
大鼠海马结构CA1位置细胞对感觉错配适应的相关反应
2014-02-28 11:23邹丹吴敏范金戈符文双郑燕
中国医科大学学报 2014年2期
邹丹,吴敏范,金戈,符文双,郑燕
(沈阳医学院1.病理生理学教研室;2.生理学教研室;3.机能实验中心,4.2012级临床医学,沈阳110034)
大鼠海马结构CA1位置细胞对感觉错配适应的相关反应
邹丹1,吴敏范2,金戈1,符文双3,郑燕4
(沈阳医学院1.病理生理学教研室;2.生理学教研室;3.机能实验中心,4.2012级临床医学,沈阳110034)
目的观察大鼠经过学习其海马结构接受视觉-前庭-本体感觉错配格局并将其视为匹配的状态后CA1位置细胞的电活动,为揭示海马结构可编码感觉输入的任何组合提供依据。方法应用微电极细胞外记录方法,记录清醒大鼠在适应视觉-前庭-本体感觉错配条件后海马结构CA1位置细胞神经元放电情况。结果56个位置细胞中,29个神经元在正向状态中有显著的空间放电(正向相关神经元),19个神经元在反向状态中有显著的空间放电(反向相关神经元)。位置野内放电频率的分布呈现不对称及负偏斜。结论大鼠海马结构位置细胞可编码不能自然发生的新的感觉输入配置,更新海马结构比较器内的信息。
感觉错配;海马;位置细胞;神经元放电
晕动病是感觉错配引起机体功能紊乱的代表。药理学及行为学研究表明:海马结构与啮齿类动物的晕动病发生有关[1],神经不匹配信号可能产生于海马结构。我们以前研究表明[2],在感觉错配发生时海马结构的θ活动增强,反复暴露于此感觉冲突环境中5 d,θ活动逐渐降低,这表明通过学习海马结构可接受此种新的感觉配置并将其视为匹配的状态,另有文献报道:感觉错配条件下的训练可改善晕动病症状[3]。海马结构位置细胞的放电情况可反映海马结构在编码新的感觉配置时的内在活动,大鼠适应感觉错配条件后,其海马结构CA1位置细胞的电活动尚未见报道。本研究拟用清醒大鼠在视觉-前庭感觉-本体感觉错配环境适应后采用电生理方法记录CA1位置细胞放电,探讨海马结构编码新的感觉配置输入的机制,为海马结构参与晕动病的发生提供依据。
1 材料与方法
1.1 实验动物
Wistar大鼠,雄性10只,体质量200~300 g,分笼饲养,自由摄食、饮水。
1.2 方法
1.2.1 手术:戊巴比妥钠(40 mg/kg)腹腔麻醉,将牙托水泥帽安放在颅骨外表面,作为人造耳杆用于后续实验时将清醒大鼠无痛的固定于立体定位仪上。经过1周恢复,在正向条件下(视觉-前庭感觉-本体感觉正常配置)训练大鼠2周。用钻头穿过牙
托水泥帽和其下面的颅骨(AP:-2.0~-4.0,LR:2.0~6.0)钻一直径为3~5 mm的圆孔,暴露脑组织,用无菌的特氟龙软片覆盖,外涂环氧树脂胶封闭,以备神经细胞记录。
1.2.2 实验装置:将带有透明塑料罩的立体定位装置及一个小型动物跑步机安装于一个可移动转台上。大鼠的头部无痛的固定于可移动转台的立体定位架上,大鼠背部置于透明塑料罩内,四肢在跑步机上行走。在正向对照条件下,可移动转台以20 cm/s的速度沿8字型轨道(包含路径1和2)移动,大鼠在跑步机上的运动方向与可移动转台的运动方向一致。在反向条件下(即视觉-前庭感觉-本体感觉错配时),可移动转台在运动前旋转180°尽管大鼠在跑步机上向前跑,但就大鼠的运动方向而言,可移动转台是向后移动的,于是产生视觉-前庭感觉-本体感觉错配的效果。整个实验过程中跑步机运行速度与可移动转台移动速度(20 cm/s)相同。
