
三种菜粕对罗非鱼成鱼生长性能的影响及性价比
2014-02-27 08:34:50谭芳芳高启平王若军LesNernberg
饲料工业 2014年20期
■谭芳芳 高启平 王若军 Les Nernberg
(1.通威股份有限公司,四川成都610041;2.中国农业大学神内中心,北京100193;3.加拿大双低油菜理事会,温尼伯MBR3BOT6)
由于菜籽粕中含有较多的硫代葡萄糖甙(简称硫甙)、芥酸、植酸和单宁等抗营养因子,在畜禽饲料中的使用量受到较为严格地限制,而水产动物对上述抗营养因子耐受力相对较强。在蛋白质资源紧缺的今天,菜籽粕是一种亟待开发的优良水产植物蛋白源。目前国内水产饲料中常用的菜粕有国产200型菜粕、印度菜粕和加拿大双低菜粕等。加拿大双低菜粕硫甙和芥酸含量相对最低,但价格高;国产200型菜粕和印度菜粕的毒素含量高、适口性差,但其蛋白溶解度高,价格低,因此有必要在相同条件下比较三种菜粕对鱼类的影响,探讨三种菜粕的性价比。关于菜粕对草鱼、异育银鲫、罗非鱼、斑点叉尾鮰、鲈鱼的影响已有报道,但将几种菜粕在同一试验中相同条件下进行比较的报道较少。金素雅等(2011)研究了4种菜籽饼粕对草鱼生长性能的影响,但几种菜粕在罗非鱼上进行对比未见报道。本试验以罗非鱼成鱼为对象,研究三种菜粕及其不同添加水平对罗非鱼生长、形体的影响,并在相同条件下比较三种菜粕的性价比。
1 材料与方法
1.1 试验菜粕
加拿大双低菜粕:加拿大双低菜籽在国内压榨;国产200型菜粕:国产菜籽,产地湖北沙市,国内压榨;印度菜粕:印度进口。三种菜粕的成分实测值见表1。
1.2 试验设计与饲料
试验饲料参考罗非鱼配合饲料SC/T 1025—2004的营养标准,并结合生产实际,以豆粕、棉粕为主要蛋白源,配制粗蛋白质含量为30%的无鱼粉实用配合饲料作为对照组,在此基础上,以米糠粕为填充物,分别以20%、30%用量的加拿大双低菜粕、国产200型菜粕、印度菜粕等量替代豆粕。相同水平不同种类菜粕的饲料之间,除了菜粕的种类不同,其他原料完全相同。因菜粕的赖氨酸含量较豆粕低,脂肪含量较豆粕和棉粕高,因此随着菜粕水平的增加,在饲料中补充晶体赖氨酸,降低大豆油的含量。试验饲料配方见表2,营养成分见表3。试验饲料在广东佛山通威生产线(型号南联350)上生产,制成直径3.5 mm的硬颗粒饲料。
