
黄石采石场废弃地植物的生长与光合特性研究
2014-01-04 09:07卢广超许建新杨振意吴彩琼邵怡若
中南林业科技大学学报 2014年3期
卢广超,许建新,,薛 立,杨振意,吴彩琼 ,邵怡若
(1.华南农业大学 林学院,广东 广州510642;2.深圳市铁汉生态环境股份有限公司,广东 深圳518040)
采石场的开采严重破坏了植被和土壤,形成了大量的裸露岩石斜坡。由于缺少表土覆盖,造成土壤温差大、保水能力差[1-3]、养分利用低等,限制植物的生长[4-5]。通过控制水土流失和提供绿化[6]等人为干预措施,能使采石场废弃地已被破坏的环境得以稳定和复绿[3],甚至被重新使用[7]。因此选择适于采石场废弃地生长的植物对于其生态重建非常必要。光合和荧光是植物重要的合成代谢过程,可以反映植物生长及对环境条件的适应能力[8]。例如,通过测量植物的光合和荧光特性,Monneveux等[9]研究了不同基因型的硬质小麦的野外生长状况,Wu等[10]研究了白刺花Sophora davidii对不同水分和氮供应的响应情况,谭雪红等[11]评价了高速公路5种植物的抗旱性。
湖北省黄石市矿产丰富,多年来大规模的掠夺性开采形成了大量废弃地,面积达52多hm2。为了改善环境,黄石市于2009年7月通过搭建种植槽、挡土墙和飘台对其进行了生态重建[12]。目前对飘台等客土重建模式下的植被生长研究较少[13-14]。作者对黄石市的种植槽、挡土墙和飘台的植物生长和光合特性进行研究,可以为采石场废弃地生态重建中的植物选择提供参考。
1 材料与方法
1.1 研究地区概况
研究区位于湖北省黄石市黄荆山脉北麓5号塘口(30°10’N,115°03’E),属亚热带季风气候,年平均气温17 ℃,年平均降水量1 400 mm。5号塘口为废弃水泥厂开采坡面,岩性为灰岩或石灰岩,坡度近90°,自然植被荡然无存。
1.2 试验设置和材料
通过在坡顶搭建种植槽(以下称种植槽)、坡下建挡土墙种植台(以下称挡土墙)和坡面搭建飘台种植槽(以下称飘台)对坡面进行工程改良,覆盖客土。根据“因地制宜、适地适树”原则种植若干种乡土植物。乔木有大叶女贞Ligustrum lucidum、樟树Cinnamomun camphora,灌木有小叶女贞Ligustrum quihoui、冬青卫矛Buxus megistophylla、 夹 竹 桃Nerium oleander和插田泡Rubus coreanus,藤本有扶芳藤Euonymus fortunei。乔木株行距为1 m,灌木株行距为40 cm,藤本株行距为20 cm;乔木每个穴种植1株,灌木2株,藤本是2~3株,生态重建工程于2010年7月顺利完成施工,并进行浇水、看护,2013年3个样地均完全郁闭。
1.3 测定指标和方法
2011年4月分别在种植槽、挡土墙和飘台建立样地,测定种植植物地径和树高。2013年4月调查各样地的植物地径和树高生长情况及气体交换和叶绿素荧光参数。
用美国LI-COR公司生产的LI-6400便捷式光合测定仪测定叶片气体交换参数,包括叶片光合速率Pn、蒸腾速率Tr、气孔导度Gs和胞间CO2浓度Ci,并计算气孔限制值Ls和叶片水分利用率RWUE。每种植物选3株个体上的3枚成熟叶,在光合作用比较活跃的9:00~11:00进行测定,每个叶片测定3个重复。