1.2.3 神经元记录及神经元单位的隔离:整个实验包括3个时段[正向条件(正常感觉配置即对照)、反向条件(感觉错配)、正向条件(正常感觉配置即对照)]。经过连续5 d的EEG记录,EEG电功率明显下降后,从第6天开始,在海马结构CA1区记录其神经元的电活动。
玻璃隔离的钨丝微电极(Z=1.0~1.5 MΩ,1 kHz)用立体定位的方式通过脉冲电动操纵器(SM-20,Narishige,日本)逐步插入大鼠海马结构CA1区的不同部位。神经元电活动通过信号获取处理器储存在计算机中。数字化的神经元活动使用Offline Sorter程序,通过它们的波形成分隔离成独立的单位,然后将其导入NeuroExplorer程序中进一步分析。记录大鼠海马结构CA1区神经元在3个实验时段的活动情况,每个实验时段都从路径1开始。
1.2.4 位置细胞的分类:当海马结构神经元在反向时段表现出位置野,且其放电率的峰值超过对照时段相应路径的4倍,该神经元定义为相关系数r无关的反向相关神经元。当海马结构神经元表现的位置野至少出现在第1和/或第3正向时段,且该时段位置野的峰放电率是相应路径反向时段的4倍以上,该神经元叫做此路径上相关系数r无关的正向相关神经元。
如果在给定路径(路径1或2)第1和第3正向时段之间的Pearson′s相关系数r≥0.4,这种位置细胞对此路径称为经验-非依赖型正向相关神经元。如果相关系数r没有达到0.4,这些位置细胞对此路径称作经验-依赖型正向相关神经元。
1.2.5 组织学:所有实验结束时,每只大鼠用戊巴比妥钠(50 mg/kg,i.p.)再次麻醉,在神经元记录的位置处进行电凝损毁(80 μA,60 s),切片、染色确定记录位置。
1.3 统计学分析
英格曼神甫却没有生气,好像他根本没听见法比的话。法比激动起来就当不了英文的家,发音语法都糟,确实也难懂。英格曼神甫可以选择听不懂他。
2 结果
记录161个海马结构CA1神经元电活动,其中位置细胞56个。每个神经元记录均经历正向(对照)、反向(感觉错配)、再正向(对照)3种实验时段,各实验时段均包括3整圈(每圈包括路径1和2)的可移动转台的运动。海马结构神经元锋电位的叠加波形及锋电位自相关图分别显示于图1A和1B中。自相关图表明:海马结构神经元的不应期为2~3 ms,其锋电位的记录来自于单一神经元。
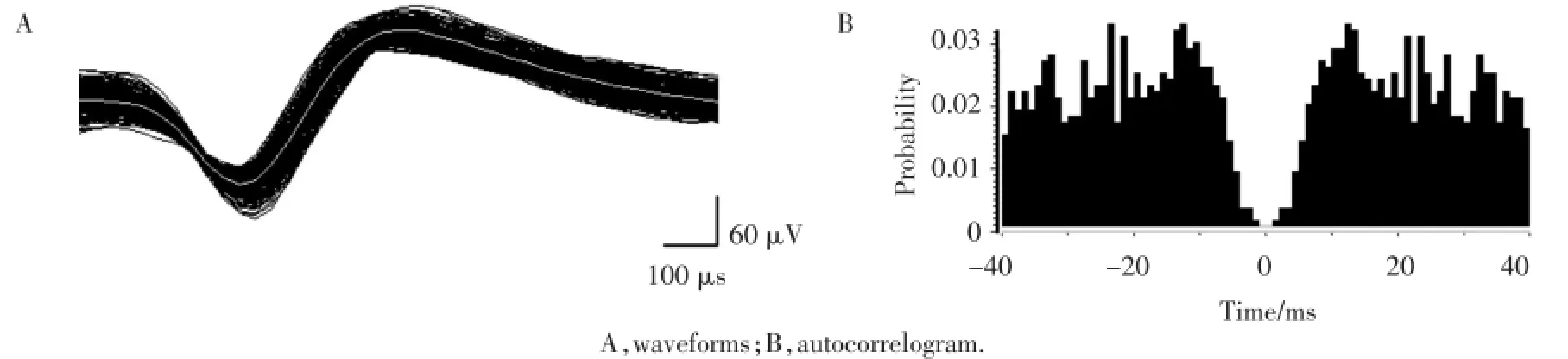
图1 海马结构CA1区用于单一神经元鉴定的细胞外记录生理参数Fig.1 Identification of a single neuron in the HF CA1 subfield on the basis of extracellularly recorded physiological parameters
2.1 正向活动相关位置细胞
在路径1,12个(21.4%)神经元显示第1及第3正向时段间有高相关系数的位置野,但在第2反向时段未显示明显位置野。在路径2,17个(30.4%)神经元显示在第1及第3正向时段存在高相关系数的位置野,但在第2反向时段未显示明显位置野。29个神经元第1与第3正向对照时段的平均相关系数(0.64±0.03)明显高于第1正向与第2反向时段的平均相关系数(-0.001±0.05)差异有统计学意义,(P<0.001,秩和检验)。
图2显示空间放电模式依赖于正向移动的海马结构位置细胞的典型位置野结构。该神经元显示第1和第3正向对照时段在路径1(r=0.76)和路径2(r=0.83)存在稳定的位置野。在第2反向时段的路径1和2上,位置野放电率的峰值低于第1正向对照时段1/4,尽管在路径1第1及第2时段存在高相关系数(r=0.58)。

