
饲用木聚糖酶基因的克隆及序列分析
2013-10-25 05:42四川农业大学生命科学与理学院韩学易李治国
中国饲料 2013年4期
四川农业大学生命科学与理学院 韩学易 李治国 陈 惠
木聚糖酶属于水解酶类,是一类可以将木聚糖降解成低聚木糖或木糖的复合酶系(苏玉春等,2008)。目前研究和应用较多的木聚糖酶多是来源于细菌、真菌和放线菌。较真菌所产木聚糖酶而言,细菌所产木聚糖酶具有更好的热稳定性(Panbangred等,1983)。本文报道了来自短小芽孢杆菌HZ-01产木聚糖酶基因的克隆及其序列分析,为下一步木聚糖酶的异源表达及其产业化应用奠定良好基础。
1 材料和方法
1.1 载体与菌株 短小芽孢杆菌HZ-01由本实验室 (四川农业大学生物化学与分子生物学实验室)自行筛选;大肠杆菌DH5α为本实验室保存;pMD19-T载体购于TaKaRa公司。引物合成和基因测序由上海英骏生物技术有限公司和北京诺赛基因组研究有限公司完成。
1.2 培养基和主要试剂 大肠杆菌培养基为LB培养基。
T4 DNA连接酶与Taq DNA聚合酶(TaKaRa公司);DNA限制性内切酶 (Fermentas公司);DNA纯化试剂盒 (TANGEN公司);DNA Marker[天根生化科技(北京)有限公司];质粒提取试剂盒(上海Omega生物技术有限公司);蛋白胨和酵母粉(Oxoid 公司);琼脂糖(Amresco 公司);其他均为国产分析纯试剂。
1.3 木聚糖酶基因的扩增 以短小芽孢杆菌HZ-01基因组DNA为模板,根据网上木聚糖酶基因序列设计引物:
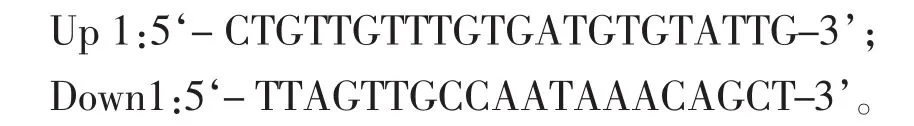
PCR扩增体系:总DNA 1 μL,上下游引物各1 μL,2X Taq PCR MasterMix 12.5 μL,ddH2O 9.5 μL,总体积 25 μL。 PCR 扩增条件:94 ℃预变性 2 min,94 ℃变性 1 min,55 ℃退火 50 s,72 ℃延伸1 min,30个循环,最后72℃延伸4 min。电泳检测PCR扩增产物,纯化回收,连接到T载体上,转入感受态细胞,菌落PCR鉴定阳性转化子。DNA测序由北京诺赛基因组研究有限公司完成。
1.4 木聚糖酶基因的序列分析 由网上提供的信息学数据库和分子生物学软件分析短小芽孢杆菌HZ-01木聚糖酶基因序列,并进行生物信息学预测。
2 结果与分析
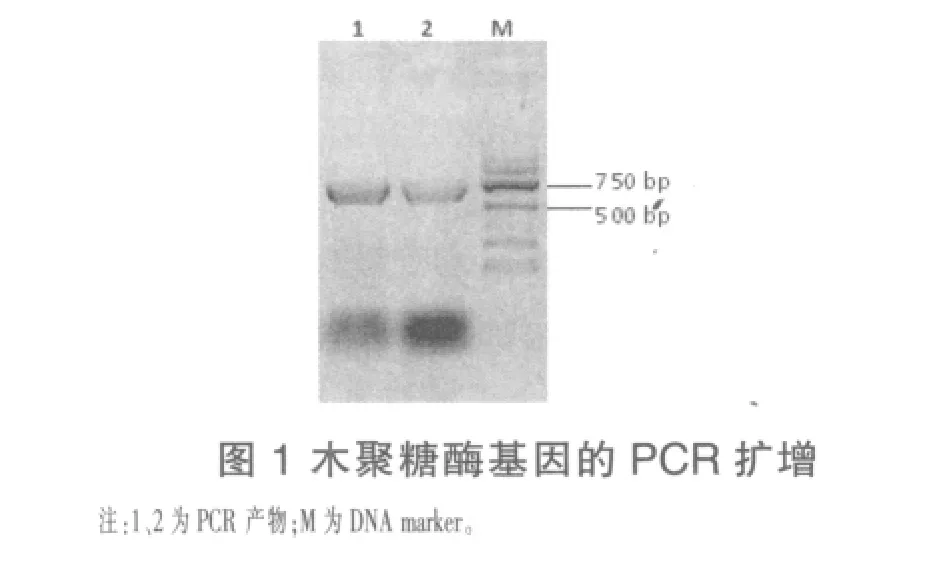
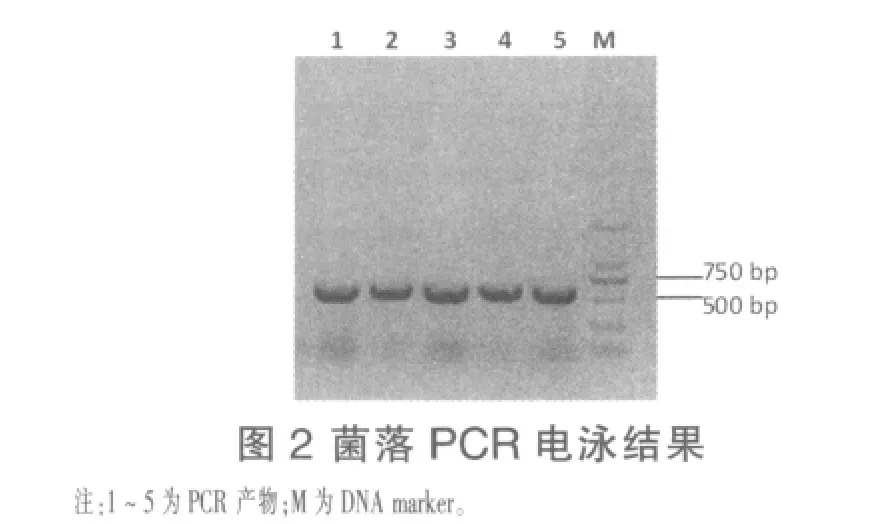
