
长江常熟江段似鳊的生长特征及形态特性
2014-02-15 08:01徐东坡张敏莹周彦锋赖婉婷施炜纲
大连海洋大学学报 2014年4期
徐东坡,张敏莹,周彦锋,赖婉婷,施炜纲
(1.中国水产科学研究院淡水渔业研究中心 中国水产科学研究院内陆渔业生态环境和资源重点开放实验室 农业部长江下游渔业资源环境科学观测实验站,江苏 无锡214081;2.南京农业大学 无锡渔业学院,江苏 无锡214081)
长江下游流域江河湖泊众多,水系发达,渔业水域生态环境类型丰富多样,自然条件优越,孕育了丰富的水生生物资源,然而长江下游水域重要经济渔获种类日趋单一,渔获物逐渐朝着低龄化、小型化、低质化方向演变[1]。由于过度捕捞导致主要经济鱼类种群减小,大量空间和营养生态位的空出,为小型鱼类的种群增大创造了条件。似鳊Pseudobrama simoni 属鲤形目、鲤科、鲴亚科、似鳊属,主要分布于长江、黄河和海河等水系的干支流及附属湖泊,是江湖中常见的小型鱼类。似鳊个体虽小,但由于其繁殖力强,种群数量较大,在渔业上具有一定的经济价值[2-3]。似鳊在长江下游江段[4]以及河口区潮间带[5]均为优势种类,山东、山西的许多水库中似鳊也是主要野杂鱼类中的优势种类之一,其渔获量可占水库总渔获量的20%[2]。本研究中,作者对常熟江段似鳊的年龄、生长特征和形态特征进行了研究,旨在丰富似鳊的生物学研究数据,同时为长江常熟江段的渔业资源利用和保护提供参考依据。
1 材料与方法
1.1 材料
2009年8月—12月在长江常熟江段以地笼、定置刺网等网具采集标本,从渔获物中随机选取362 尾似鳊作为样本。
1.2 样本测量及年龄鉴定
用吸水纸吸干样本体表水分,用游标卡尺测量其体长(精确到0.01 mm),用天平称量其体质量(精确到0.1 g)。用鳞片作为年龄鉴定和生长特征分析的材料,参照殷名称[6]的方法,取每尾样本背鳍起点下方、左侧线上方第3 行鳞,自前而后5 ~7 枚。鳞片用体积分数为4%的KOH 溶液浸泡12 ~24 h 后,洗去表皮和黏液,用清水漂洗后晾干,用两片载玻片夹制。用Leica M165C 体视显微镜观察年轮并照相记录,同时测量鳞径(精确到0.01 mm),所得5 ~7 枚鳞片资料取其平均值用于后续分析。年龄的分析按世代为准,经过1个生长季节,鳞片上尚未形成或正在形成年龄标志的0+龄鱼为1 龄鱼;经过2 个生长季节,鳞片上已有1个或正在形成第2 个年轮的1+龄鱼为2 龄鱼;以此类推。
