
青年猫与老年猫初级听皮层GABAA受体免疫表达的比较研究
2014-01-17 12:21王顺昌
中国组织化学与细胞化学杂志 2014年1期
罗 勋 王 云 王顺昌
(淮南师范学院生命科学系 安徽232001)
研究发现,在年龄超过65岁的老年人中,老年性聋患者达到35%以上,年龄在75岁以上的老年人中,老年性聋患者达到45%以上[1]。目前,年龄相关性听觉机能衰退的生理机制还尚不清楚,一些研究者提出可能是由于听觉通路中下丘(inferior colliculus,IC)及 内 侧 膝 状 体 (medial geniculate body,MGB)结构的变化,如神经元突触密度下降、丢失;以及生理功能改变,如谷氨酸脱羧酶(GAD)的变化导致GABA及GABAA受体含量的减少,进而导致抑制系统功能减弱[2-4]。亦有研究者认为主要是由于皮质中枢增龄性变化所伴随出现的对声音信号时间分析速度下降,而非皮质以下中枢(下丘和内膝体)[5]的结构和功能的增龄性变化所引起。
γ-氨 基 丁 酸 (gamma-aminobutyric acid,GABA)是神经系统重要的抑制性递质,广泛分布于各听觉脑区,发挥抑制作用,其抑制效应主要由GABAA和GABAB两种受体亚型介导。已有研究显示,在脊椎动物中,GABAA受体较GABAB受体分布更为广泛,且GABAA受体和皮层中枢听觉功能关系较为密切[6]。针对初级听皮层,已有大量报道,主要涵盖输入输出信号,细胞结构,选择性反应。选择性反应的不同,部分原因是由跨不同脑区抑制性环路所引起,例如,在初级听皮层(A1)和次级听皮层(AII)相互通联的神经元解剖结构不同,可能导致其具有特殊的整合性能[7],且神经元呈现出感受野的年龄性变化[8]。
我们已经报道了青年猫与老年猫初级听皮层GABA呈增龄性变化[9],但作为听觉高级中枢的初级听皮层GABAA受体的增龄性变化尚未见报道。本实验旨在通过免疫组织化学与免疫印迹相结合的方法,探讨初级听皮层神经元GABAA受体是否存在年龄相关变化,以期为探讨听觉机能衰退的生理原因提供有价值的形态学数据。
材料和方法
1.材料及组织准备
取青年猫(2龄,3-3.5kg)和老年猫(10-13龄,3-3.5kg)各5只。盐酸氯胺酮(40mg/kg体重)麻醉,开胸后暴露心脏,从心尖处将导管插入左心室,直至升主动脉,将0.9%的生理盐水注入导管,以便洗去血管内血液,至肝脏发白后接着将4%多聚甲醛溶液(200ml/kg体重)注入导管,以便固定液能够通过血管输送至全身组织实行预固定,随后开颅取脑(见图1),切取相应的初级听皮层(AI),放入戊二醛中固定24h,使用PBS(0.1mol/L,pH7.4)洗三次后,行常规脱水、透明、包埋,作厚5μm切片,裱片于贴有粘片剂(APES)的载玻片上,待用于免疫组织化学染色和阴性对照实验[5]。
图1 猫的全脑解剖图,箭头所示方框为初级听皮层组织Fig.1Picture of the cat's whole brain anatomy,Arrow box as shown in the primary auditory cortex
取青年猫和老年猫初级听皮层组织于裂解液A(50mmol/L pH7.4三羟甲基氨基甲烷盐酸缓冲液,150mmol/L 氯化钠,2mmol/L 乙二胺四乙酸,20mg/ml二硫苏糖醇,0.5g/ml氟化钠,17.5mg/ml苯甲基磺酰氟,5%叠氮化钠)中,浸泡约10min,用解剖刀切成碎块,将组织碎片移入二倍体积的裂解液B(50mmol/L pH7.4三羟甲基氨基甲烷盐酸缓冲液,10%(v/v)丙三醇,0.5mmol/L乙二胺四乙酸,1mg/ml亮肽素,1mg/ml抑肽素,15.5mg/ml二硫苏糖醇,0.5g/ml氟化钠,17.5mg/ml苯甲基磺酰氟,5%叠氮化钠),电动匀浆10000转3min,浸泡60min。超声振荡5min,低温离心10000转/min,15min,取上清液-20℃保存,待用。
