源质量浓度对羊栖菜氮磷营养盐吸收的影响Fig.6 The uptaking rates of Sargassum fusiforme under different carbon concentrations
2.4 水流对营养盐吸收的影响
水流对这 2种海藻对氮磷营养盐吸收的影响分别见图7和图8。铁钉菜PO4-P的吸收速率以对照组最大[12.8µmol/(L·h)],其次是 50cm/s;NO3-N、NH4-N的吸收速率以 50cm/s时最大,分别为 27.1、16.4µmol/(L·h);其次是 100cm/s(图7)。羊栖菜 PO4-P的吸收速率以对照组最大[13.3µmol/(L·h)],其次是50cm/s;NO3-N、NH4-N吸收速率在50cm/s时最大,分别为 27.9、17.3µmol/(L·h);其次是 100cm/s(图8)。
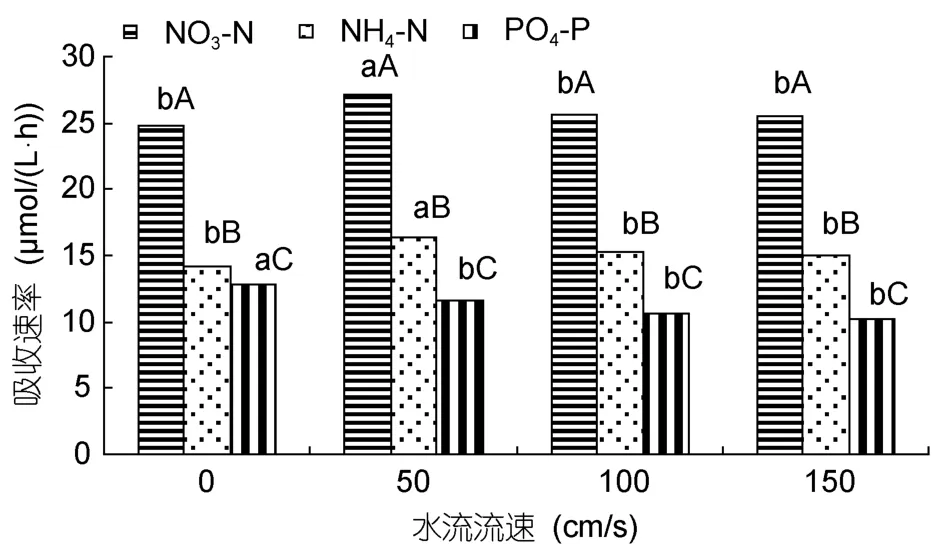
图7 不同流速对铁钉菜氮磷营养盐吸收的影响Fig.7 The uptaking rates of Ishige foliaceaokamurai under different flow speeds
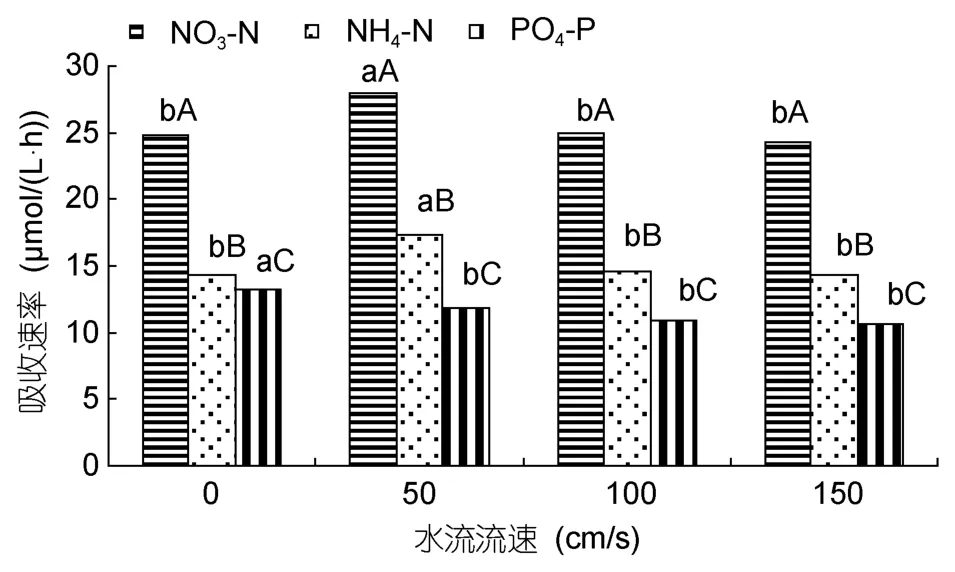
图8 不同流速对羊栖菜氮磷营养盐吸收的影响Fig.8 The uptaking rates of Sargassum fusiforme under different flow speeds
3 讨论
3.1 藻体部位对营养盐吸收的影响
试验结果表明,2种大型海藻不同部位对营养盐的吸收速率不同。铁钉菜下部对氮磷营养盐的吸收速率最大,其次是固着器。羊栖菜顶部对氮磷营养盐的吸收速率最大,其次是中部。这主要是因为铁钉菜以基部生长方式为主;羊栖菜以顶端生长方式为主(王全喜等,2008)。试验结果还可以看出,从营养盐吸收速率方面来看,羊栖菜的藻体吸收速率稍高于铁钉菜,是较好的营养盐生物过滤器。这与2种藻体的形态结构相关,铁钉菜是软骨状,体圆柱状,叉状分枝,分枝细圆柱形;羊栖菜藻体黄褐色,肥厚多汁,叶状体的变异很大。
资料显示(Flynn,1991),NO3-N和NH4-N都可以作为藻类的氮源,NH4-N的吸收明显地快于NO3-N。该研究发现,2种大型海藻对于NO3-N的吸收速率高于 NH4-N。这与某些试验结果是一致的:巨藻(Nereocystis luetkeana)试验中发现也是优先吸收NO3-N;这可能与NO3-N、NH4-N的吸收和光合作用速率受本身营养库的大小调节有关。在氮源足够的环境中,藻体细胞内的NO3-N的含量是多于NH4-N的,NO3-N较大的存储能力也会加快它的吸收速率,而过多的 NH4-N积累存储则会对藻体细胞有一定的毒害作用(包杰等,2008)。
3.2 藻质量对营养盐吸收的影响
试验中设了海藻的藻质量梯度对营养盐吸收的影响,这是因为藻质量的增加即增加了环境中大型海藻的密度,在相同的营养盐浓度环境中,大型海藻密度的增加,即增加了对环境中营养盐的吸收作用。但是该试验结果可以看出,铁钉菜藻质量 1.0g对氮磷营养盐的吸收速率最大;其次是 1.5g;2.0和 2.5g吸收速率相差不多。羊栖菜藻质量 0.5g对氮磷营养盐的吸收速率最大;其次是1.0g;2.0和2.5g吸收速率相差不多。由此可以看出,藻质量对2种大型海藻氮磷营养盐的吸收有一定的作用,每种藻体都有最适宜的添加藻质量;超过了这个藻质量范围,藻体添加再多,吸收速率也不会明显增大。这主要是因为:养殖环境容量是一定的,如果加入大量的藻体,会影响藻体进行光合作用;因为藻体的光合作用受光照的影响很大,如果加入大量的藻体,在有限的水体环境中,藻体的相互遮挡影响了它们对光的吸收,光合作用率下降,从而影响了对氮磷营养盐的吸收,所以没有体现出藻质量的影响优势(王萍等,2010)。
3.3 碳源对营养盐吸收的影响