1.3 数据处理
各生长指标的计算公式为
生长指标=(lg Lt+1-lg Lt)×Lt/0.4343,
年增重率=(Wt+1-Wt)/Wt×100%,
其中,Lt为t 龄时鱼的体长(mm),Wt为t 龄时鱼的体质量(g)。
体长(L)和体质量(W)的关系式用幂函数方程表示为W=aLb。体长、体质量与年龄的生长规律用Von Bertalanffy 生长方程描述:
体长生长方程为Lt=L∞[1 - e-k(t-t0)],
体质量生长方程为Wt=W∞[1 - e-k(t-t0)]b,
其中参数L∞、k和t0参照殷名称[6]描述的方法计算。回归方程的显著性采用F 检验,雌、雄生长的差异性采用残差平方和(ARSS)检验[7]。运用Excel 2010和SPSS 19.0 软件对数据进行处理。
2 结果
2.1 似鳊的种群组成
似鳊渔获物由3 个年龄组组成,1 ~3 龄鱼分别占总数的54%、29%、16%。362 尾样本中,解剖后可辨别雌、雄的334 尾,其中雌性个体156尾,雄性个体178 尾,其雌雄性比为1∶ 1.14,1龄和2 龄似鳊的雌雄性比均接近1∶ 1,但3 龄似鳊的雌雄性比为1∶ 1.68(表1)。

表1 长江常熟江段似鳊不同年龄组的生长参数Tab.1 Growth indices of Pseudobrama simoni in different age groups from Changshu section of Yangtze River
样本的体长范围为62.33 ~133.65 mm,接近正态分布,优势体长范围为75 ~110 mm,占总数的75.41%,对雌、雄个体的体长分布进行F 检验,P=0.48 >0.05,即总体方差相等,雌、雄个体体长的波动性无显著性差异,再对两组体长进行双样本等方差假设的t 检验,| t |=4.55 >t0.025(332)=1.97,P=7.61 ×10-6<0.01,表明其差异极显著;样本的体质量分布范围为3.70 ~46.10 g,呈偏态分布,优势体质量范围为3.70 ~18.70 g,占总数的80.11%,对雌、雄个体的体质量分布进行F 检验,P=0.0011<0.05,即总体方差不等,表明雌性个体体质量的波动性大于雄性个体,再对两组体质量进行双样本异方差假设的t 检验,| t|=4.64 >t0.025(293)=2.25,P=5.28 ×10-6<0.01,表明其差异极显著(图1、图2)。
2.2 似鳊的生长特征