营养成分的测定参照国家有关标准进行,水分按照GB/T 6435—2006检测,粗蛋白按照GB/T 6432—1994检测,粗灰分按照GB/T 6438—2007检测,粗脂肪按照GB/T 6433—2006检测,异硫氰酸酯按照GB/T 13087—1991检测,芥酸按照GB/T 17377—2008(2)检测,硒按照GB 5009.93—2010检测。由通威股份有限公司检测中心检测。
1.3 试验鱼
试验罗非鱼由通威原种场海南基地提供,初始平均规格285 g/尾。所有试验鱼进行10 d的适应性驯养,选择体健活泼的罗非鱼随机分组,35尾/桶,每种试验 饲料作为1个处理,共7个处理,每一处理设3个重复。
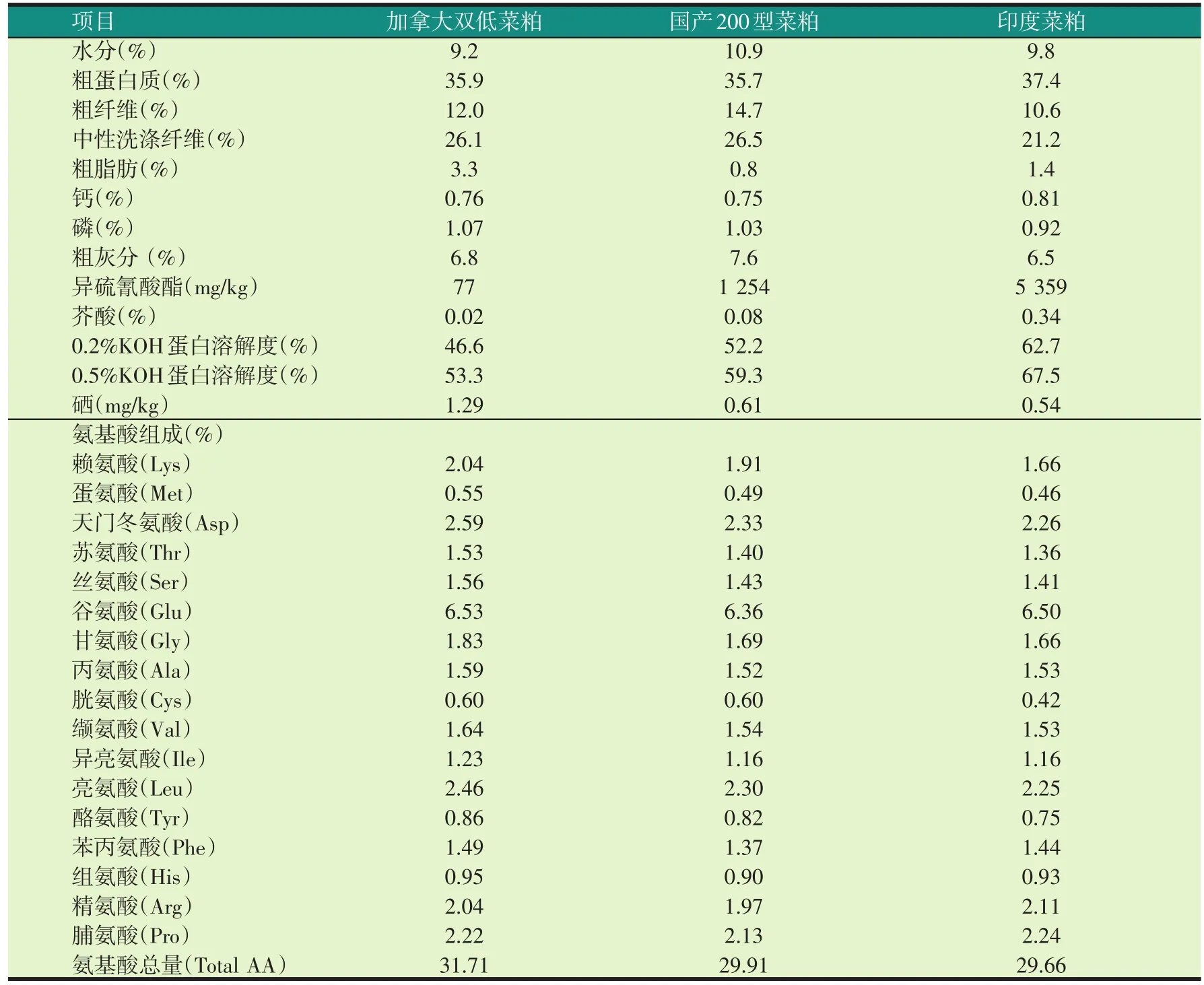
表1 三种菜粕的实测化学成分(风干基础)

表2 试验饲料配方(‰)
1.4 饲养管理
养殖试验在通威原种场海南基地室外圆柱形玻璃钢养殖桶(直径200 cm,高100 cm,容积约3.14 m3)中进行。水源为过滤的河水,空压泵24 h增氧,每两天换一次水,每天投喂3次,时间为8:00、14:00、17:00。投喂到饱食状态,根据摄食情况适当调整投饲量。养殖时间从2011年7月11日开始到10月3日结束,共84 d。试验期间水温为26.0~32.0℃,溶氧4.0~6.0 mg/l,pH值6.5~7.0,氨氮0.2~0.6 mg/l,亚硝酸盐≤0.1 mg/l,硫化物<0.05 mg/l。
1.5 观测指标
本试验从投喂饲料的第3 d开始,饲养员对各个养殖桶内的鱼的抢食状况进行打分,以评估饲料的适口性,打分从7月13日至7月27日,共15 d。评分细则如下:每天14:00投喂饲料时观察每桶鱼的抢食状况,抢食状况很好10分、较好9分、一般8分、较差7分、差6分,仅有几条鱼抢食5分,看不到鱼抢食4分。
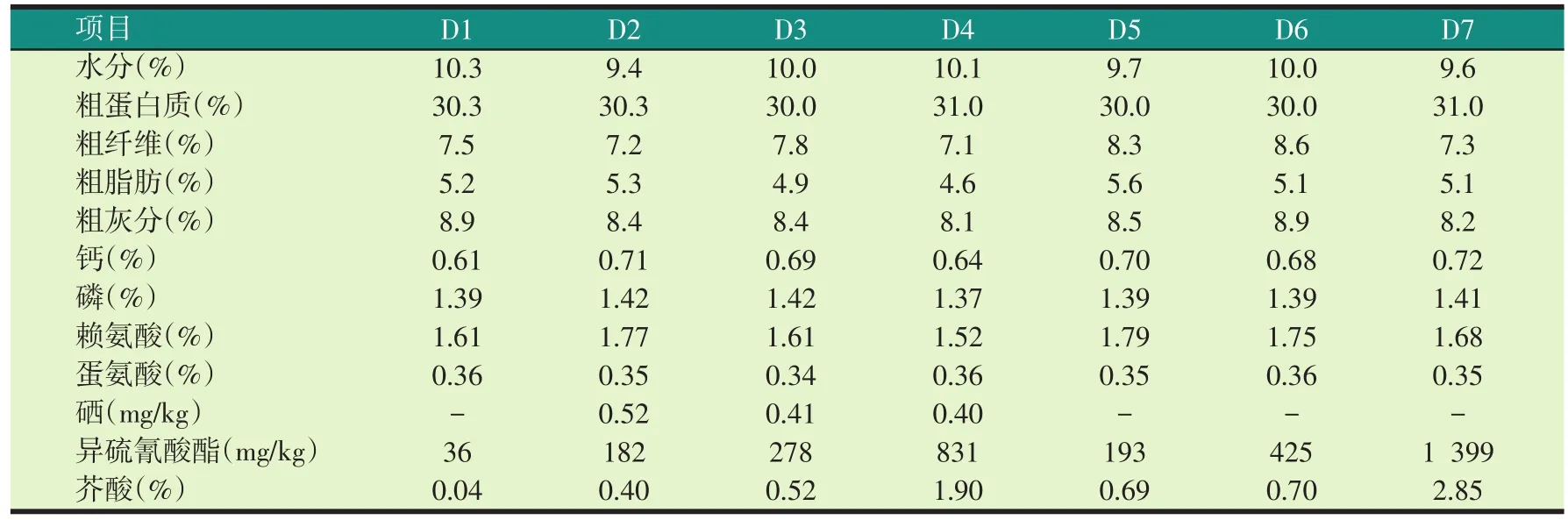
表3 试验饲料实测化学成分(风干基础)
养殖试验结束时,试验鱼饥饿1 d,每桶鱼称总重,记录尾数,计算成活率、增重率、特定生长率、饲料系数。每桶取5尾鱼测体重、体长、背肌厚,计算肥满度。解剖取出内脏团,称重,计算脏体比。分离肝胰脏称重,去皮取侧线以上背部肌。这5尾鱼的肝胰脏合并为一个样品,肌肉合并为一个样品,-20℃保存,测定异硫氰酸酯、芥酸、硒含量。
