
糖基化终产物对人肾小球系膜细胞氧化应激及MCP-1表达的影响及机制*
2014-01-23 03:43徐成波温俊平林桂芳黄国良
中国应用生理学杂志 2014年4期
冯 敏,徐成波,温俊平,林桂芳,吕 琦,黄国良
(1.福建医科大学附属教学医院福建省老年医院内分泌科,福州350003;2.福建中医药大学附属人民医院血液科,福州350004;3.福建医科大学附属协和医院内分泌科,福州350001)
糖尿病肾病(diabetic nephropathy,DN)是以肾小球硬化,小管间质纤维化为病理特征的糖尿病慢性并发症,其发病机制尚无定论。长期高血糖刺激下引起的蛋白质非酶糖基化,导致糖基化终末产物(advanced glycosylation end products,AGEs)形成与堆积是DN的可能发病机制之一。我们的前期研究发现,AGEs可以通过其受体(receptor for AGEs,RAGE)结合,诱导肾脏纤维化因子的表达,促进DN肾脏纤维化。国外研究表明,DN作为一种炎症性肾病,在其发生体系中,氧化应激和RAGE存在相互作用的关系,可以扩大细胞内炎症反应,加速组织器官功能损坏,加速病程的进展。然而AGEs促进这些损伤因子合成增加的具体机制尚不清楚。
本研究拟通过观察AGEs通过其受体RAGE对人肾小球系膜细胞中氧化应激(oxidative stress)及单核细胞趋化因子-1(monocyte chemoattractant protein-1,MCP-1)水平的影响,以期阐明三者之间的相互作用,为寻找DN新的治疗靶点提供理论依据。
1 材料与方法
1.1 主要试剂
牛血清白蛋白(bovine serum albumin,BSA)(美国Amresco公司);无血清低糖DMEM培养基(美国Gibco公司);胎牛血清(杭州四季青公司);D-葡萄糖(分析纯,重庆北培化学试剂厂);焦碳酸二乙酯(diethypyrocarbonate,DEPC)和 Trizol试剂(上海生物工程公司);人IgG、兔抗人RAGE多克隆中和性抗体(北京博奥森生物公司);兔抗人RAGE单克隆抗体(美国Santa Cruz公司);辣根过氧化物酶标记的抗兔二抗(北京中杉金桥生物公司);MCP-1,GAPDH引物(上海 Invitrogen公司);活性氧(reactive oxygen species,ROS)检测试剂盒(北京碧云天生物科技公司);免疫组化试剂盒(北京中杉金桥生物公司)
1.2 主要仪器
37℃饱和湿度5%CO2培养箱(中国力康公司);凝胶成像分析扫描仪(美国BIO-Rad公司);FACScan型流式细胞仪(美国Becton Dickinson公司);PCR扩增仪(美国ABI公司);核酸蛋白定量仪(美国Thermo公司);紫外分光光度仪(日本Shimadzu公司);水平式核酸电泳仪(北京六一仪器厂);低温高速离心机(美国Thermo公司)。
1.3 人肾小球系膜细胞的培养
人肾小球系膜细胞系(huamn renal mesangial cells,HRMCs)购自中国培养物保藏中心。用含10%胎牛血清的低糖DMEM培养基,于37℃、5%CO2、饱和湿度培养箱中培养,将处于对数生长期的细胞按1×105移入6孔板中,待细胞生长至80%~90%融合状态时,用无血清的低糖DMEM培养液培养24 h,使细胞生长同步化后用于实验。
1.4 AGE-BSA的制备
BSA浓度为 5.0 g/L,葡萄糖浓度为 50 mmol/L,充分溶解于pH 7.4的磷酸盐缓冲液中,过滤除菌,37℃下无菌避光孵育3个月。用磷酸盐缓冲溶液透析除去未结合的葡萄糖。同样条件制备不含葡萄糖的BSA,作为阴性对照。荧光分光光度计(激发波长370 nm,发射波长440 nm,狭缝3 nm)鉴定AGEs。
1.5 细胞干预及实验分组