叶片叶绿素荧光参数测定与叶片气体交换参数同步测量,采用脉冲调制荧光仪OS-1P(OPTISCIENCES,美国)将测定测试叶片在暗适应20 min后,在非化学淬灭(NPQ)模式下设定每次测量时间为1 min,每次测量3次重复,测定内容有初始荧光(Fo)、最大荧光(Fm)、任意时间实际荧光(F′)、光适应下最大荧光(Fm′)。计算PSII最大光能转换效率Fv/Fm=(Fm-Fo)/Fm(暗)、实际光化学效率 Yield=(Fm′-F′)/Fm′、非光化学淬灭系数 NPQ=Fm/Fm′-1。
1.4 数据处理软件
运用Microsoft Excel 2003软件对数据进行统计分析和作图,运用SAS软件进行DUNCUN多重比较和逐步回归分析。
2 结果与分析
2.1 植物生长状况
2.1.1 种植槽植物的生长状况
经过2年复绿,冬青卫矛、夹竹桃、插田泡3种灌木树种的年平均地径生长量分别为0.14、0.45、0.28 cm,而年平均高生长量分别为0.09、0.31、1.10 m(见表1)。夹竹桃地径生长较大,插田泡高生长较快。藤本植物扶芳藤年平均地径生长量为0.06 cm,年平均高生长量为0.49 m,地径和高同比年平均分别增加16%和213%。种植槽的灌木植物年平均地径生长量仅为0.29 cm,高为0.51 m,同比年平均分别增加18%和54%,高生长明显要快于地径生长。

表1 种植槽的植物生长状况Table 1 Growth status of plants in planting trough plot
2.1.2 挡土墙植物的生长状况
经过2年复绿,樟树和大叶女贞2种乔木树种的年平均地径生长量分别为1.14、1.40 cm,年平均高生长量分别为1.19和1.36 m,大叶女贞的地径和高生长均快于樟树(见表2)。灌木树种小叶女贞和夹竹桃的年平均地径生长量分别为1.29和0.89 cm,而年平均高生长量分别为1.14 和0.57 m,小叶女贞地径和高生长均快于夹竹桃。 挡土墙乔木的年平均地径生长量为1.27 cm,高为1.28 m,同比年平均分别增长25%和59%;灌木年平均地径生长量为1.09 cm,高为0.86 m,同比平均升高97%和118%。乔木和灌木的高生长均明显要快于地径生长。

表2 挡土墙植物的生长状况Table 2 Growth status of plants in retaining wall plot
2.1.3 飘台植物的生长状况
经过2年复绿,乔木树种大叶女贞年平均地径生长量为1.00 cm,年平均高生长量为0.69 cm,地径和高同比年平均分别增加41%和42%(见表3)。灌木树种小叶女贞、冬青卫矛和夹竹桃的年平均地径生长量分别为1.04、0.02和0.51 cm,而年平均高生长量分别为0.77、0.14和0.58 m。小叶女贞的地径和高生长最快。飘台灌木年平均地径生长量为0.53 cm,高为0.50 m,同比分别增加了37%和53%,高生长均明显要快于地径生长。

表3 飘台各植物的生长状况Table 3 Growth status of plant in floating plate plot
2.2 植物叶片的气体交换参数
2.2.1 种植槽4种植物叶片的气体交换参数