成活率(%)=100×(末尾数/初尾数);
增重率(%)=100×(末总重+死亡鱼重-初总重)/初总重;
特定生长率(%/d)=100×[Ln(末总重+死亡鱼重)-Ln初总重]/饲养天数;饲料系数=投饵量/(末总重+死亡鱼重-初总重);日摄食率(%)=100×2×投饵量/(末总重+死亡鱼重+初总重)/饲养天数;
肥满度(g/cm3)=100×体重/体长3;肝体比(%)=100×肝脏重量/鱼体重;脏体比(%)=100×内脏重量/鱼体重。
1.6 数据分析
试验数据用平均值±标准方差(Mean±S.D.,n=3)表示,结果采用Excel和SPSS13.0版统计软件中One-Way ANOVA过程进行方差分析,并进行Dun⁃can’s法多重比较,显著水平为0.05。
2 结果
2.1 试验饲料的适口性(见表4)
由表4可知,在20%水平,加拿大双低菜粕组饲料(D2)和国产200型菜粕(D3)饲料的适口性得分相当,均略高于对照组(D1),但与对照组相比无显著性差异(P>0.05)。印度菜粕组(D4)的适口性得分显著低于加拿大双低菜粕组饲料(D2)和国产200型菜粕(D3)饲料(P<0.05),也低于对照组(D1),但无显著性差异(P>0.05)。在30%水平,加拿大双低菜粕组(D5)饲料的适口性得分要略高于国产200型菜粕(D6),也高于对照组(D1),但无显著性差异(P>0.05)。印度菜粕组(D7)的适口性得分显著低于加拿大双低菜粕组(D5)饲料、国产200型菜粕(D6)组以及对照组(D1)(P<0.05)。
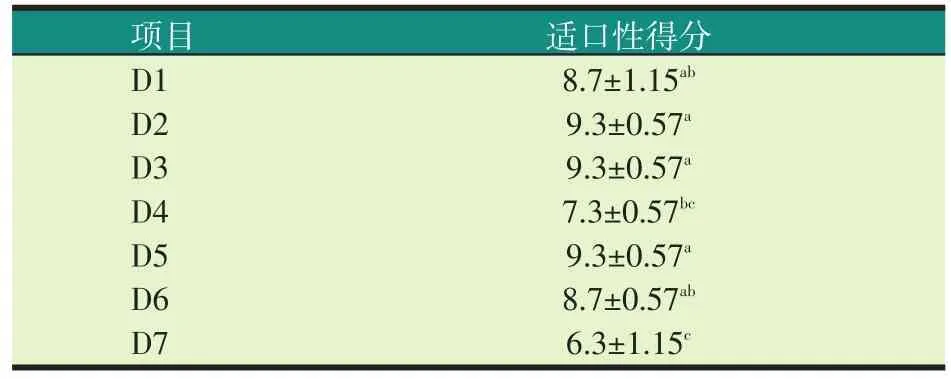
表4 试验饲料的适口性得分
加拿大双低菜粕从20%到30%水平饲料的适口性得分相当,没有下降的趋势,国产200型菜粕从20%到30%水平饲料的适口性得分略有下降,但无显著性差异(P>0.05)。印度菜粕从20%到30%水平饲料的适口性得分呈下降的趋势。
2.2 试验饲料对罗非鱼生长和饲料系数的影响(见表5)
由表5可知,罗非鱼的成活率都在97%以上,各试验组之间无显著性差异(P>0.05)。