分别用不同浓度的 AGE-BSA(50、100、200及400mg/L)干预培养,以不含葡萄糖的相应质量浓度的BSA为阴性对照,不含任何刺激物的低糖DMEM培养液为空白对照组,每组3个复孔,48 h后收集细胞。选取敏感浓度200 mg/L的AGE-BSA和相同质量浓度的 BSA分别培养(12、24、48及 72 h),每组 3个复孔,于不同干预时间收集细胞。抗RAGE抗体阻断实验中,将细胞分为5组:(1)不含任何刺激物的低糖 DMEM空白对照组;(2)BSA 200 mg/L组;(3)AGE-BSA 200mg/L组;(4)AGE-BSA 200mg/L+人 IgG 50mg/L组;(5)AGE-BSA 200 mg/L+兔抗人RAGE多克隆中和性抗体 50 mg/L组。每组3个复孔,48 h后收集细胞。
1.6 细胞免疫化学方法检测RAGE的蛋白表达水平
将贴壁于载玻片上的HRMCs用非免疫血清封闭后,用1∶200兔抗人RAGE抗体4℃孵育过夜后再和二抗孵育30min,加入显色剂后苏木素复染,95%乙醇脱水后封片,应用Image-Pro Plus 6.0软件分析光密度值,得出RAGE蛋白相对表达量。
1.7 ROS捕获及流式细胞仪检测
用氧化敏感探针二氯二氢荧光素-乙酰乙酸酯(2’,7’-Dichlorofluorescin diacetate,DCFH-DA)作为ROS捕获剂,以5μmol/L的DCFH-DA孵育实验组细胞30min,同时另设空白对照(不加DCFH-DA),在流式细胞仪上检测ROS水平,激发波长为488 nm,发射波长为525 nm。每个样品测定105个活细胞,检测二氯荧光黄(dichlorofluorescin,DCF)的平均荧光强度。
1.8 RT-PCR法检测MCP-1的mRNA表达水平
Trizol试剂一步法提取细胞总RNA,并鉴定RNA的完整性。取2μg RNA逆转录合成cDNA,以3-磷酸甘油醛脱氢酶(GAPDH)为内参照,上游引物5’-CAA GGTCATCCATGACAACTTTG-3’,下游引物 5’-GTCCACCACCCTGTTGCTGTAG-3’,扩增片段长度为497 bp;单核细胞趋化因子-1(MCP-1)上游引物 5’-2 TGTCTGGACCCATTCCTTCT-3’,下 游 引 物 5’-ACCAGCAAGATGATCCCAAT-3’,扩增片段长度为140 bp,扩增条件:94℃预变性 2 min,94℃变性 30 s,56℃退火30 s,72℃延伸 1 min(共 25个循环)。取PCR产物进行凝胶电泳,图像分析仪上分析目的条带与内参照条带的光密度比值进行半定量分析。
1.9 统计学处理
计量资料以均数 ±标准差(¯x±s)表示,采用SPSS 16.0统计软件进行资料分析,多组间比较采用单因素方差分析析(one-way ANOVA)。
2 结 果
2.1 不同浓度AGE-BSA对HRMCs细胞RAGE蛋白表达水平的影响
细胞免疫化学法检测显示,正常人HRMCs细胞中存在RAGE的微量表达,BSA组RAGE蛋白表达与空白对照组无明显差异,而加入不同浓度(100、200、400mg/L)的 AGE-BSA作用 48 h后,RAGE蛋白水平随AGE-BSA浓度的增高而表达增强,与空白对照组和BSA组比较有显著差异(P<0.05,图1)。

Fig.1 Expression of RAGE in HRMCs after treatmentwith different concentrations of AGE-BSA for 48 h(¯x±s,n=3)1:Blank group;2:BSA group(200 mg/L);3:AGE-BSA group(100 mg/L);4:AGE-BSA group(200 mg/L);5:AGE-BSA group(400mg/L);RAGE:Receptor for advanced glycosylation end product;HRMCs:Human renal mesangial cells;AGE-BSA:Advanced glycosylation end product-bovine serum albumin*P<0.05 vs BSA group;#P<0.05 vs blank group
2.2 不同浓度 AGE-BSA对 HRMCs细胞 ROS水平及MCP-1mRNA表达的影响