种植槽的夹竹桃、冬青卫矛、插田泡和扶芳藤的净光合速率Pn分别为3.15、2.22、1.88、1.27µmol•m-2s-1,夹竹桃显著大于其它植物,扶芳藤显著小于其它植物(见图1,P<0.05);蒸 腾 速 率Tr分 别 为 0.42、0.41、0.27和 0.43 mol•m-2s-1,气孔导度Gs分别为 0.038、0.037、0.024和0.039 mol•m-2s-1,均为插田泡显著小于其它植物(P<0.05);胞间CO2浓度Ci分别为 250.13、297.85、265.01 和 341.39 μmol•mol-1,扶芳藤显著大于其它植物,夹竹桃与插田泡显著小于其它植物(P<0.05);气孔限制值Ls分别为0.38、0.26、0.35和0.16,夹竹桃和插田泡显著大于其它植物,扶芳藤显著小于其它植物(P<0.05);水分利用率分别为8.02、5.44、7.41和3.00 μmol•mol-1,夹竹桃与插田泡显著大于其它植物(P<0.05)。
2.2.2 挡土墙4种植物叶片的气体交换参数
挡土墙中樟树、大叶女贞、小叶女贞和夹竹桃的净光合速率Pn分别为2.26、5.47、2.55和2.40 µmol•m-2s-1,大叶女贞显著高于其它 植物(P<0.05)(见图2);蒸腾速率Tr分别为0.68、0.86、0.46 和 0.33 mol•m-2s-1,表现为大叶女贞>樟树>小叶女贞>夹竹桃(P<0.05);气孔导度Gs分别为0.062、0.129、0.042和0.030 mol•m-2s-1,表现为大叶女贞>樟树>小叶女贞>夹竹桃(P<0.05);胞间CO2浓度Ci分别为328.38、325.47、304.45 和 261.76 μmol•mol-1, 夹竹桃显著小于其它植物(P<0.05);气孔限制值Ls分别为0.20、0.20、0.29和0.37,小叶女贞和夹竹桃显著大于其它植物(P<0.05);水分利用率RWUE分别为 4.10、6.89、 6.48 和 6.70 μmol•mol-1,樟树显著低于其它植物(P<0.05)。
2.2.3 飘台4种植物叶片的气体交换参数
飘台的大叶女贞、小叶女贞、冬青卫矛和夹竹桃的净光合速率Pn分别为2.85、2.65、2.21和 2.35 µmol•m-2s-1,冬青卫矛显著小于其它植物(见图3,P<0.05);蒸腾速率Tr分别为0.32、0.29、0.36 和 0.51 mol•m-2s-1,表现为夹竹桃显著大于其它植物,小叶女贞显著小于其它植物(P<0.05);气孔导度Gs分别为0.028、0.025、0.030和0.049 mol•m-2s-1,夹竹桃显著大于其它植物(P<0.05);胞间CO2浓度Ci分别为173.52、242.75、305.31 和 313.44 μmol•mol-1, 夹 竹 桃 和冬青卫矛显著大于其它植物,大叶女贞显著小于其它植物(P<0.05);气孔限制值Ls分别为0.58、0.43、0.26和0.23,表现为大叶女贞显著高于其它植物,夹竹桃和冬青卫矛显著低于其它植物(P<0.05);RWUE分别为12.06、9.28、5.22和5.19 μmol•mol-1,大叶女贞显著高于其它植物,小叶女贞处于中间水平,夹竹桃和冬青卫矛显著低于其它植物(P<0.05)。

图1 种植槽4种植物叶片的气体交换参数Fig. 1 Gas exchange parameters of leaves of four plants in planting trough plot

图2 挡土墙4种植物叶片的气体交换参数Fig. 2 Gas exchange parameters of leaves of four plants in retaining wall plot