在20%水平,饲喂20%加拿大双低菜粕(D2)和20%国产200型菜粕(D3)饲料的罗非鱼的特定生长率、饲料系数无显著性差异(P>0.05),与对照组(D1)相比也无显著性差异(P>0.05)。饲喂20%印度菜粕(D4)的罗非鱼比饲喂对照组(D1)、20%加拿大双低菜粕(D2)、20%国产200型菜粕(D3)饲料的罗非鱼饲料系数分别高6.04%、5.46%、7.22%。特定生长率比饲喂对照组(D1)、20%加拿大双低菜粕(D2)、20%国产200型菜粕(D3)的罗非鱼低9.62%、8.74%、10.48%。
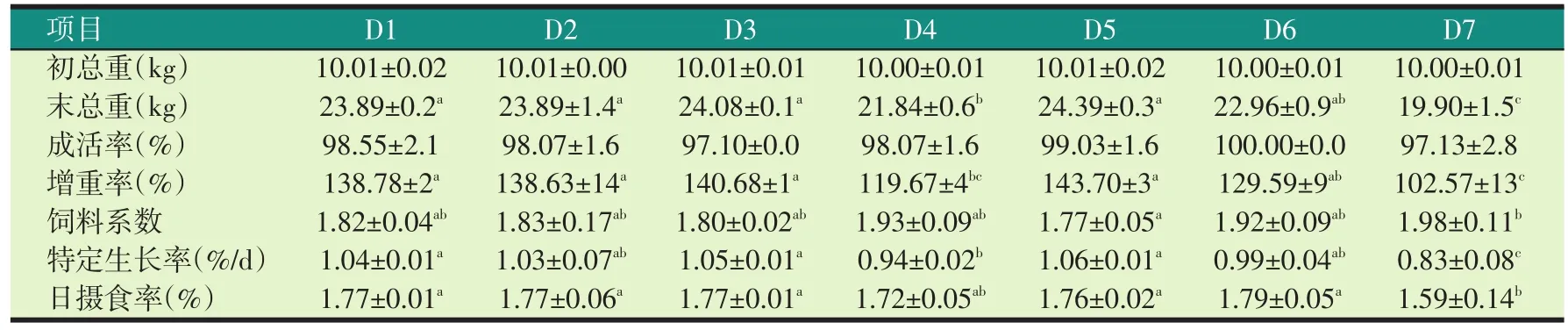
表5 试验饲料对罗非鱼生长性能的影响
在30%水平,饲喂30%加拿大双低菜粕(D5)饲料的罗非鱼特定生长率、饲料系数与对照组(D1)相比无显著性差异(P>0.05)。饲喂30%国产200型菜粕(D6)饲料的罗非鱼比对照组(D1)和30%加拿大双低菜粕(D5)饲料的罗非鱼饲料系数分别高5.49%、8.48%,特定生长率分别低4.81%、6.60%。饲喂30%印度菜粕(D7)的罗非鱼比对照组(D1)、饲喂30%加拿大双低菜粕(D5)组饲料系数分别高8.79%(P>0.05)、11.86%(P<0.05);比饲喂30%国产200型菜粕(D6)的罗非鱼饲料系数高3.13%(P>0.05)。饲喂30%印度菜粕(D7)的罗非鱼特定生长率比饲喂对照组(D1)、30%加拿大双低菜粕(D5)、30%国产200型菜粕(D6)饲料的罗非鱼低20.19%、21.70%、16.17%,差异显著(P<0.05)。
加拿大双低菜粕从20%到30%水平饲料的罗非鱼的末总重、增重率、特定生长率呈上升趋势,饲料系数呈下降趋势,国产200型菜粕和印度菜粕从20%到30%水平罗非鱼的末总重、增重率、特定生长率呈下降趋势,饲料系数呈上升趋势。
饲喂30%印度菜粕(D7)的罗非鱼日摄食率显著低于其他组(P<0.05)(除饲喂20%印度菜粕组(D4))。
2.3 试验饲料对罗非鱼形体指标的影响(见表6)
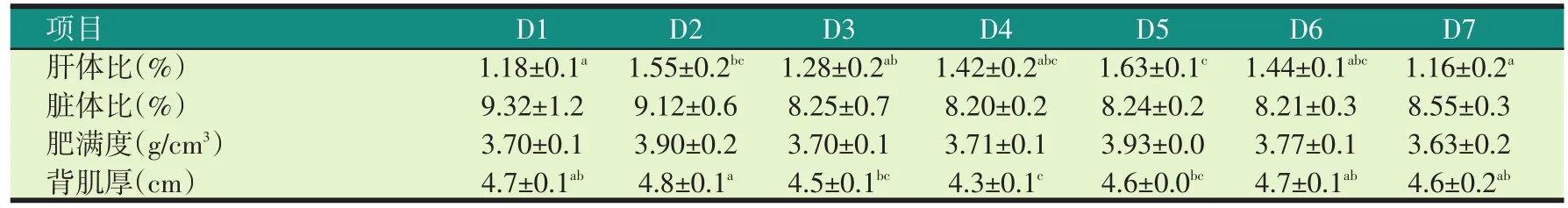
表6 试验饲料对罗非鱼形体指标的影响
由表6可知,饲喂30%加拿大双低菜粕组(D5)的罗非鱼的肝体比显著高于对照组(D1)、20%国产200型菜粕(D3)和30%印度菜粕组饲料(D7),饲喂20%加拿大双低菜粕组(D2)饲料的罗非鱼的背肌厚显著高于D3、D4、D5(P<0.05)。各试验组罗非鱼的脏体比和肥满度无显著性差异(P>0.05)。