不同浓度AGE-BSA作用HRMCs细胞48 h后,AGE-BSA各组ROS产生量与空白组和相应浓度BSA组比较明显升高(P<0.05),并呈浓度依赖性上升,而BSA干预细胞后,ROS水平与空白组比较无显著差异(表1)。
RT-PCR结果显示,正常HRMCs细胞表达少量的 MCP-1。加入不同浓度(50、100、200、400 mg/L)的AGE-BSA作用48 h后,MCP-1 mRNA水平与相应浓度的BSA组和空白对照组比较均明显升高,差异有统计学意义(P<0.05),且随着AGE-BSA浓度的增加而表达增高,而相应BSA组MCP-1水平与空白组比较均无显著差异(图2)。
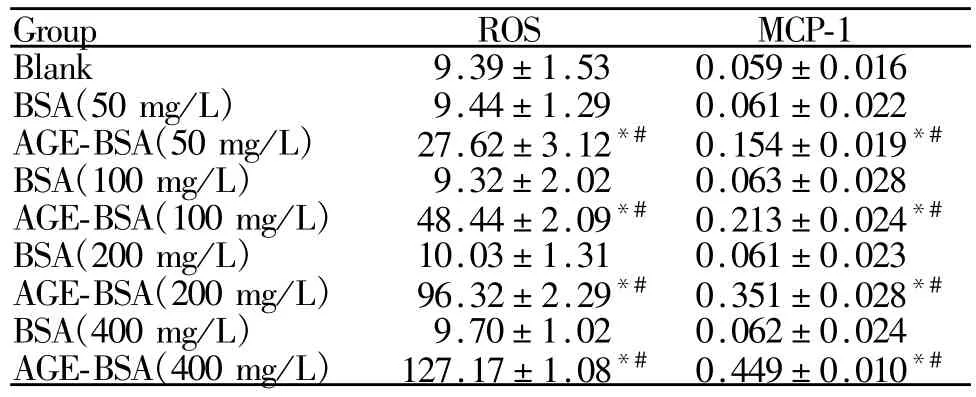
Tab.1 Product of ROS and MCP-1 in the HRMCs after treatment with different concentrations of AGE-BSA or BSA for48 h(¯x±s,n=3)
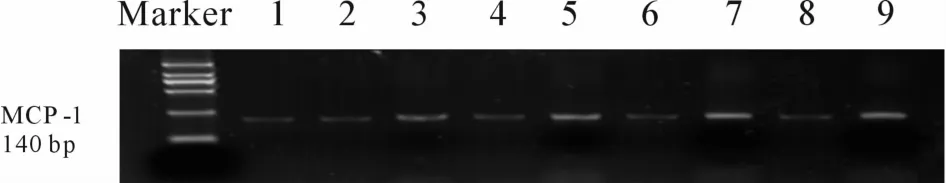
Fig.2 Expression ofMCP-1 in HRMCs after treatmentwith different concentrations of AGE-BSA or BSA for 48 h(n=3)Marker:DNAmarker(DL2000);1:Blank;2:BSA(50mg/L);3:AGE-BSA(50 mg/L);4:BSA(100 mg/L);5:AGE-BSA(100 mg/L);6:BSA(200 mg/L);7:AGE-BSA(200mg/L);8:BSA(400 mg/L);9:AGE-BSA(400 mg/L);MCP-1:Monocyte chemoattractant protein-1;HRMCs:Human renalmesangial cells;AGE-BSA:Advanced glycosylation end product-bovine serum albumin
2.3 AGE-BSA作用HRMCs不同时间后ROS水平及MCP-1mRNA表达的影响
200mg/L的AGE-BSA作用不同时间后,各作用时间点ROS产生量与空白对照组和BSA组比较明显升高(P<0.05),呈时间依赖性上升,而不同作用时间的BSA干预细胞后,ROS水平与空白组比较无显著差异(表2)。

Tab.2 Product of ROSand MCP-1 in the HRMCs after treatment with 200 mg/L AGE-BSA or BSA detected at different time points(¯x±s,n=3)
RT-PCR结果显示,加入敏感浓度200 mg/L的AGE-BSA作用不同时间(12、24、48、72h)后 MCP-1 mRNA的表达水平与200mg/LBSA相应作用时间组和空白对照组比较均显著增高,差异有统计学意义(P<0.05),且随着作用时间的延长而表达增加,而不同时间BSA组与空白组比较均无显著差异(P>0.05,图 3)。

Fig.3 Expression ofMCP-1 in HRMCs after treatmentwith AGEBSA(200 mg/L)or BSA(200 mg/L)detected at different time points(¯x±s,n=3)Maker:DNAmarker(DL2000);1:Blank;2:BSA(12 h);3:AGE-BSA(12 h);4:BSA(24 h);5:AGE-BSA(24 h);6:BSA(48 h);7:AGE-BSA(48 h);8:BSA(72 h);9:AGE-BSA(72 h);MCP-1:Monocyte chemoattractant protein-1;HRMCs:Human renal mesangial cells;AGE-BSA:Advanced glycosylation end product-bovine serum albumin