2.免疫组织化学ABC染色法[5]
将3组已选石蜡切片作GABAA抗体和阴性对照染色。先脱蜡至水,后3%双氧水室温25°C孵育15min;使用去离子水冲洗后,再用磷酸缓冲液(PBS)浸泡3min;用山羊血清孵育10min;用吸水纸吸去血清,随后用兔抗人GABAA多克隆抗体(稀释比1∶2000),于37℃孵育2h;接下来用PBS洗后用生物素标记羊抗兔IgG孵育15min;再用PBS洗后;使用链酶卵白素过氧化物酶(ABC)孵育15min,最后用PBS洗后;直接用3,3-二氨基联苯胺(3,3NDiaminobenzidine Tertrahydrochloride,DAB)显色,去离子水冲洗,脱水,透明,封片。采用正常山羊血清代替一抗作阴性对照实验。
3.Western Blot
取青年猫和老年猫A1区总蛋白于15%十二烷基硫酸钠-聚丙烯酰胺凝胶电泳后,湿转入聚偏氟乙烯(PVDF)膜。在分子量70kD处将PVDF膜分为二部分,根据分子量上半部分是GABAAR,下半部分是内参Beta肌动蛋白(β-actin)。上部分PVDF膜分别经①5%脱脂奶粉封闭1h;②4%牛血清白蛋白(BSA)封闭15min;③兔抗 GABAAR (1∶2000)4°C,12h;④辣根过氧化物酶标记的羊抗兔IgG,室温孵育1h;下半部分将β-actin (1∶2000)代替GABAAR抗体,其余步骤同上;增强化学发光法(ECL)显色曝光,10s后冲洗胶片[10]。
4.数据统计和图像处理分析
实验处理后的切片用Olympus BX-51型显微镜观察拍照。灰质部 GABAAR-IR (GABAAR-immunoreaction)神经元测量方法相同(计数细胞以细胞核可见为准)。辨别及神经元数量统计,随机选取10张切片,采用Image-Pro Express 5.1图像分析软件测量初级听皮层内GABAAR-IR神经元密度,取平均值。测量视野内GABAAR阳性反应灰度,取平均值。Western Blot结果亦采用Image-Pro Express 5.1图像分析软件测量其灰度。对所得数据均采用平均值±标准差±s)表示,t检验作差异显著性分析。
结 果
1.免疫组织化学阳性反应结果
GABAAR-IR细胞显微镜下呈深棕黄色,清晰易辨。分布于初级听皮层各层,尤其是Ⅲ、Ⅳ、Ⅵ较为密集,胞浆呈阳性反应,胞核不着色,胞体大小不一(图2A、B)。老年猫 AI区的GABAAR-IR细胞分布密度显著降低,尤其是Ⅲ、Ⅳ、Ⅵ层,阳性反应亦明显减弱(图2C、D)。统计分析发现老年猫初级听皮层GABAAR-IR阳性细胞密度比青年猫的减小了29.19%,其光密度值增加了20.7%(Image-Pro Express 5.1图像分析软件所测量的灰度值与阳性反应强弱是反向关系)具显著差异(P<0.05)(表1);青年猫阳性反应细胞密度113.28占神经元总密度605.05[9]的18.72%,老年猫阳性反应细胞密度80.21占神经元总密度598.67[9]的13.4%,减少了5.32%。阴性对照组未见阳性反应。
表1 AI区GABAA-IR神经元密度(细胞数/mm2)及GABAAR免疫阳性反应±s)Table 1 Density(number of cells/mm2)of and gray values of GABAA-IR in AI area±s)

表1 AI区GABAA-IR神经元密度(细胞数/mm2)及GABAAR免疫阳性反应±s)Table 1 Density(number of cells/mm2)of and gray values of GABAA-IR in AI area±s)
*表示P<0.05(t-test).