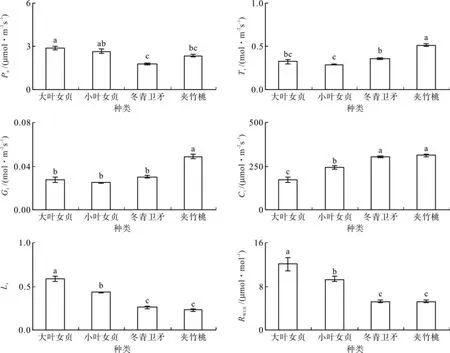
图3 飘台4种植物叶片的气体交换参数Fig. 3 Gas exchange parameters of leaves of four plants in fl oating plate plot
2.3 植物叶片叶绿素荧光参数
2.3.1 种植槽4种植物叶片的叶绿素荧光参数
Fo是光系统Ⅱ反应中心处于完全开放时的最低荧光产量,代表的是色素所吸收的光能中不参与光化学反应的能量;Fv/Fm指最大光化学效率(原初光能转化效率),反映PSⅡ反应中心内禀光能转换效率,ETR指表观光合电子传递速率,非光化学淬灭系数NPQ反映的是PSⅡ吸收的光能中不能用于光合电子传递而以热的形式耗散掉的光能部分。种植槽的夹竹桃、冬青卫矛、插田泡和扶芳藤的初始荧光Fo分别为138.63、168.92、125.66和199.89,扶芳藤显著大于其它植物,夹竹桃与插田泡显著小于其它植物(见图4,P<0.05);PSⅡ最大光能转换效率Fv/Fm分别为0.76、0.79、0.81和0.68,夹竹桃、冬青卫矛、插田泡基本都在0.8左右,扶芳藤显著小于其它植物(P<0.05);实际光化学效率Yield分别为0.13、0.06、0.15和0.07,表现为夹竹桃和插田泡显著大于其它植物(P<0.05);非光化学淬灭系数NPQ分别为1.86、2.26、2.49和2.27,表现为插田泡显著大于其它植物,夹竹桃显著小于其它植物,冬青卫矛与插田泡处于中间水平(P<0.05)。
2.3.2 挡土墙4种植物叶片的叶绿素荧光参数
挡土墙中樟树、大叶女贞、小叶女贞和夹竹桃的初始荧光Fo分别为134.78、124.35、133.15和151.36,夹竹桃显著高于大叶女贞(P<0.05)(见图5);PSⅡ最大光能转换效率Fv/Fm分别为0.78、0.79、0.79和0.79,均处于正常水平(P<0.05);实际光化学效率Yield分别为0.08、0.16、0.14和0.13,大叶女贞显著大于其它植物,樟树显著小于其它植物(P<0.05);非光化学淬灭系数NPQ分别为2.42、1.83、2.49和1.76,樟树和小叶女贞显著大于大叶女贞和夹竹桃(P<0.05)。
2.3.3 飘台4植物叶片的叶绿素荧光参数
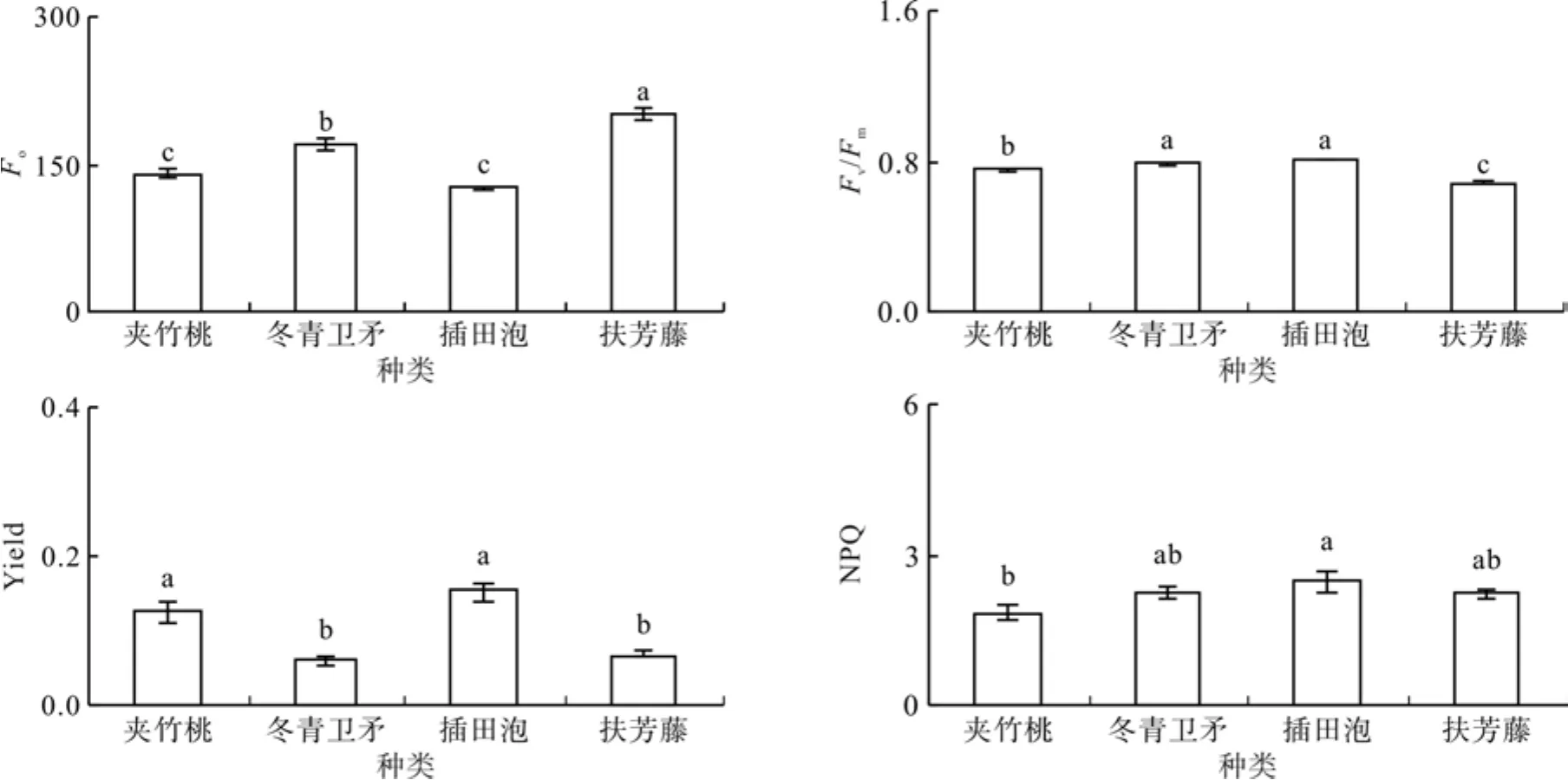
图4 种植槽4种植物叶片的叶绿素荧光参数Fig. 4 Chlorophyll fl uorescence parameters of leaves of four plants in planting trough plot

图5 挡土墙4种植物叶片的叶绿素荧光参数Fig. 5 Chlorophyll fl uorescence parameters of leaves of four plants in retaining wall plot
大叶女贞、小叶女贞、冬青卫矛和夹竹桃的初 始 荧 光Fo分 别 为 135.26、115.68、191.95和150.77,表现为冬青卫矛>夹竹桃>大叶女贞>小叶女贞(P<0.05)(见图6);PSⅡ最大光能转换效率Fv/Fm(暗)分别为0.80、0.82、0.78和0.80,均处于正常水平(P<0.05);实际光化学效率Yield分别为0.10、0.11、0.04和0.08,冬青卫矛显著低于其它植物(P<0.05);非光化学淬灭系数NPQ分别为2.38、2.48、2.00和1.73,表现为大叶女贞和小叶女贞较高,夹竹桃较低,显著小于前两者(P<0.05)。