2.4 抗RAGE中和性抗体对AGE-BSA诱导HRMCs细胞内ROS水平及MCP-1mRNA表达的影响
ROS表达水平在空白组与BSA组间差异无统计学意义(P>0.05);200 mg/L的 AGE-BSA作用 48 h后,与空白组和BSA组比较ROS水平明显增加(P<0.05);AGE-BSA+抗RAGE抗体组明显抑制AGEBSA诱导ROS的产生,与AGE-BSA组比较,两组之间有显著差异(P<0.05),而 AGE-BSA+IgG组与AGE-BSA组比较无显著差异(P>0.05,表 3)。

Tab.3 Product of ROSand MCP-1 in the HRMCs after treatment with AGE-BSA or neutralizing antibodies to RAGE(¯x±s,n=3)
RT-PCR结果显示,MCP-1的表达在空白组与BSA组间差异无统计学意义;AGE-BSA组MCP-1的表达水平与空白组和BSA组比较均明显增加,差异有统计学意义(P<0.05);AGE-BSA+抗 RAGE抗体组明显抑制AGE-BSA诱导MCP-1的表达,与AGEBSA组比较,两组之间有显著差异(P<0.05),而AGE-BSA+IgG组则没有这种作用(图4)。

Fig.4 Expression of MCP-1 in the HRMCs after treatment with AGE-BSA or neutralizing antibodies to RAGE(n=3)1:Blank;2:BSA(200 mg/L);3:AGE-BSA(200 mg/L);4:AGE-BSA(200 mg/L+IgG(50 mg/L);5:AGE-BSA(200 mg/L)+ RAGE antibody(50 mg/L);MCP-1:Monocyte chemoattractant protein-1;HRMCs:Human renalmesangial cells;AGE-BSA:Advanced glycosylation end productbovine serum albumin;RAGE:Receptor for advanced glycosylation end products
3 讨 论
AGEs是由蛋白质、脂质或核酸等大分子物质在没有酶的催化下,自发地与葡萄糖或其他还原单糖反应生成的稳定共价化合物。它广泛存在人体血液及各种组织中,正常人体内AGEs水平随年龄增长可缓慢增加,但是在糖尿病状态下,持续的高糖刺激,AGEs生成增加,并与其受体RAGE结合,激发多条信号途径,诱导产生多种纤维化因子及炎症介质,导致糖尿病慢性并发症的发生。
近年来,DN被认为是一种自然免疫和低度炎症性疾病的观点日益受到关注。其中心环节可能是通过氧化应激,产生ROS,激活多条信号通路,干扰细胞胰岛素信号传导;同时又促进炎症因子的合成与分泌,形成恶性循环。其中MCP-1介导的单核细胞/巨噬细胞在肾脏的聚集和活化所引起的炎症反应是当前的研究热点之一,MCP-1过度表达会介导大量单核巨噬细胞趋化、激活,促进多种活性物质释放,以及蛋白水解酶导致肾小球结构损伤。
氧化应激是ROS生成增多或抗氧化剂清除防御作用减弱从而引起体内氧化与抗氧化的作用失衡,造成了体内大量活性氧自由基的积聚,导致活性氧连锁反应,从而促进糖尿病并发症的发生和发展[1]。在AGEs的生成过程中可伴有葡萄糖和酮胺化合物(Amadori产物)的自氧化而产生ROS,而ROS的产生也能促进AGEs的生成。ROS作为一种促炎症刺激的信号分子,介导包括RAGE在内的多种细胞膜表面受体的信号传导通路[2]。同时NADPH氧化酶作为细胞内ROS产生的主要来源,是氧化应激的基本环节[3]。AGEs与 RAGE结合后直接激活NADPH氧化酶,ROS的生成便迅速增加,并进一步激活促分裂素原活化蛋白激酶(mitogen-activated protein kinase,MAPK)、细胞外信号调节激酶 1/2(extracellular signal-regulated kinase,ERK1/2)和 P38等介导的信号通路,使核因子-κB(nuclear factor-κB,NF-κB)发生磷酸化而活化,促进肾脏细胞多种炎症因子、趋化因子及黏附因子的释放如MCP-1、白细胞介素-1、细胞黏附因子-1、转化生长因子-β、血小板源性生长因子、肿瘤坏死因子-α等,这些因子构成复杂的网络,以自分泌和旁分泌的方式作用于肾脏细胞,使肾小球发生典型的病理改变。