2.4 试验饲料对罗非鱼肌肉和肝脏异硫氰酸酯、芥酸和肌肉硒含量的影响
本次试验为了探究不同菜粕对罗非鱼肌肉和肝脏毒素残留的影响,检测了肌肉和肝脏异硫氰酸酯、芥酸的含量,但肌肉和肝胰脏均未检测出异硫氰酸酯和芥酸。本试验还检测了肌肉中硒的含量,见表7。
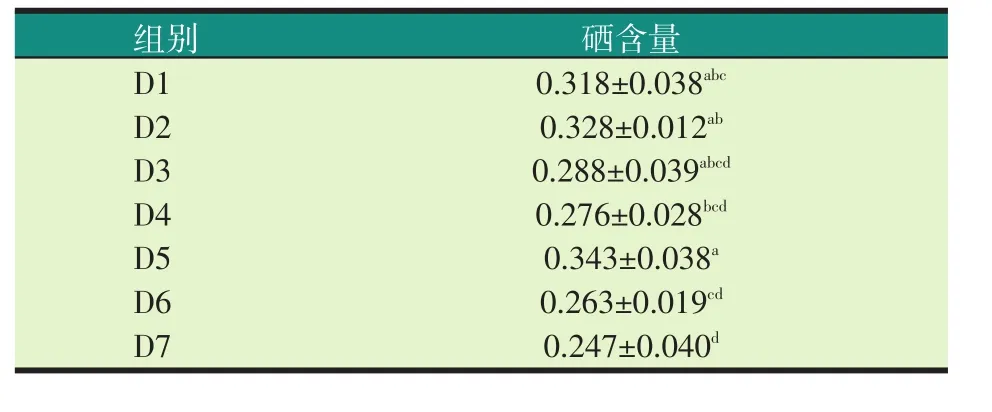
表7 各试验组罗非鱼肌肉中硒的含量(mg/kg)
由表7可知,随着加拿大菜粕水平的提高,罗非鱼肌肉中硒的含量呈上升趋势,随着国产200型菜粕和印度菜粕水平的提高,罗非鱼肌肉中硒的含量呈下降趋势。在20%水平,饲喂20%加拿大双低菜粕(D2)罗非鱼的肌肉硒含量高于20%国产200型菜粕(D3)和20%印度菜粕(D4),但差异不显著(P>0.05);在30%水平,饲喂30%加拿大双低菜粕(D5)罗非鱼的肌肉硒含量显著高于30%国产200型菜粕(D6)和30%印度菜粕(D7)(P<0.05)。
3 讨论
3.1 三种菜粕对罗非鱼生长的影响
金素雅等(2011)养殖均重为(40.4±2.4)g的草鱼60 d,结果表明:随着添加水平由23.0%增加到34.5%,国产菜籽粕组与加拿大菜籽粕组草鱼的特定生长率与饲料系数均无显著性差异(P>0.05),而印度菜籽粕组草鱼的特定生长率则显著提高(P<0.05)。本试验结果表明,随着国产200型菜粕和加拿大双低菜粕水平的增加(20%~30%),罗非鱼的增重率、饲料系数、特定生长率没有显著性差异,这与金素雅的试验结果一致。而随着印度菜粕水平的增加(20%~30%),罗非鱼的增重率、饲料转化率、特定生长率、日摄食率呈下降趋势,这与金素雅的试验结果相反。在投喂饲料时可明显观察到印度菜粕组罗非鱼的摄食不积极,随着印度菜粕水平的增加,这种现象也更加明显,这可能主要是印度菜粕苦味较重,适口性差,造成罗非鱼拒绝摄食。摄食量不足可能是饲喂印度菜粕饲料罗非鱼生长缓慢、饲料系数提高的主要原因。