灰质GABAAR-IR神经元密度Density of GABAAR-IR in gray matter(/mm2)免疫阳性反应灰度值Average gray values of GABAA-IR青年猫Young cat(n=5)113.28±7.65 105.38±5.25老年猫Old cat(n=5) 80.21±5.77*127.19±6.35*
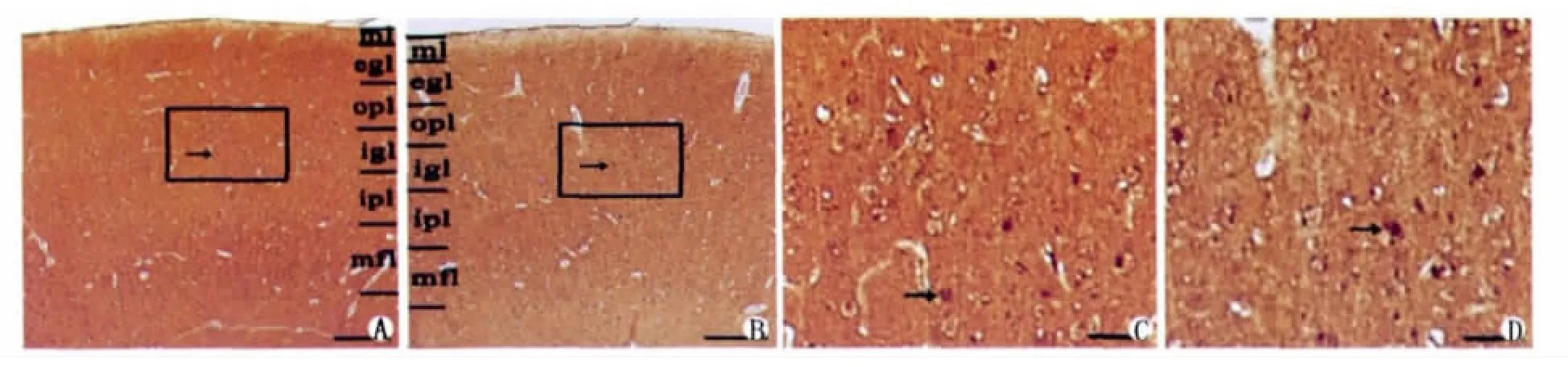
图2 A.示青年猫初级听皮层免疫组织化学染色(A1)分层:分子层(ml)、外颗粒层(ogl)、外锥体层(opl)、内颗粒层(igl)、内锥体层(ipl)、多形层(mfl)。图中方框部分为图3的截图,标尺=300μm;B.示老年猫初级听皮层免疫组织化学染色(A1)分层:分子层(ml)、外颗粒层(ogl)、外锥体层(opl)、内颗粒层(igl)、内锥体层(ipl)、多形层(mfl)。图中方框部分为图4的截图,标尺=300μm;C.示青年猫初级听皮层灰质(opl)、(igl)层的GABAAR-IR细胞(→),标尺=100μm;D.示老年猫初级听皮层灰质(opl)、(igl)层的 GABAAR-IR细胞(→),标尺=100μm 。Fig.2A.Showing all layers stained by immunohistochemical method in A1of young cat:molecular lamina(ml),external granular lamina(egl),external pyramidal lamina(opl),internal granular lamina(igl),internal pyramidal lamina(ipl),multiform layer(mfl).GABAA-IR cells(→).Scale=300μm;B.Showing all layers stained by immunohistochemical method in A1of old cat:molecular lamina(ml),external granular lamina(egl),external pyramidal lamina(opl),internal granular lamina(igl),internal pyramidal lamina(ipl),multiform layer(mfl).GABAA-IR cells(→).Scale=300μm;C.Showing GABAAR-immunoreactive cells of layers(opl),(igl)in gray matter of primary auditory cortex in young cat.GABAAR-IR cells(→).Scale=100μm;D.Showing GABAAR-immunoreactive cells of layer(opl),(igl)in gray matter of primary auditory cortex in old cat.GABAAR-IR cells(→).Scale=100μm
2.Western Blot结果