图2 正向相关位置细胞Fig.2 An example of a forward⁃related place⁃differential neuron
2.2 反向活动相关位置细胞
在路径1,14个神经元(25.0%)在第1和第3正向时段没有显示有意义的位置野,但位置野出现在反向时段。在路径2,5个神经元(8.9%)出现同样的现象。图3显示位置细胞在反向时段的路径2表现出位置野,第1和第2时段的位置野间存在低相关系数[r=-0.18(路径2)]。

图3 反向相关位置细胞Fig.3 An example of a backward⁃related place⁃differential neuron
两种类型位置细胞位置野偏斜度分别是:正向相关(-0.66±0.08),反向相关(-0.50±0.07),(P>0.05,t检验)。
3 讨论
在当前的研究中,对大鼠海马结构神经元在正向(视觉-前庭-本体感觉正常配置)及反向(视觉-前庭-本体感觉错配)条件下进行电生理记录,部分位置细胞对正向及反向条件呈现特异性的空间放电形式。在此神经元记录以前,使用相同的装置大
鼠经历同样3个实验时段充分训练至少5 d,并记录其海马结构θ节律,发现反向时段θ节律的活动逐渐降低到近似第1正向时段水平(2)。这表明大鼠能很好地适应感觉错配的条件,本实验记录的神经元不是错配细胞,而是编码新的感觉输入配置的位置细胞,这些位置细胞在位置野呈现学习相关的变异特征。
位置野的分析表明:位置野是非对称性的,位置野的偏斜度是负值,放电分布的形式与以前的研究报道相似[4]。在目前的研究中,即使在感觉配置冲突的条件下(即反向相关神经元),这些非对称的改变是与运动转台的移动方向一致的。这些结果表明:海马结构能编码并预测将来的运动,即使这种运动在自然情况下不能发生。
当前的结果表明:正向相关、反向相关的位置细胞可能分别在正向及反向时段编码会聚的感觉输入配置(视觉流、前庭传入、本体感觉或运动传出“拷贝”)。Blair and Sharp观察了在前庭线索与视觉流冲突条件下丘脑背侧核头-方向细胞的活动情况[5]。海马结构位置细胞的放电活动与头-方向细胞密切相关[6]。头-方向系统的功能之一可能是给海马结构的“认知地图”定位[7]。神经生理学的研究表明:在位置导航过程中,海马结构的位置细胞对遇到的各种信息呈现相应放电配置[8]。
在多数情况下,头-方向细胞活动与前庭信息密切相关,对探测方位的神经编码关键依赖于前庭信息[9]。在当前的研究中,一些反向相关海马结构神经元在反向时段能编码冲突的感觉/运动信息,前庭与视觉的信息不能与本体感觉(运动传出“拷贝”)配。这些结果表明海马结构神经元编码会聚的感觉传入配置,这里前庭信息是感觉输入之一,但不是主要输入。海马结构可能编码感觉输入的任何组合。
在当前的研究中,一些位置细胞在重复暴露于反向条件后可编码这一条件的感觉信息。编码新的冲突环境信息的新位置细胞的形成可能有助于更新海马结构内储存的记忆,使海马比较器[10]能够接受这种新环境作为与储存信息的一种匹配。
当前的结果表明:海马结构位置细胞有能力编码一个不能自然发生的新的感觉输入配置,更新海马结构比较器内的信息。
[1]Uno A,Takeda N,Horii A,et al.Effects of amygdala or hippocampus lesion on hypergravity-induced motion sickness in rats[J].Acta Otolaryngol,2000,120(7):860-865.