2.4 植物年平均生长量与叶片光合特性回归关系
应用SAS统计软件中逐步回归的方法,以2011~2013年各植物的年平均地径和高生长量为因变量,叶片的净光合速率Pn、蒸腾速率Tr、气孔导度Gs、胞间CO2浓度Ci、气孔限制值Ls、叶片水分利用率RWUE和叶片的初始荧光Fo、PSⅡ最大光能转换效率Fv/Fm(暗)、实际光化学效率Yield、非光化学淬灭系数NPQ作为自变量,建立植物的年平均生长量对叶片气体交换参数和叶绿素荧光参数的逐步回归模型。
逐步回归模型的结果(见表4)可以看出,植物年平均地径生长量与蒸腾速率、初始荧光显著相关,与蒸腾速率的相关性较大(P< 0.15)。植物的年平均高生长量与胞间CO2浓度、初始荧光显著相关,与初始荧光的相关性较大(P<0.15)。各样地植物年平均地径生长量和平均高生长量基本上与初始荧光Fo表现为负相关,而植物年平均地径生长量与蒸腾速率、平均高生长量与胞间CO2浓度为正相关。
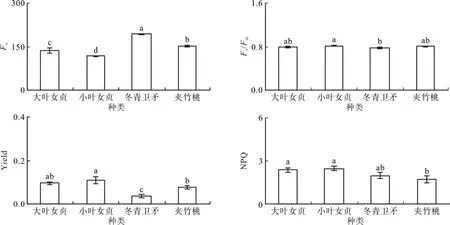
图6 飘台4种植物叶片的叶绿素荧光参数Fig. 6 Chlorophyll fl uorescence parameters of leaves of four plants in fl oating plate plot

表4 叶片光合特性与植物生长量的逐步回归模型†Table 4 Stepwise regression analysis between photosyntheticcharacteristics and average annual growth of plants
3 讨 论
3.1 恢复植被的生长状况
采石场的自然恢复过程极其缓慢[3],往往要花费几十年到一个世纪以上[15-16]。通过人工恢复重建能够短时间内实现采石场的复绿,提高植被覆盖率[17]。本研究中构建的种植槽、挡土墙和飘台的工程措施改良了采石场的坡面结构,通过覆盖客土种植乔木、灌木、藤本植物,加速了采石场的生态恢复进程,采取工程措施3年后样地的复绿情况良好。
3个样地的植物生长均表现为年平均高生长大于地径生长。有学者研究表明,植物群落郁闭后,地/胸径生长相对缓慢,而对树高影响较弱[18]。经过2年的复绿,植物生长使林分郁闭提高,导致地径生长变慢,而高生长相对较快。
3.2 矿区植物叶片的气体交换与叶绿素荧光参数
光合参数是光合作用效率的外在表现,叶绿素荧光参数是光合效率的内部引起因素[19]。光合速率反应了植物固定CO2的能力,而叶绿素荧光在测定叶片光合作用过程中光系统对光能的吸收、传递、转换、耗散、分配等方面具有独特的作用[20]。
净光合速率是光合系统功能的直接体现,其大小表明光合能力的强弱。初始荧光与激发光的强度和叶绿素浓度有关,代表的是色素所吸收的光能中不参与光化学反应的能量,与光合作用的光反应无关[21]。在相同条件下,初始荧光较高,叶片色素吸收的能量中流向光化学作用的部分较少,初始荧光相对较低,流向光化学作用的能量就相对较高。种植槽的夹竹桃、挡土墙和飘台的大叶女贞和小叶女贞的净光合速率较大,而初始荧光相对较小,地径和高生长较快,说明这些植物对采石场废弃地的恶劣环境适应力强,光能利用率高,叶片色素吸收的能量用于光合作用的部分多,所以以上植物成为采石场废弃地生态恢复的优选树种。