从图3A可以明显看出,老年猫初级听皮层GABAAR表达量明显少于青年猫;Image-Pro Express 5.1图像分析软件所测出青年猫的灰度值是89.57,老年猫的灰度值是110.32,因软件灰度值设置与表达强度的反向关系,结果老年猫的蛋白表达量相对减少了23.16%。
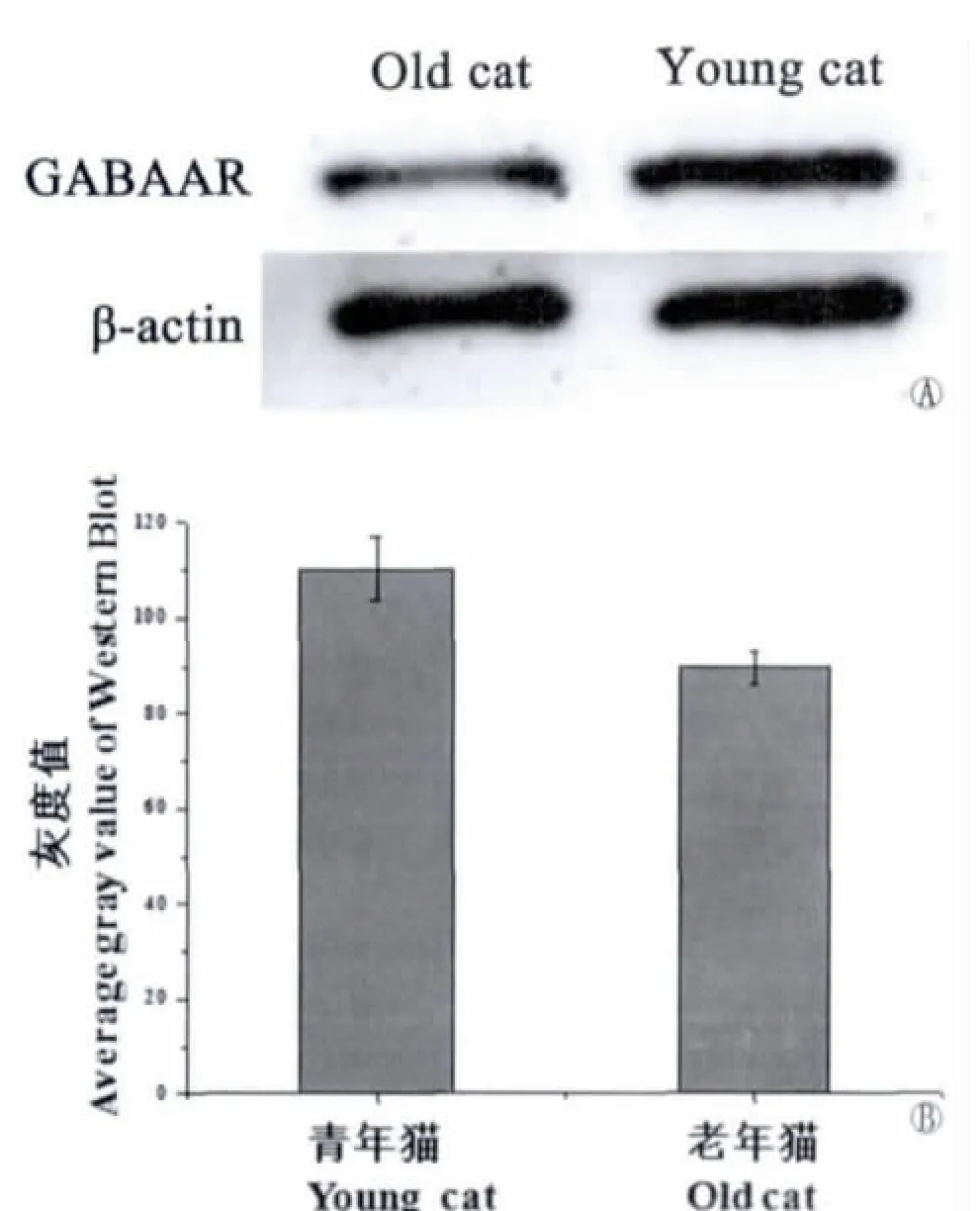
图3 Western Blot显示青年猫与老年猫AI区GABAA受体的表达情况:A显示老年猫初级听皮层蛋白含量显著下降;B显示Image-Pro Express 5.1图像分析软件测量的灰度值Fig.3Western blot demonstration the expression of GABAAR.A:Decline of protein level in A1of old cat.B:Average gray value of Western Blot was measured by Image-Pro Express 5.1software
讨 论
Prieto等[8]等人在对大鼠的初级听皮层GABA及GABA受体年龄性变化研究中发现,GABA及GABAA受体神经元广泛分布在大鼠初级听皮层。我们对青年猫与老年猫AI的GABA[9]及GABAA受体免疫组织化学染色后观察结果与之一致。GABA是听觉神经中枢主要抑制性递质,在听觉功能的实现中起重要作用。研究显示,在脊椎动物中,GABAA受体较GABAB受体分布更为广泛,且GABAA受体和皮层中枢听觉功能关系较为密切[6]。在丘脑,经突触结构中GABAA受体调节快速可塑性抑制,以及经持久的突触外高亲和力GABAA受体发挥全抑制作用。由于兴奋与抑制之间的精确平衡,随着年龄增长,丘脑抑制功能改变和GABAA受体音调调节抑制作用减弱可能破坏听觉信号的处理、听觉域、丘脑皮层的节律和曼波睡眠[4]。已有研究显示,在猫AI区沿着背腹轴Ⅲ、Ⅳ层显著的频率调谐功能与其起主要抑制作用的GABA 含量较高相适应[9,11,12],而 GABA 的抑制作用主要是由GABAA受体介导。本文实验结果从形态学角度有力支持了以上观点。
实验结果进一步显示,老年猫初级听皮层总的GABAA-IR神经元密度明显降低,老年猫初级听皮层GABAA-IR神经元占皮层神经元总数的百分比比青年猫的减少了5.32%,GABAA受体蛋白含量下降了23.16%。这与 Turner[13]和Leventhal[13]等人对大鼠和猴听觉系统皮层下中枢下丘及运动皮层和视皮层年龄相关比较研究结果相一致。老年大鼠与老年猴的初级视皮层神经元在外加GABA及GABAA受体激动剂作用下,可以使其较好的恢复对方向、方位选择性分辨,提示视觉功能的衰退很可能与GABAA受体介导的GABA能抑制作用减弱相关[14]。Richardson等[4]研究发现:大鼠丘脑或内侧膝状体(medial geniculate body,MGB)听觉通路中GABAA受体自我平衡功能随年龄变化,与青年大鼠相比,在内侧膝状体,老年大鼠GABAA受体密度减小45.5%,丘系与非丘系突触GABAA受体亦表现出显著的选择性年龄相关损伤效应。已有研究表明在AI区和AII区相互通联的神经元解剖结构不同,可能导致其具有特殊的整合性能[7]。强有力的抑制性GABAA受体对突触外受体反应性调节,可以较好地解释听觉丘脑核团神经元如何处理听觉信号[15]。听觉系统得以行使正常的听觉功能,必要条件之一是兴奋与抑制作用的相互平衡,由于GABAA受体介导的抑制功能减弱使听觉中枢失去平衡,从而影响正常的听觉功能。我们观察到初级听皮层GABAA受体含量显著减少,可能影响正常的抑制作用,推测其亦可能是导致老年性耳聋的重要原因。本文所观察到初级听皮层GABA受体表达增龄性下调,推测与老年机体神经中枢的复杂变化有关,同时也提示GABA受体表达下调可能是对初级听皮层神经元的保护具有一定关系。