资料显示,高 CO2浓度促进了石莼(Ulva lactuca)对营养盐的吸收,特别是NO3-N的吸收,这可能与硝酸还原酶活性有关。在高 CO2浓度下,Porphyra leucosticta的硝酸还原酶活性增高;但在高 CO2浓度生长条件下的海藻,NO3-N的吸收与同化作用并不相偶联(邹定辉等,2001)。这与笔者试验结果相一致。此试验结果表明,在碳源质量浓度为 1200mg/L时,铁钉菜对氮磷营养盐的吸收速率最大,其次是碳源800mg/L,对照组的吸收速率相对较低;在碳源质量浓度为800mg/L时,羊栖菜对氮磷营养盐的吸收速率最大,其次是碳源 400mg/L;对照组的吸收速率相对较低。
3.4 水流对营养盐吸收的影响
在低水流区域,任何溶解物质的吸收能导致岩礁周围的水位枯竭,从而产生折射边界层;水流增加导致这个折射边界层变薄或者破裂,从而增加岩礁附近可溶性营养盐和气体的数量,因此增加营养盐的吸收率(Patterson,1992;Shasharet al,1996;Thomaset al,1997)。该试验结果表明,铁钉菜和羊栖菜对氮磷营养盐的吸收,PO4-P的吸收速率对照组的最大;NO3-N、NH4-N吸收速率在 50cm/s时最大。这说明PO4-P的吸收不依赖水流的存在,静止时的吸收速率最大。而NO3-N、NH4-N的吸收需要水流,但是水流的流速不是很高,50cm/s是最适合的。这主要是因为在自然海区,铁钉菜和羊栖菜都生长在低潮带的岩礁上,有一定的水流,但是水流的流速都不大,这和它们的生长环境息息相关。过大的水流流速对铁钉菜和羊栖菜对氮磷营养盐的吸收有一定的影响。
王 萍,桂福坤,吴常文,2010.营养盐因子对孔石莼和繁枝蜈蚣藻氮、磷吸收的影响.水产科学,29(4):208—211
王 勤,宁准梅,林之川,2007.羊栖菜多糖提取工艺的研究.食品工业,(6):11—12
王全喜,张小平,2009.植物学.北京:科学出版社
毛玉泽,杨红生,王如才,2005.大型海藻在综合海水养殖系统中的生物修复作用.中国水产科学,12(1):225—231
包 杰,田相利,董双林等,2008.温度、盐度和光照强度对鼠尾藻氮、磷吸收的影响,中国水产科学,15(2):293—300
冯美霞,林 勇,刘艳如等,2011.铁钉菜乙酸乙酯提取物抗真菌活性的研究.福建师范大学学报:自然科学版,27(2):109—132
汤海峰,易杨华,姚 新等,2003.褐藻铁钉菜化学成分的研究.中国海洋药物,94(4):8—12
李 恒,李美真,徐智广等,2012.不同营养盐浓度对 3种大型红藻氮、磷吸收及其生长的影响.中国水产科学,19(3):462—470
李再亮,申玉春,谢恩义等,2011.羊栖菜对氮、磷的吸收速率研究.河南农业科学,40(3):73—77
邹定辉,夏建荣,2011.大型海藻的营养盐代谢及其与近岸海域富营养化的关系.生态学杂志,30(3):589—595
邹定辉,高坤山,阮柞禧,2001.高 CO2浓度对石莼光合作用及营养盐吸收的影响.青岛海洋大学学报,31(6):877—882
张 迪,刘艳如,郑 怡等,2009.铁钉菜抗生物质的初步提取及检测.福建师范大学学报:自然科学版,25(3):89—93
张永正,王 萍,桂福坤等,2012.石莼、铁钉菜、蜈蚣藻对氮源利用的研究.海洋环境科学,31(3):341—345
张金鼎,1998.海洋药物与效方.北京:中医古藉出版社
陈丽梅,李德茂,赵玉山等,2010.海带不同部位褐藻胶含量及其凝胶性能研究.中国食品添加剂
陈聚法,赵 俊,过 锋等,2012.条斑紫菜对胶州湾湿地浅海富营养化状况的生物修复效果.渔业科学进展,33(1):93—101
林雄平,彭 彪,陈惠斌等,2012.铁钉菜和珊瑚藻提取物抗菌活性的初步研究.宁德师范学院学报:自然科学版,24(4):352—358
徐智广,邹定辉,高坤山等,2011.不同温度、光照强度和硝氮浓度下龙须菜对无机磷吸收的影响.水产学报,35(7):1023—1029
Day J P,2003.Effect of solar irradiance and ammonium on growth,nitrogen uptake,and pigment content ofPorphyra umbilicalisKutzing,P.linearisGreville,P.leucostictaThuret in Lejolis,andP.amplissimaKjellman.MSc Thesis,University of New Hampshire,63