2.2.1 体长与鳞径的关系 对似鳊雌、雄个体的体长(L)与鳞径(R)的关系分别进行拟合(图3)。结果显示,四项式方程最为合适,即
L♀=391.67R4- 2211.8R3+4609.7R2-4171.4R+1463.2,n=156,P<0.001,r2=0.6973;
L♂=171.22R4- 895.44R3+1719.8R2-1406.4R+486.47,n=178,P<0.001,r2=0.7821。
2.2.2 体长与体质量的关系 根据渔获物的实测体长和体质量,经过散点图分析,似鳊的体质量(W)和体长呈幂函数相关(图4),其回归方程为
W♀=4.52 ×10-6L3.2901,n=156,r2=0.94;
W♂=1.09 ×10-5L3.0915,n=178,r2=0.98;
W总=7.33 ×10-6L3.1818,n=362,r2=0.97。
所有方程经F 检验,P<0.005,回归均极显著。
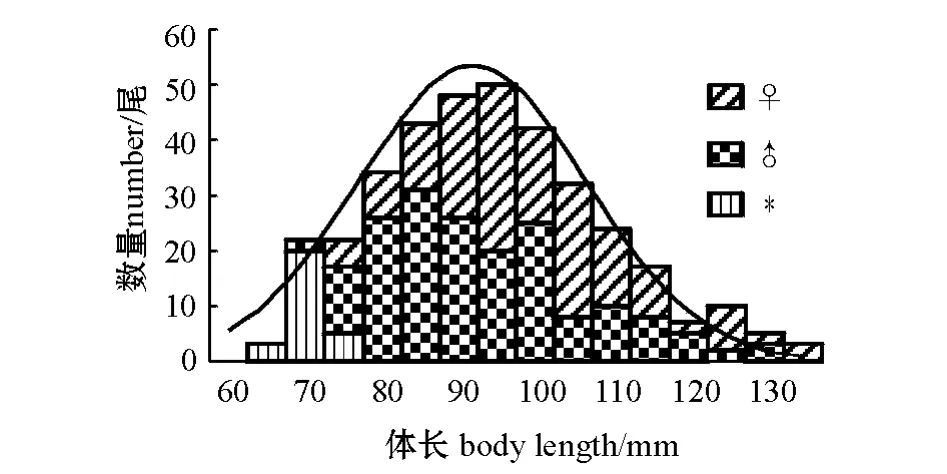
图1 长江常熟江段似鳊的体长分布Fig.1 The frequency of body length of Pseudobrama simoni in Changshu section of Yangtze River
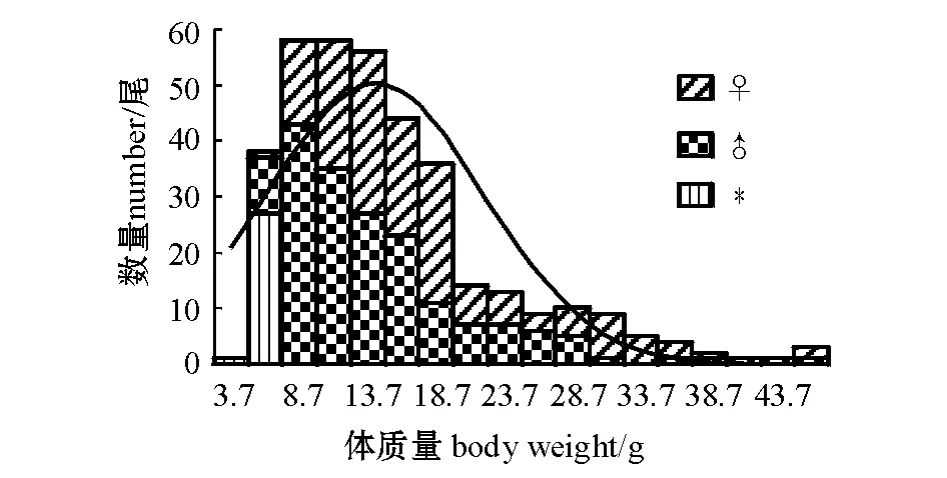
图2 长江常熟江段似鳊的体质量分布Fig.2 The frequency of body weight of Pseudobrama simoni in Changshu section of Yangtze River

图3 长江常熟江段似鳊鳞径与体长的关系Fig.3 Relationship between scale radius and body length of Pseudobrama simoni in Changshu section of Yangtze River
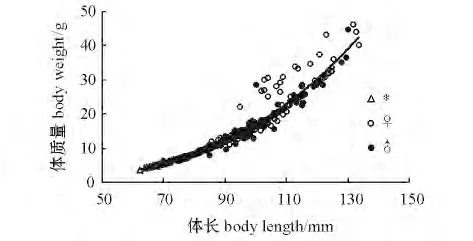
图4 长江常熟江段似鳊体质量与体长的关系Fig.4 Relationship between body weight and body length of Pseudobrama simoni in Changshu section of Yangtze River