总之,本实验虽然观察到初级听皮层GABAA受体免疫阳性反应与蛋白表达存在明显增龄性变化,但其较深层次的神经机制仍相当复杂,有待今后进一步研究。
[1]Gates GA,Mills JH.Presbycusis.Lancet,2005,366(9491):1111-1120
[2]Xu J,Yu L,Cai R,et al.Early continuous white noise exposure alters auditory spatial sensitivity and expression of GAD65and GABAAreceptor subunits in rat auditory cortex.Cereb Cortex,2010,20(4):804-812
[3]Caspary DM,Holder TM,Hughes LF.et al.Age-related changes in GABAAreceptor subunitcomposition and function in rat auditory system.Neuroscience,1999,93:307-312
[4]Richardson BD,Ling LL,Uteshev VV,et al.Reduced GABAAreceptor-mediated tonic inhibition in aged rat auditory thalamus.J Neurosci,2013,33(3):1218-1227
[5]Mendelson JR,Lui B.The effects of aging in the medial genicukate nucleus:a comparison with the inferior colliculus and auditory cortex.Hearing Research,2004,191(1-2):21-23
[6]Fubara BM,Casseday JH,Covey ES,et al.Distribution of GABAA,GABAB,and Glycine receptors in the central auditory system of the big brown bat,Eptesicus fuscus.J Comp Neurol,1996,369(1):83-92
[7]Yuan K,Shih JY,Winer JA,et al.Functional networks of parvalbumin-immunoreactive neurons in cat au-ditory cortex.J Neurosci,2011,31(37):13333-13342
[8]Turner JG,Hughes LF,Caspary DM.Affects of aging on receptive fields in rat primary auditory cortex layer V neurons.J Neurophysiol,2005,94(4):2738-2747