2.1 木聚糖酶基因的克隆 以短小芽孢杆菌HZ-01基因组DNA为模板进行PCR扩增,产物经琼脂糖凝胶电泳后,得到一条大小约700 bp的特异条带(图1)。将目的条带回收,连接到pMD19-T载体,转入大肠杆菌,阳性克隆经菌落PCR检测(图2)及测序鉴定,表明成功克隆目的基因。
2.2 木聚糖酶基因的序列分析 PCR扩增产物由北京诺赛基因组研究有限公司测序(图3)。测序结果表明,该木聚糖酶基因全长687 bp,GC含量为43%,共编码228个氨基酸,理论分子质量为 25 kDa,等电点(pI)为 8.96。

采用NetNGlyc1.0在线分析软件进行短小芽孢杆菌HZ-01木聚糖酶氨基酸序列N-糖基化位点(Asn-Xaa-Ser/Thr)的预测。在全长氨基酸序列中共有4个Asn残基可能被糖基化:当阈值为0.5 时,分别为第 3、17、57、63 位。
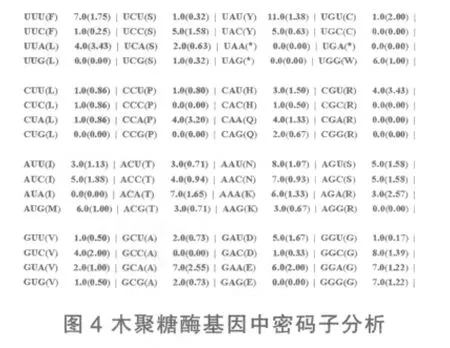
利用MEGA4.0中codon usage对短小芽孢杆菌HZ-01木聚糖酶基因密码子使用频率进行分析,结果见图4。由图4可见,该木聚糖酶基因在密码子选择和使用上有明显偏爱性,谷氨酸和半胱氨酸只出现一种密码子,分别为GAA、UGU,亮氨酸未使用UUG、CUG,脯氨酸未使用CCC、CCG,异亮氨酸未使用AUA,丙氨酸未使用GCC,精氨酸密码子使用具有明显偏爱性,只出现CGU和AGA。
3 讨论
周晨妍等(2005)采用RT-PCR技术扩增了宇佐美曲霉 (Aspergillus usamii)E001木聚糖酶(Xyn II)成熟肽和3’非编码区的cDNA片段,DNA序列分析表明,该cDNA全长733 bp,其中3’非编码区178 bp,成熟肽编码区555 bp,编码184个氨基酸。邬敏辰等 (2007)以宇佐美曲霉E001基因组DNA为模板,采用单侧PCR等技术扩增了木聚糖酶基因xyn I的DNA全序列,该DNA全长1098 bp,含启动子序列、内含子和外显子等核苷酸序列。白爱枝等 (2009)以黑曲霉(Aspergillus niger)A3的基因 组 DNA 为 模板,PCR扩增出了木聚糖酶xynB基因,通过序列分析,xynB基因的cDNA序列大小为678 bp,编码225个氨基酸。本研究从自主分离的短小芽孢杆菌HZ-01基因组中克隆了木聚糖酶基因,该基因全长为687 bp,编码228个氨基酸,理论分子质量约为25 kDa。