2.2.3 生长方程 似鳊体长与体质量的关系式中b 接近3,表明似鳊的生长符合VBGF 的假设条件,因此,可用VBGF 方程描述似鳊的生长特征。根据Ford、Walford 方程和Beverton 法求得生长方程参数L∞、k、t0,似鳊的VBGF 方程为
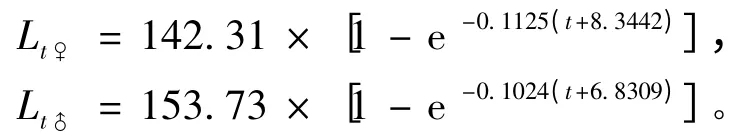
根据体质量与体长的关系式,得到体质量的生长方程为
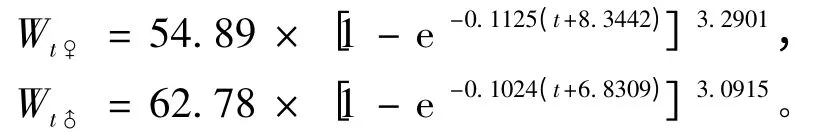
经ARSS 检验,似鳊雌、雄个体生长存在极显著性差异(F=19.73 >F(0.001,3,328)=5.55)。雄鱼比雌鱼有更大的渐近体长和渐近体质量。
为了研究似鳊生长随时间的变化特征,分别对生长方程求一阶、二阶导数,得出体长、体质量的生长速度方程和加速度方程。
(1)体长生长速度方程为

(2)体质量生长速度方程为
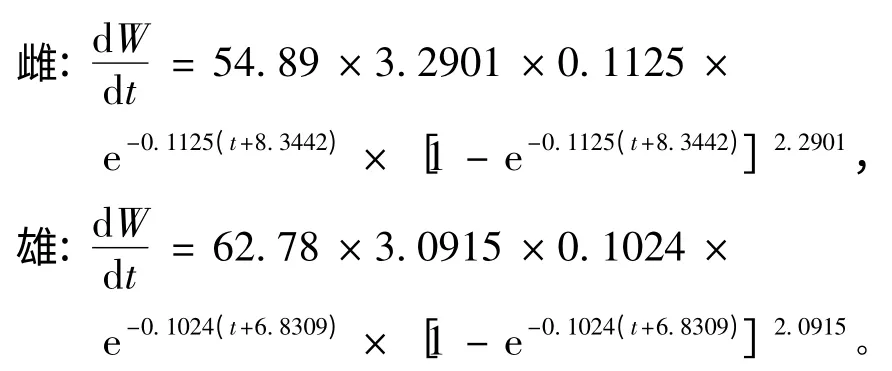
(3)体长生长加速度方程为
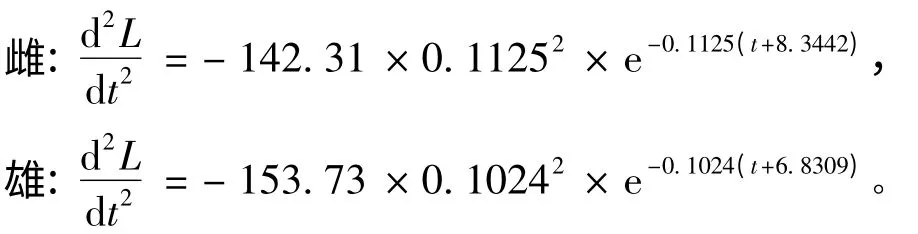
(4)体质量生长加速度方程为
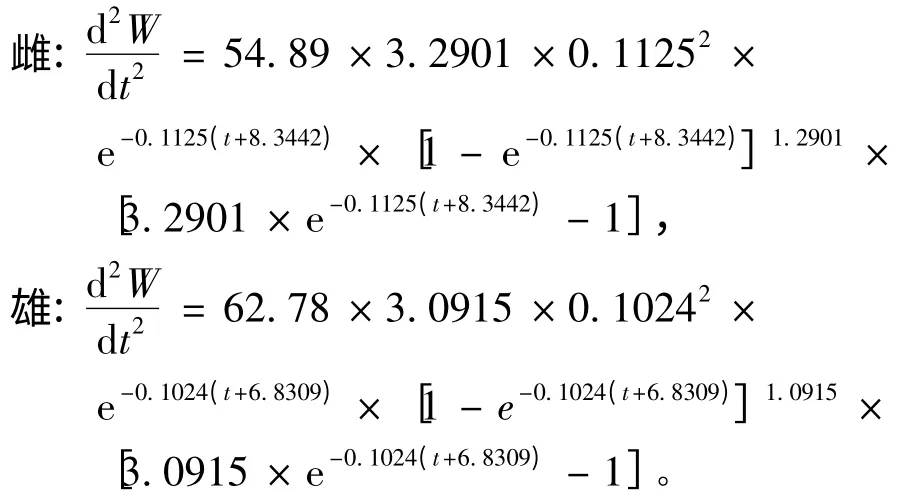
当d2W/dt2=0 时,得到体质量生长拐点年龄为2.24 龄(雌)和3.96 龄(雄),此时对应的体长分别为99.06 mm和102.82 mm,对应的体质量分别为16.66 g和18.10 g。
2.3 似鳊雌雄个体形态特征的比较
通过对61 尾似鳊(30 尾雌、31 尾雄)31 个形态指标进行比较分析,在0.1 显著性水平上,雌性和雄性似鳊各形态指标的方差均无显著性差异(P >0.05),并且各形态指标与体长均存在比较强的线性关系。表2 列出了雌性和雄性似鳊各形态指标的平均值、范围和以体长为协变量的单因素协方差分析结果,表明似鳊是无显著的两性异型。
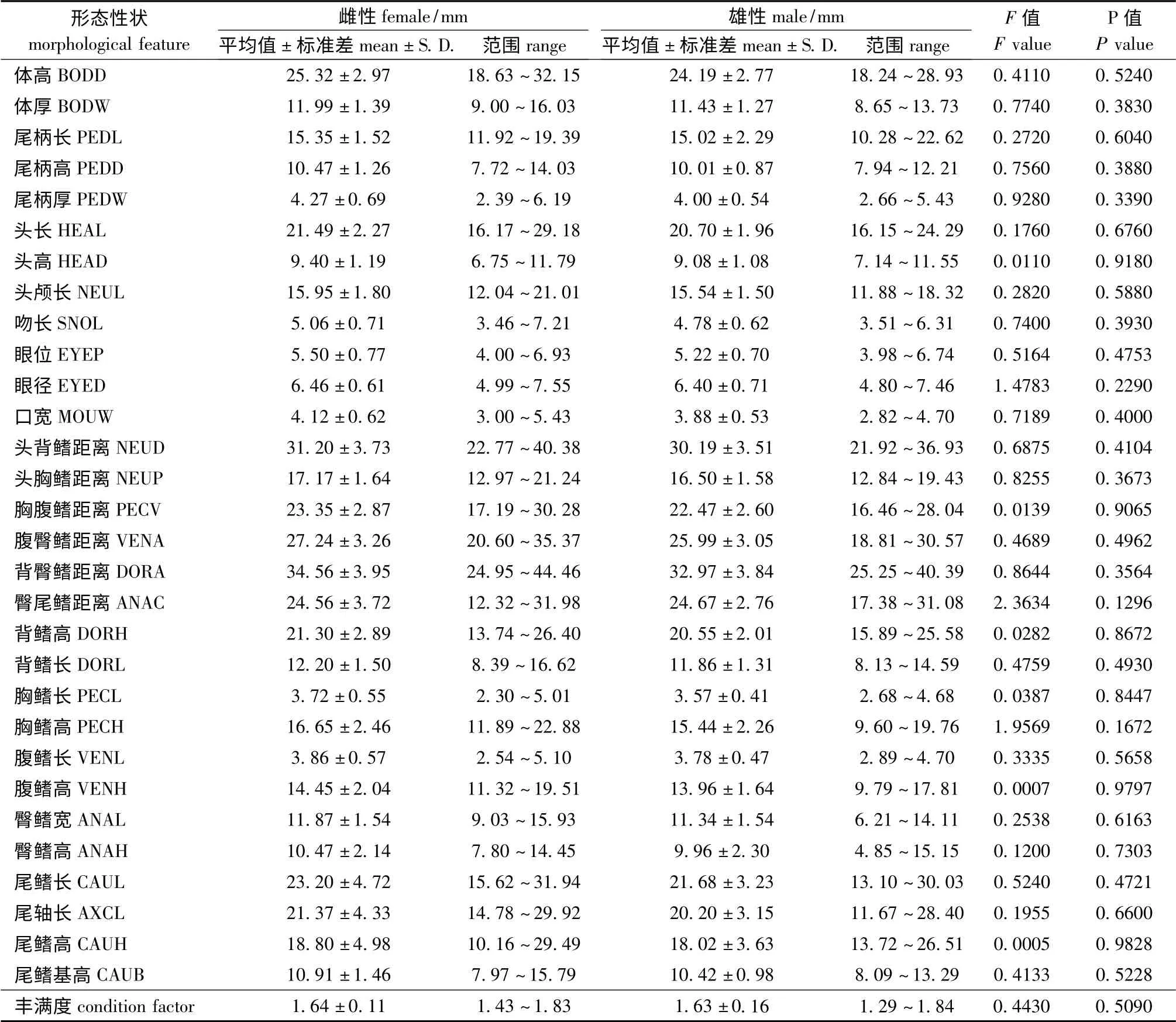
表2 似鳊雌、雄个体形态特征的比较Tab.2 Descriptive statistics and comparison of morphological feature between male and female Pseudobrama simoni
3 讨论