插田泡的净光合速率中等,初始荧光较小,实际光化学效率大,说明其实际光量子效率高,因而地径和高生长较快。其它植物具有较低的净光合速率,但是初始荧光通常较高,说明它们吸收光能的能力强,但是吸收的光能中不能用于光合电子传递而进行光合作用的部分多,大部分光能以热的形式耗散[22]。其中樟树具有较高的非光化学淬灭系数值,说明它吸引光能的能力极强, 但光能的转化率低, 吸收的光能主要以热的形式耗散[23],但是适应开阔的采石场废弃地高光照条件, 说明其对于过强的光照具有良好的自我保护能力,能适应采石场废弃地的恶劣环境,可以成为采石场废弃地生态恢复的参考树种;冬青卫矛的净光合速率中等或低,初始荧光较高,扶芳藤的净光合速率低,初始荧光较高,最大光能转换系数仅为0.68,小于0.8的正常生长临界值[24],表明这两种植物的光能利用率低,不适合采石场废弃地的恶劣环境;挡土墙和飘台的夹竹桃的净光合速率和非光化学淬灭系数低,而初始荧光较高,表明其热耗散能力低,不利于利用适应开阔采石场废弃地的高光照条件,对于过强光照的自我保护能力差,不适应采石场废弃地的恶劣环境[25]。
3.3 植物生长状况与光合生理特性逐步回归关系
通过逐步回归分析得出植物的年平均地径生长量与蒸腾速率呈正相关,与初始荧光呈负相关。蒸腾速率越大,年平均地径生长量越高。蒸腾促进生长的原因可能有:1)蒸腾作用促进了植物被动吸水与转运水分,保证有充足的水参与光合作用、呼吸作用、有机物的合成和分解过程中;2)蒸腾作用将土壤中的无机养分运输到植物各部分,保证了植物生长所需养分;3)蒸腾作用使气孔保持开放,有利于CO2通过气孔进入细胞,提高了植物的光合速率[26-31]。初始荧光大表明色素所吸收的光能中不参与光化学反应的能量大,流向光化学作用的部分较少,光合效率低,不利于植物生长[22]。
对植物年平均高生长量逐步回归分析得出植物年平均高生长量与胞间CO2浓度呈正相关,与初始荧光呈负相关。二氧化碳是光合作用的原料,对光合速率影响很大,光合速率随CO2浓度的提高而增大。胞间CO2浓度越高,表明植物在光合作用中吸收的CO2越多,制造的碳水化合物就越多,高生长也就越快。本研究中初始荧光相对较低的植物实际光量子效率相对较高,导致了较高的光合速率,所以年平均高生长量大。
[1] Wang Z, Wu L, Liu T. Revegetation of steep rocky slopes:Planting climbing vegetation species in artif i cially drilled holes [J].Ecological Engineering, 2009, 35(7): 1079-1084.
[2] Beikircher B, Florineth F, Mayr S. Restoration of rocky slopes based on planted gabions and use of drought-preconditioned woody species [J]. Ecological Engineering, 2010, 36(4):421-426.