Flynn K J,1991.Algal carbon-nitrogen metabolism:a biochemical basis for modeling the interactions between nitrate and ammonium uptake.J Plankt Res,13(2):373—387
Gao K,Aruga Y,Asada Ket al,1993.Influence of enhanced CO2on growth and photosynthesis of the red algaeGracilariasp.andG.chilensis.J Appl Phycol,5(6):563—571
Gregory N N,Ryuta T,Tadahide N,2005.Effect of temperature and irradiance on the uptake of ammonium and nitrate byLaurencia brongniartii(Rhodophyta,Ceramiales).J Appl Phycol,17(5):371—377
Patterson M R A,1992.Chemical engineering view of cnidarian symbiosys.Am Zool,32(4):566—582
Reay D S,Nedwell D B,Priddle Jet al,1999.Temperature dependence of inorganic nitrogen uptake:reduced affinity for nitrate at suboptimal temperatures in both algae and bacteria.Applied and Environmental Microbiology,65(6):2577—2584
Sandnes J M,Kallqvist T,Wenner Det al,2005.Combined influence of light and temperature on growth rates ofNannochloropsis Oceania:linking cellular responses to large-scale biomass production.J Appl Phycol,17(6):515—525
Shashar N,Kinane S,Jokielet al,1996.Hydromechanical boundary layers over a coral reef.J Exp Mar Biol Ecol,199(1):17—28
Thomas F I M,Atkinson M J,1997.Ammonium uptake by coral reefs:effects of water velocity and surface roughness on mass transfer.Limnol Oeanogr,42(1):81—88
猜你喜欢
中国海洋大学学报(自然科学版)(2022年8期)2022-08-15 06:35:20
数学小灵通(1-2年级)(2021年4期)2021-06-09 06:26:16
数学小灵通(1-2年级)(2018年4期)2018-05-07 12:13:10
水利技术监督(2017年6期)2017-12-19 13:28:17
科技创新与应用(2017年20期)2017-07-15 20:43:02
红领巾·探索(2017年3期)2017-05-04 14:37:49
广东海洋大学学报(2015年3期)2015-12-22 10:05:28
应用海洋学学报(2015年3期)2015-11-22 07:39:10
湖泊科学(2015年4期)2015-02-17 06:43:49
应用海洋学学报(2014年2期)2014-11-26 01:20:48