种群密度、饵料生物等因子的变动都会影响种群的生长[8]。似鳊为性成熟较早的鱼类,只要经过一冬龄便可达到初次性成熟,许多鱼类均通过个体大小的增长来提高繁殖力,因此,部分在生境、饵料竞争中占优势的似鳊可以获得较好的生长条件,从而尽早达到性成熟。而在竞争中处于劣势的似鳊则生长较为缓慢,这就造成了似鳊3 个年龄组中体长、体质量均有较大的变化幅度。由于饵料和竞争等原因会直接影响到幼体规格的形成[9],加之似鳊的快速生长期主要在1 龄阶段[2],因此,不同年龄组似鳊个体的体长、体质量的重叠现象均较为严重。
黄颡鱼Pseudobagrus fulvidraco 成体中,雌性个体的体长显著小于雄性个体,雌性个体的眼径、头高、体高、体质量等均大于雄性个体[10],其他一些参数则无显著性差异;花鱼骨Hemibarbus maculates雌、雄成体的体长无显著性差异,产后雌性群体的头宽、头高、尾柄长、尾柄高均大于成年雄性群体,其他参数无显著性差异[11];3 龄亚东鲑Salmon trutta fraio 雌性个体大于雄性个体[12];但海水鱼类浅色黄姑鱼Nibea coibor[13]及蓝鳍金枪鱼Thunnus thynnus[14]的体长(叉长)和体质量在雌、雄个体间无显著性差异。本研究中,不同年龄组内雌性似鳊的体长、体质量均显著大于雄性,这与李思发[3]在姚江水域的研究结果相同,这些研究结果反映了不同物种、不同地理位置、不同生境会导致生物间雌、雄差异现象的产生。除体长、体质量外,本研究中选取的其余31 个形态特征并无显著的两性差异,这与王丹婷等[15]对刀鲚Coilia nasus、徐德钦等[16]对温州厚唇鱼Cossocheilus wenchowensis的研究结果相似,尤其是背鳍、尾鳍等重要的功能性状[17]也无显著的两性差异,因此,雌、雄似鳊应未出现生态习性上的分化。
鱼类的生长是由其遗传型所决定的生长潜能与其生长过程中所遇到的各种环境条件之间相互作用的结果,同一物种不同种群的生长也随该种群的地理分布而异[6,18]。不同水域中的环境因子诸如饵料生物、水温、捕捞压力和种间竞争压力的不同,使得不同种群的鱼类生长参数会存在一定差异[19]。常熟江段似鳊的体长范围和体质量范围均与姚江群体较为接近,但远小于钱塘江群体;而与长江靖江段相比,常熟江段似鳊群体的规格略大(表3)。

表3 不同生境似鳊生长参数的比较Tab.3 Growth indices of Pseudobrama simoni in different habitats