[3] Meira-Neto J A A, Clemente A, Oliveira G, et al. Post-fire and post-quarry rehabilitation successions in Mediterraneanlike ecosystems: Implications for ecological restoration [J].Ecological Engineering, 2011, 37(8): 1132-1139.
[4] Rowe E C, Healey J R, Edwards-Jones G, et al. Fertilizer application during primary succession changes the structure of plant and herbivore communities [J]. Biological Conservation,2006, 131(4):510-522.
[5] 杨振意, 薛 立, 许建新. 采石场废弃地的生态重建研究进展[J]. 生态学报, 2012, 32(16): 5264-5274.
[6] Zhang H, Chu L M. Plant community structure, soil properties and microbial characteristics in revegetated quarries [J].Ecological Engineering, 2011, 37(8): 1104-1111.
[7] Milgrom T. Environmental aspects of rehabilitating abandoned quarries: Israel as a case study [J]. Landscape and Urban Planning, 2008, 87(3): 172-179.
[8] 朱 琳, 潘国富, 杜广明, 等. 呼伦贝尔草原三种优势植物叶绿素荧光特性的比较[J].中国草地学报, 2012, 34(4): 82-86.
[9] Monneveux P, Rekika D, Acevedo E, et al. Effect of drought on leaf gas exchange, carbon isotope discrimination, transpiration efficiency and productivity in field grown durum wheat genotypes [J]. Plant Science, 2006, 170(4): 867-872.
[10] Wu F Z, Bao W K, Li F L, et al. Effects of water stress and nitrogen supply on leaf gas exchange and fl uorescence parameters of Sophora davidii seedlings [J]. Photosynthetica, 2008, 46(1):40-48.
[11] 谭雪红, 高艳鹏, 郭小平, 等.五种高速公路边坡绿化植物的生理特性及抗旱性综合评价[J]. 生态学报, 2012, 32(16):5076-5086.
[12] 周 琼, 许建新, 秦永宏, 等.黄石市岩质边坡生态修复综合技术应用探[J]. 中国水利, 2011, (2): 36-38.
[13] 陆志敏, 吴鹏敏, 汤社平, 等.废弃采石场绿化树种选择及其配套技术研究[J]. 浙江林业科技, 2006, 26(3):59-65.