采用目前较为成熟的生物信息学在线分析数据库和软件包,来分析短小芽孢杆菌HZ-01木聚糖酶基因的糖基化位点和密码子偏爱性,可以为其进一步的异源高效表达奠定基础。对其糖基化位点分析时,当阈值为0.5时,氨基酸序列中共有4个潜在的 N-糖基化位点,分别为第 3、17、57、63位的Asn残基。密码子偏爱性分析表明,该木聚糖酶基因在密码子选择和使用上有明显偏爱性,因此在进行异源高效表达时需要针对不同宿主进行高频密码子的定点突变或部分片段的人工合成。
本研究拟进一步对其表达载体进行构建,并分别在大肠杆菌和毕赤巴斯德酵母中进行表达,实现木聚糖酶高效表达,对其功能研究及生产应用均具有重要意义。
[1]白爱枝,闫祖威,唐国敏,等.黑曲霉木聚糖酶结构基因和5’调控区基因克隆及其分析[J].华北农学报,2009,24(5):73 ~ 76.
[2]苏玉春,陈光,白晶,等.木聚糖酶的研究进展[J].牧草与饲料,2008,2(4):14 ~ 18.
[3]邬敏辰,王时良,周晨妍.木聚糖酶基因xyn I的克隆和序列分析[J].林产化学与工业,2007,27(6):59 ~ 64.
[4]周晨妍,符丹丹,朱劼,等.宇佐美曲霉木聚糖酶基因的克隆和序列分析[J].食品与发酵工业,2005,31(10):29 ~ 32.
[5]Beg Q,Kapoor M,Mahajan L,et al.Microbiol xylanases and their industrial applications:a review[J].Appl Microbiol Biotechnol,2001,56:326 ~ 338.
[6]Panbangred W,Shinmyo A,Kinoshita S,et al.Purification and properties of endoxylanase produced by Bacillus pumilus[J].Agric Biol Chem,1983,47:957~963.
[7]Wong K,Tan L,Saddler J.Multiplicity of in microorganisms:functions and applications[J].Microbiol Rev,1988,52:305 ~ 317.
猜你喜欢
林产化学与工业(2022年4期)2022-09-13
粮食加工(2021年5期)2021-11-01
复旦学报(医学版)(2021年5期)2021-10-13
福建农业学报(2021年6期)2021-08-18
生物学通报(2020年11期)2020-10-22
湖南饲料(2019年5期)2019-10-15
发明与创新·中学生(2019年6期)2019-06-26
癌症进展(2018年11期)2018-12-30
中成药(2018年7期)2018-08-04
医学研究杂志(2015年7期)2015-06-22