[9]罗勋,华田苗,孙庆艳等.猫初级听皮层γ-氨基丁酸能神经元和星形胶质细胞年龄相关变化.解剖学报,2006,37(5):514-519
[10]彭建安,郭萍,瘳家万等.大鼠纹状体GABA受体的表达及年龄性变化.中国组织化学与细胞化学杂志,2008,17(3):323-326
[11]Schreiner CE,Mendelson JR.Functional topography of cat primary auditory cortex:distribution of integrated excitation.J Neurophysiol,1990,64(5):1442-1459
[12]Horikawa J,Hosokawa Y,Kubota M,et al.Optical imaging of spatiotemporal patterns of glutamatergic excitation and GABAergic inhibition in the guinea-pig auditory cortex in vivo.J Physiol,1996,497(3):629-638
[13]Turner JG,Caspary DM.Comparison of two rat models of aging:peripheral pathology and GABA changes in the inferior colliculus.In:(Syka,Merzenich)Plasticity of the Central Auditory System and Processing of Complex Acoustic Signals.London:Kluwer/Plenum,2005,217-225
[14]Leventhal AG,Wang Y,Zhou PM,et al.GABA and its agonists improved visual cortical function in senescent monkeys.Science,2003,300(5620):812-815
[15]Ben DR,Lynne LL,Victor VU,et al.Extrasynaptic GABAAReceptors and Tonic Inhibition in Rat Auditory Thalamus.PLoS One,2011,6(1):e16508
猜你喜欢
北京航空航天大学学报(2022年6期)2022-07-02
影像诊断与介入放射学(2021年5期)2021-11-29
天津医科大学学报(2021年1期)2021-01-26
中国医学影像技术(2020年11期)2021-01-04
中国现代医药杂志(2020年3期)2020-05-08
中国生物医学工程学报(2019年6期)2019-07-16
中国组织化学与细胞化学杂志(2017年1期)2017-06-15
自动化学报(2017年5期)2017-05-14
中国医学科学院学报(2015年5期)2015-03-01
浙江理工大学学报(自然科学版)(2015年7期)2015-03-01