研究表明,在慢性肾脏病(chronic kidney disease,CKD)中,尤其是炎症性肾脏病中MCP-1的表达可显著升高[4]。DN病人尿中MCP-1的水平和蛋白尿的水平明显呈正相关[5],同时实验研究证明在特定条件的刺激下,三种肾小球固有细胞均可以表达MCP-1[6],体外实验显示高糖以及糖化血红蛋白可以直接诱导系膜细胞产生 MCP-1[7,8]。
RAGE是目前研究最明确的AGEs特异性结合受体,广泛存在于多种细胞表面,如肾小球系膜细胞、单核巨噬细胞、内皮细胞、纤维细胞和小胶质细胞等。RAGE在正常情况下表达很低,但在病理条件下如糖尿病、炎症、机械血管损伤时则可表达增加,且与配体之间存在正反馈调节机制,在炎症因子参与的病理生理网络中可能发挥着重要的介导作用,是炎症因子导致机体损伤的共同作用通道。
抗RAGE抗体和可溶性RAGE是公认的RAGE阻断剂。可溶性RAGE是RAGE的细胞外段部分,它可特异结合AGEs,但由于不具有胞内段部分,所以不介导生物效应。抗RAGE抗体则是通过封闭细胞膜上的RAGE使其不能与AGEs结合发挥生物效应。研究表明用抗RAGE抗体封闭RAGE可以减少AGEs-RAGE介导的氧化应激及炎症因子、纤维化因子的表达[9]。我们前期的实验发现AGEs可以引起肾脏纤维化因子如结缔组织生长因子(connective tissue growth factor,CTGF)、纤维连接蛋白(fibronectin,FN)的表达增加,给予抗RAGE抗体进行干预后,可以完全阻断AGEs的诱导作用,无关抗体则不能阻断AGEs使 CTGF、FN表达升高的效应,提示 AGEs诱导肾小球系膜细胞中纤维化因子的表达是由RAGE介导的。其机制可能是,AGEs与RAGE结合通过TGF-β依赖性途径诱导CTGF表达增加,并通过蛋白激酶C(protein kinase C,PKC)通路增加 FN的表达[10]。
本实验研究发现,在AGE-BSA刺激下,RAGE蛋白水平随AGE-BSA浓度的增高而表达上调,并明显促进肾小球系膜细胞中ROS的水平,呈时间和剂量依赖性。与此同时,随着浓度的增加和时间的延长,MCP-1表达也逐渐增强,通过中和抗RAGE抗体封闭细胞膜上RAGE,阻止AGEs与受体结合,则可抑制这些作用,表明 AGEs的这一生物学作用是由RAGE介导。AGEs以RAGE依赖方式诱导肾小球系膜细胞内ROS快速地产生,并直接诱导系膜细胞表达MCP-1,且这种作用可以被抗RAGE抗体所抑制,证明ROS作为一种信号分子,可以直接诱导细胞表达MCP-1。因此ROS在AGEs诱导的信号传导途径中具有中心地位。
总之,本研究证实AGEs与细胞表面受体RAGE相互作用可激活氧化应激并诱导肾小球系膜细胞中MCP-1的表达增强,是糖尿病炎症损伤过程中的重要环节。在糖尿病及其慢性并发症的发生机制和防治研究中,RAGE可能作为一个有潜力的干预靶点,通过抑制氧化应激或阻断AGEs-RAGE的相互作用,从而减少ACES的生成,为糖尿病及其并发症防治提供新思路、新方法。
[1] 张利华,张 薇,韦广洪,等.α-亚麻酸对糖尿病大鼠炎症介质及氧化应激的影响[J].中国应用生理学杂志,2012,28(1):64-67.
[2] Jiang Z,Seo JY,Ha H,etal.Reactive oxygen speciesmediate TGF-beta1-induced plasminogen activator inhibitor-1 upregulation inmesangial cells[J].Biochem Biophys Res Commun,2003,309(4):961-966.