马利等(2005)的研究表明,随着菜粕水平的升高,草鱼增重率、特定生长率、饲料转化率、蛋白质效率均有所下降,但在0、10.0%、20.0%和30.0%组没有显著差异。初始平均体重5.0 g的草鱼肌肉和肝脏中硫代葡萄糖甙、唑烷硫酮和异硫氰酸盐的含量随饲料中菜粕水平的升高而升高。马利所用的菜粕是国产菜粕,他的生长试验结果与本试验国产200型菜粕的结果一致。但本试验中不论是国产200型菜粕,还是加拿大双低菜粕,还是印度菜粕,在0~30%水平,罗非鱼的肌肉和肝脏中均未检测到异硫氰酸酯和芥酸。这可能因为不同种类的鱼对菜粕的耐受力也各不相同(黄辉龙等,2010)。
双低油菜籽粕粉中含有丰富的硒,本试验测得加拿大双低菜粕的硒含量为1.29 mg/kg,国产200型菜粕为0.61 mg/kg,印度菜粕为0.54 mg/kg,加拿大双低菜粕的硒含量为国产200型菜粕和印度菜粕的2倍多,许多研究表明,动物体组织中硒的含量随着饲料中硒浓度的提高而增加(黄辉龙等,2010),因此在相同菜粕水平,饲喂加拿大双低菜粕的罗非鱼肌肉硒含量高于国产200型菜粕和印度菜粕组。本试验中随着加拿大双低菜粕水平的提高,罗非鱼肌肉硒的含量增加,而国产200型菜粕和印度菜粕则随着菜粕水平的增加,罗非鱼肌肉硒的含量降低,这可能是因为硒是谷胱甘肽过氧化物酶的重要组成成分,谷胱甘肽过氧化物酶是动物体内重要的抗氧化酶,催化谷胱甘肽还原、清除机体内有毒的过氧化物,加拿大双低菜粕的硒含量高,但异硫氰酸酯和芥酸含量相对较低,用于对抗菜粕中这些毒素的硒含量越低,沉积在肌肉中的硒就越多。国产200型菜粕和印度菜粕的毒素含量较高,用于对抗菜粕中这些毒素的硒,沉积在肌肉中的硒就越少。
3.2 三种菜粕的性价比
本试验中印度菜粕用量达到20%时养殖罗非鱼的特定生长率显著低于对照组,当达到30%用量时饲料的适口性和罗非鱼的生长速度即显著下降,但是印度菜粕在价格上比其他两种菜粕低100元/t左右,因此在原料价格压力较大的情况下,可考虑使用印度菜粕,但使用量应控制在20%以内。国产200型菜粕和加拿大双低菜粕都是在国内压榨,加工工艺是相同的,菜籽品种不同,国产200型菜籽粕以国产菜籽(为黄菜籽、黑菜籽、普通菜籽、双低菜籽等的混合物)为原料(金素雅等,2011),加拿大双低菜粕以双低菜籽为原料。随着种植国产双低菜籽的推广,国产菜粕中的双低菜籽比例会越来越高,硫代葡萄糖甙和芥酸的含量也会降低,但与加拿大双低菜粕的低硫代葡萄糖甙和芥酸还有很大差距(邹新中等,2004),另外国产双低菜粕由于在种植、收购和加工环节的生物混杂和机械混杂都会导致硫苷含量的显著增加(彭健,2001),因此,加拿大双低菜粕与国产200型菜粕相比,具有硫甙和芥酸含量低,质量稳定的优势。