常熟段雌、雄似鳊的k 值分别为0.11和0.10,小于天鹅洲故道似鳊(0.16和0.18)[22]及靖江段似鳊(0.17)[21];雌、雄似鳊的拐点年龄分别为2.24和3.96 龄,小于天鹅洲故道似鳊(5.80和5.22 龄),k 值和拐点年龄的差异均表明,常熟段似鳊的生长速度慢于天鹅洲故道及靖江段似鳊。本研究中,拐点年龄对应的体长、体质量值与实测结果有一定的差异,造成这一差异的原因有两方面:一个可能是本研究仅具备最少的年龄组用于计算生长方程参数L∞、k、t0,从而给VBGF 方程及拐点年龄的计算带来了误差;另一个可能是不同年龄组似鳊个体的体长、体质量的重叠现象较为严重,造成了鳞径大小与体长的相关程度较低,使得退算体长有一定的误差,进而使VBGF 方程存在一定的误差。本研究中,似鳊在1 ~2 龄处于快速生长阶段的研究结果与李德尚等[2]和李思发[3]的研究结果基本一致,同时就似鳊渔获物年龄组成而言,长江常熟段和靖江段均以1 龄似鳊为主,而其余水域则以2 龄或2 ~3 龄为主,20世纪70年代长江水域的优势年龄组也为2 龄[23],近年天鹅洲故道似鳊的优势年龄组则为1 ~3 龄,其中2 龄最多[22]。由于似鳊本身为小型鱼类,其1 冬龄即可达性成熟的特点使其容易适应较高强度的捕捞压力,而同龄的雌性似鳊规格相对大于雄性似鳊,因此,其被捕获的概率较大,这在一定程度上解释了在长期捕捞压力的选择下,雌性似鳊拐点年龄小于雄性拐点年龄的原因。
李德尚等[2]认为,似鳊1 龄阶段生长较快,之后生长缓慢的特点,以及与鳊Parabramis pekinensis和其他鲴类在食性上的重叠,使其在一定程度上浪费了水库的生产能力,目前似鳊在长江下游近岸的渔获物种占有一定的优势地位,一般被作为鳜Siniperca chuatsi、翘嘴鲌Erythroculter ilishaeformes和中华绒螯蟹Eriochier sinensis 的饵料鱼,因此,可依据似鳊繁殖力强以及1 龄期间生长速度快的特点加以开发利用。长江渔业资源在2000年之前遭到严重破坏[24],从2002 开始实行春季禁渔措施后,渔业资源衰退速度得以减缓[25],然而作为小型野杂鱼的似鳊出现低龄化的现象显示出长江渔业资源的现状仍不容乐观。
[1]施炜纲,王博,王利民.长江下游水生动物群落生物多样性变动趋势初探[J].水生生物学报,2002,26(6):654 -661.
[2]李德尚,吴雄飞,谢宗墉,等.杂鱼在养鱼水库中的作用及其种群控制[J].生态学报,1988,8(4):304 -310.