[14] 鲁统春, 高德武, 王创争, 等. 废弃采石场植被快速恢复研究[J]. 水土保持研究, 2006, 13(6):210-212.
[15] Novák J, Prach K. Vegetation succession in basalt quarries:Pattern on a landscape scale [J]. Applied Vegetation Science,2003, 6(2): 111-116.
[16] Novák J, Konvicka M. Proximity of valuable habitats affects succession patterns in abandoned quarries [J]. Ecological Engineering, 2006, 26(2): 113-122.
[17] Novák J, Prach K. Artif i cial sowing of endangered drygrassland speciesin to disused basalt quarries [J]. Flora, 2010, 205:179-183.
[18] 方发之, 陈素灵, 薛 扬. 立地与密度对黄桐中幼林生长量影响的研究[J]. 热带林业, 2008, 36(1):25-29.
[19] 王玉珏, 付秋实, 郑 禾, 等. 干旱胁迫对黄瓜幼苗生长、光合生理及气孔特征的影响[J]. 中国农业大学学报, 2010,15(5): 12-18.
[20] 徐伟红, 郭卫华, 徐 飞, 等. 三种枣树叶绿素荧光参数的日变化[J]. 山东农业科学, 2007, (2): 29-32.
[21] 云 菲, 刘国顺, 史宏志, 等. 光氮互作对烤烟叶片光合色素及荧光特性的影响[J]. 中国烟草学报, 2010, 16(4):44-50.
[22] 吕芳德, 徐德聪, 侯红波. 5种红山茶叶绿素荧光特性的比较研究[J]. 经济林研究, 2003, 21 (4):4-7.
[23] Badger M R, Caemmerer S, Ruuska S, et al. Electron flow to oxygen in higher plants and algae: rates and control of direct photoreduction (Mehler reaction) and rubisco oxygenase [J].Biological Sciences, 2000, 14 (2):1433-1445.
[24] 周蕴薇, 刘艳萍, 戴思兰. 用叶绿素荧光分析技术鉴定植物抗寒性的剖析[J]. 植物生理学通讯, 2006, 42(5):945-950.
[25] 张 谧, 王慧娟, 于长青. 北京西山采石场不同自然恢复期主要植物叶绿素荧光特征[J]. 环境保护科学, 2010, 36(3):67-69.
[26] 房 用, 慕宗昭, 王月海, 等. 116个杨树无性系蒸腾特性及其影响因子研究[J]. 山东大学学报:理学版, 2006, 41(6):168-171.
[27] 何 茜, 李吉跃, 姜 枫.毛白杨无性系抗旱节水特性与评价[M]. 北京:中国林业出版社, 2010.
[28] 刘青柏,刘明国,刘明忠,等. 朝阳地区不同品种枣树光合及水分利用特征研究[J].经济林研究,2013,31(3):16-20.
[29] 李芳东,吕德国,杜国栋,等.生草覆盖对高温强光下苹果叶片光合特性的影响[J].经济林研究,2013,31(4): 67-72.
[30] 王旭军,潘百红,程 勇,等.红榉不同种源光合特性的比较[J].中南林业科技大学学报,2013,33(6):37-42.
[31] 蒋 燚,李志辉,朱积余,等.不同红锥种源幼林光合特性日变化测定与分析[J].中南林业科技大学学报,2013,33(6):43-47.
猜你喜欢
名家名作(2022年7期)2022-08-05
安徽文学(2022年7期)2022-07-14
文萃报·周五版(2022年24期)2022-06-21
天天爱科学(2021年2期)2021-09-10
疯狂英语·新阅版(2021年1期)2021-03-03
疯狂英语·新悦读(2021年1期)2021-01-27
意林(2020年22期)2020-12-04
海外文摘·文学版(2020年12期)2020-01-03
检察风云(2015年3期)2015-02-12
筑路机械与施工机械化(2014年8期)2014-03-01