在低水平时(20%),加拿大双低菜粕与国产200型菜粕养殖的罗非鱼生长速度和饲料系数比较接近,但当菜粕水平增加到30%时,加拿大双低菜粕比国产200型菜粕养殖的罗非鱼生长速度快,饲料系数低。本试验在养殖效果上加拿大双低菜粕从20%增加到30%,罗非鱼的特定生长率呈上升趋势、饲料系数呈下降趋势,而国产200型菜粕从20%增加到30%,罗非鱼的特定生长率呈下降趋势、饲料系数呈上升趋势,这表明在不影响罗非鱼生长的前提下加拿大双低菜粕的使用量比国产200型菜粕可提高10%。低硫甙的双低油菜育成以后,双低菜籽粕在猪日粮中的用量得到提高(卿中全等,1999),同样在豆粕供应紧张,价格高涨时,在罗非鱼饲料中加拿大双低菜粕可更高比例地替代豆粕,降低饲料成本。
4 结论
①加拿大双低菜粕和国产200型菜粕的用量在20%~30%时对罗非鱼的生长没有显著的负面影响,两种菜粕对罗非鱼生长的影响也无显著性差异(P>0.05)。但当菜粕水平从20%增加到30%,饲喂加拿大双低菜粕罗非鱼的特定生长率呈上升趋势、饲料系数呈下降趋势,而饲喂国产200型菜粕罗非鱼的特定生长率呈下降趋势、饲料系数呈上升趋势;
②印度菜粕用量在30%时对罗非鱼的生长有负面影响(P<0.05),其用量应控制在20%以内。
猜你喜欢
CHINA TODAY(2022年4期)2022-11-22 12:43:48
花卉(2021年17期)2021-09-15 08:51:16
防护林科技(2020年9期)2020-11-09 01:45:24
中国畜牧杂志(2020年9期)2020-09-18 03:48:54
中国饲料(2020年8期)2020-05-19 02:50:26
中南林业调查规划(2018年3期)2018-11-24 03:18:48
北方牧业(2016年6期)2016-12-17 14:04:49
中南林业调查规划(2015年3期)2015-12-20 05:22:31
农产品市场周刊(2015年37期)2015-10-29 19:14:51
农产品市场周刊(2014年19期)2014-08-22 05:39:25