[3]李思发.关于逆鱼的生态学资料[J].动物学杂志,1981(4):6-7.
[4]陈校辉,边文冀,赵钦,等.长江江苏段鱼类种类组成和优势种研究[J].长江流域资源与环境,2007,16(5):571 -577.
[5]张衡,朱国平.长江河口潮间带鱼类群落的时空变化[J].应用生态学报,2009,20(10):2519 -2526.
[6]殷名称.鱼类生态学[M].北京:中国农业出版社,1993.
[7]Chen Y,Jacson D A,Harvey H H.A comparison of von Bertalanffy and polynomial functions in modelling fish growth data[J].Canadian Journal of Fisheries and Aquatic Sciences,1992,49:1228 -1235.
[8]张其永,李福振,杜金瑞.厦门杏林湾鲻鱼年龄和生长的研究[J].水产学报,1981,5(2):121 -131.
[9]徐钢春,顾若波,闻海波,等.澄湖似刺鳊鮈的年龄和生长特征[J].中国水产科学,2009,16(3):307 -315.
[10]林植华,雷焕宗.黄颡鱼的两性异形和雌性繁殖特征[J].动物学杂志,2004,39(6):13 -17.
[11]林植华,雷焕宗,林植云,等.花鱼骨的两性异形和雌体繁殖输出[J].上海交通大学学报:农业科学版,2005,23(3):284 -288.
[12]豪富华,陈毅峰,唐卫星,等.亚东鲑的年龄与生长的研究[J].水生生物学报,2007,31(5):731 -737.
[13]朱清澄,陈丙见,金鑫,等.缅甸安达曼海浅色黄姑鱼生物学特性的初步研究[J].大连海洋大学学报,2013,28(5):487 -491.
[14]方陈,戴小杰,朱江峰.冰岛南部海域蓝鳍金枪鱼的群体组成特征[J].大连海洋大学学报,2012,27(5):483 -488.
[15]王丹婷,杨健,姜涛,等.不同水域刀鲚形态的分析比较[J].水产学报,2012,36(1):78 -90.
[16]徐德钦,林植华,雷焕宗.温州厚唇鱼形态特征的两性异形和雌性个体生育力[J].上海交通大学学报:农业科学版,2006,24(4):335 -340.
[17]Kishida M,Kanaji Y,Xie S,et al.Ecomorphological dimorphism of juvenile Trachurus japonicus in Wakasa Bay,Japan[J].Environmental Biology of Fishes,2011,90(3):301 -315.
[18]李加儿,区又君.珠江口东岸池养鲻鱼的年龄与生长[J].热带海洋,1996,15(4):31 -37.
[19]李秀启,陈毅峰,李堃.抚仙湖外来黄颡鱼种群的年龄和生长特征[J].动物学报,2006,52(2):263 -271.
[20]陈马康,童合一,俞泰济,等.钱塘江鱼类资源[M].上海:上海科学技术文献出版社,1990.
[21]程兴华,唐文乔,郭弘艺,等.长江靖江段沿岸似鳊的时间格局及生长特征[J].上海海洋大学学报,2012,21(1):97-104.
[22]孙广文,沈建忠,胡少迪,等.长江天鹅洲故道似鳊年龄、生长和死亡率[J].淡水渔业,2013(4):29 -34.
[23]湖北省水生生物研究所鱼类研究室.长江鱼类[M].北京:科学出版社,1976.
[24]陈大庆,段辛斌,刘绍平,等.长江渔业资源变动和管理对策[J].水生生物学报,2002,26(6):685 -690.
[25]张敏莹,刘凯,徐东坡,等.春季禁渔对常熟江段渔业群落结构及物种多样性影响的初步研究[J].长江流域资源与环境,2006,15(4):442 -446.
猜你喜欢
天天爱科学(2022年9期)2022-09-15
昆明医科大学学报(2022年1期)2022-02-28
世界科学技术-中医药现代化(2021年5期)2021-11-05
中国水利(2020年23期)2020-12-24
科学导报(2020年27期)2020-05-13
现代园艺(2018年2期)2018-03-15
猪业科学(2016年1期)2016-03-30
湖南中医药大学学报(2015年1期)2016-01-06
学习月刊(2015年23期)2015-07-09
中国海洋大学学报(自然科学版)(2014年9期)2014-02-28