[3] 钟林娜,黄国良,冯 敏,等.缬沙坦对人肾小球系膜细胞糖基化终产物受体表达的影响[J].中国应用生理学杂志,2011,27(3):338-342.
[4] ProdjosudjadiW,Gerritsma JS,van Es LA,etal.Monocyte chenoattractant protein-1 in normal and diseased human kidneys:an immunohistochemical analysis[J].Clin Nephrol,1995,44(3):148-155.
[5] Banba N,Nakamura T,Matsumura M,et al.Possible relationship ofmonocyte chem oattractant protein-1 with diabetic nephropathy[J].Kidney Int,2000,58(2):684-690.
[6] Segerer S, Nelson PJ, Schl ndorff D.Chemokines,chemokine receptors,and renal disease:from basic science to pathophysiologic and therapeutic studies[J].J Am Soc Nephrol,2000,11(1):152-176.
[7] Ha H,Yu MR,Choi YJ,et al.Role of high glucose-induced nuclear factor-kappaB activation inmonocyte chemoattractant protein-1 expression by mesangial cells[J].J Am Soc Nephrol,2002,13(4):894-902.
[8] Yamagishi S,InagakiY,Okamoto T,etal.Advanced glycosylation end product-inducted apoptosis and overexpression of vascular endothelial growth factor and monocyte chemoattractant protein-1 in human cultured mesangial cells[J].J Biol Chem,2002,277(23):20309-20315.
[9] Basta G,LazzeriniG,Massaro M,etal.Advanced glycation end products activate endothelium through signal-transduction receptor RAGE:amechanism for amplification of inflammatory responses[J].Circulation,2002,105(7):816-822.
[10] Twigg SM,Cao Z,MCLenna SV,et al.Renal connective tissue growth factor induction in experimental diabetes is prevented by aminoguanidine[J].Endocrinol,2002,143(12):4907-4915.
猜你喜欢
世界科学技术-中医药现代化(2021年8期)2021-12-21
天津医科大学学报(2021年4期)2021-08-21
世界科学技术-中医药现代化(2020年2期)2020-07-25
中成药(2018年6期)2018-07-11
中成药(2018年5期)2018-06-06
中成药(2017年12期)2018-01-19
中成药(2017年8期)2017-11-22
医学研究杂志(2015年9期)2015-07-01
癌变·畸变·突变(2015年3期)2015-02-27
老友(2010年9期)2010-09-16