[2]Zou D,Aitake M,Hori E,et al.Rat hippocampal theta rhythm during sensory mismatch[J].Hippocampus,2009,19(4):350-359.
[3]Newman MC,McCarthy GW,Glaser ST,et al.Motion sickness adaptation to Coriolis-inducing head movements in a sustained G flight simulator[J].Aviat Space Environ Med,2013,84(2):104-109.
[4]Mizuseki K,Buzsaki G.Preconfigured,skewed distribution of firing rates in the hippocampus and entorhinal cortex[J].Cell Rep,2013,4(5):1010-1021.
[5]Blair HT,Sharp PE.Visual and vestibular influences on head-direction cells in the anterior thalamus of the rat[J].Behav Neurosci,1996,110(4):643-660.
[6]Jankowski MM,Ronnqvist KC,Tsanov M,et al.The anterior thalamus provides a subcortical circuit supporting memory and spatial navigation[J].Front Syst Neurosci,2013,7,1-12.
[7]Gibson B,Butler WN,Taube JS.The head-direction signal is critical for navigation requiring a cognitive map but not for learning a spatial habit[J].Curr Biol,2013,23(16):1536-1540.
[8]Ho SA,Hori E,Kobayashi T,et al.Hippocampal place cell activity during chasing of a moving object associated with reward in rats[J]. Neuroscience,2008,157(1):254-270.
[9]Clark BJ,Taube JS.Vestibular and attractor network basis of the head direction cell signal in subcortical circuits[J].Front Neural Circuits,2012,6:7.
[10]Duncan K,Ketz N,Inati SJ,et al.Evidence for area CA1 as a match/mismatch detector:a high-resolution fMRI study of the human[J].Hippocampus,2012,22(3),389-398.
(编辑 裘孝琦)
Relative Response ofHippocampalFormation CA1 Place Cellto the Adaptation ofSensory Mismatch in Rat
ZOUDan1,WUMin-fan2,JINGe1,FU Wen-shuang3,ZHENGYan4
(1.DepartmentofPathophysiology,Shenyang MedicalCollege,Shenyang 110034,China;2.DepartmentofPhysiology,Shenyang MedicalCollege,Shenyang 110034,China;3.Center of Functional Experiment,Shenyang Medical College,Shenyang 110034,China;4.Grade 2012 Clinical Medicine,Shenyang Medical College,Shenyang 110034,China)
ObjectiveTo study the discharge change ofCA1 place cellsafterhippocampalformation(HF)accepting the visual-vestibular-proprioceptive mismatch configuration as a match by learning,which could provide new scientific experimental data for revealing the fact that HF may encode any combination ofsensory inputs.MethodsThe unitdischarge ofCA1 place cells were recorded in an awake ratby using extracellularmicroelectrode record and stereotaxic techniques afterthe ratadapted to sensory mismatch.ResultsOutof56 place cells,29 showed predominantspatial firings in the forward sessions(forward-related neuron),while 19 showed predominant spatial firings in the backward sessions(backward-related neuron).Firing rate distributions in the place fields were negatively skewed and asymmetric.ConclusionHF place cells have the ability to encode a new configuration ofsensory inputwhich doesnotoccurnaturally to update information in the HF comparator.
sensory mismatch;hippocampus;place cell;neuron discharge
R338.8
A
0258-4646(2014)02-0146-04
教育部留学归国人员科研启动基金(2013-693)
邹丹(1971-),女,副教授,博士.
E-mail:zoudan1166@hotmail.com
2013-11-18
网络出版时间:
猜你喜欢
中华耳科学杂志(2022年3期)2022-11-24
基层中医药(2022年3期)2022-07-22
中老年保健(2021年4期)2021-08-22
家教世界·V家长(2021年3期)2021-04-09
中国生殖健康(2019年8期)2019-01-07
综艺报(2018年17期)2018-09-14
现代装饰(2018年5期)2018-05-26
中国生化药物杂志(2015年4期)2015-07-07
弹箭与制导学报(2015年1期)2015-03-11
郑州大学学报(医学版)(2015年2期)2015-02-27
