
血管内皮钙黏蛋白在非小细胞肺癌血管生成中的作用
2013-08-11 15:54顾雪霜周俊豪彭单伊张丽君重庆医科大学附属第一医院呼吸内科重庆400016
吉林大学学报(医学版) 2013年3期
陶 韬,陈 虹,顾雪霜,周俊豪,彭单伊,张丽君(重庆医科大学附属第一医院呼吸内科,重庆 400016)
血管生成是肿瘤生长、浸润和转移的必要条件,抗血管生成疗法成为治疗肿瘤的新策略。目前已发现许多因子参与血管的生成与调控,其中血管内皮钙黏蛋白(vascular endothelial-cadherin,VE-cadherin)是特异性表达于血管内皮细胞的一类细胞黏附分子,在血管形成过程中必不可少[1]。VE-cadherin与多种恶性肿瘤血管生成相关,但在非小细胞肺癌(non-small cell lung cancer,NSCLC)中的研究尚少。本课题组前期研究[2]显示:VE-cadherin在NSCLC组织中的表达明显高于正常组织,且在癌细胞中大量表达,但其是直接对癌细胞起作用还是对血管内皮细胞起作用尚不清楚。本研究采用RNA干扰技术,沉默肺癌细胞中VE-cadherin的表达,并观察VE-cadherin对人脐静脉内皮细胞(human umbilical vein endothelial cells,HUVECs)的影响,为NSCLC的抗血管生成治疗提供新的实验依据。
1 材料与方法
1.1 细胞株、主要试剂及仪器 人肺癌细胞株A549由本实验室保存,HUVECs购自中南大学湘雅医学院。DMEM培养基、RPMI-1640培养基和胎牛血清均购自Gibco公司;TRIzol试剂、Primscript RT reagent Kit试剂盒、SYBR Premix Ex Taq荧光定量PCR试剂盒均购于TaKaRa公司;兔抗VE-cadherin购自Abcam公司;MTS试剂购自Promega公司;Matrigel胶购自BD公司;Lipofectamin RNAi MAX Reagent购自Invitrogen公司;AnnexinⅤ-FITC凋亡检测试剂盒购自Sigma公司;细胞周期检测试剂盒购自南京凯基生物科技发展有限公司;人VE-cadherin-双抗夹心ELISA试剂盒购自上海沪尚生物工程有限公司;Western blotting相关试剂购自碧云天公司;RNA干扰序列[3]:正义链,5′-GGAACCAGAUGCACAUUGAUU-3′;反义链,5′-UCAAUGUGCAUCUGGUUCCUU-3′;NC-siRNA 由上海吉玛制药技术有限公司合成;VE-cadherin引物:上游,5′-TTTCCAGCAGCCTTTCTACCA-3′; 下游,5′-GGAAGAACTGGCCCTTGTCA-3′,扩增产物长度为145bp;GAPDH引物:上游,5′-ACCTGACCTGCCGTCTAGAA-3′;下游,5′-TCCACCACCCTGTTGCTGTA-3′,扩增产物长度为227bp,由上海生工生物工程技术服务有限公司合成。
1.2 细胞培养和转染 人肺癌细胞株A549用含10%FBS的DMEM培养基,置于37℃、5%CO2条件下培养,转染前l d换新鲜培养液。采用RNAi MAX脂质体转染反转录法,细胞分为空白对照组、干扰组和阴性对照组,各组细胞均加500μL Optimer和5μL RNAi MAX,干扰组细胞另加入1.5μL VE-cadherin-siRNA ,阴性对照组细胞另加入1.5μL NC-siRNA,混匀,室温静止20min后,每孔接种2×104细胞于6孔板中,6h后换液。HUVECs分为干扰组和阴性对照组,用含10%FBS的RPMI-1640培养基,置于37℃、5%CO2条件下培养。
1.3 实时定量PCR法检测 VE-cadherin mRNA表达水平 转染48h后,采用TRIzol法提取RNA,将逆转录产物cDNA稀释20倍,逆转录体系及条件、扩增体系均按说明书进行,扩增条件:95℃预变性5min后,95℃、10s,55℃、30s,共40个循环,65℃、5s。独立重复实验3次。
1.4 Western blotting法检测VE-cadherin蛋白表达水平 转染72h后,常规方法提取细胞总蛋白,取蛋白提取物40μg进行SDS-PAGE电泳。在250mA电流下,转膜100min,封闭1h,兔抗VE-cadherin(1∶1000稀释)4℃过夜,TBST 洗5min×5次,与二抗在室温下反应1h,TBST洗5min×5次,ECL法显影。同时,以GAPDH(1∶800稀释)为对照。独立重复实验3次。
1.5 新型四唑氮盐(MTS)比色法检测VE-cadherin作用后A549细胞增殖情况 转染12h后重悬细胞,调整细胞密度为2×104mL-1,每孔100μL接种于96孔板,以此时为0h,分别于0、24、48和72h加入10μL MTS,孵育1h,酶标仪测定各孔于490nm处的光密度(A490)值,每个时间点每组设5个复孔。独立重复实验3次。
1.6 流式细胞术检测VE-cadherin作用后A549细胞周期和凋亡率 转染48h后,收集细胞,采用流式细胞术检测VE-cadherin对A549细胞周期、凋亡的影响,操作步骤按照试剂盒说明书进行。独立重复实验3次。
1.7 ELISA法检测各组 A549细胞上清液VE-cadherin表达水平 细胞转染72h后取上清液,采用人VE-cadherin-双抗夹心ELISA检测试剂盒进行检测,操作按说明书进行。检测各组上清液的A490值,根据标准曲线得出VE-cadherin表达水平。独立重复实验3次。
1.8 MTS比色法检测VE-cadherin作用后HUVECs增殖情况 重悬HUVECs至密度为1×104mL-1,每孔100μL接种于96孔板,6h后干扰组和阴性对照组分别加上清液100μL,以此时为0h,分别于0、24、48和72h加入20μL MTS,酶标仪测定各孔A490值,每个时间点每组设5个复孔。细胞增殖抑制率=(1-干扰组A490值/阴性对照组A490值)×100%。独立重复实验3次。
1.9 流式细胞术检测VE-cadherin作用后HUVECs周期和凋亡率 将8000个/孔HUVECs接种于6孔板,第2天用PBS洗2遍,1∶1加入含10%FBS的RPMI-1640培养基和收集的上清液共2mL。48h后采用流式细胞术检测VE-cadherin对HUVECs周期和凋亡的影响,操作步骤按照试剂盒说明书进行。独立重复实验3次。
1.10 小管形成实验 将Matrigel胶在4℃溶解过夜,用无血清的DMEM培养液稀释Matrigel(1∶1)后,50μL铺于预冷的96孔板中,聚合1h;消化 HUVECs,1000r·min-1离心5min,将干扰组、阴性对照组上清液分别重悬细胞并调整密度至8×105mL-1,50μL种板,6h后200倍倒置显微镜下观察并照相。每组5个随机视野计数小管形成数,取平均值。独立重复实验3次。
1.11 统计学分析 采用SPSS 17.0统计软件对数据进行分析,VE-cadherin的表达水平、细胞A490值、各细胞周期细胞所占比例、细胞凋亡率和小管形成数均以表示,组间比较采用单因素方差分析。
2 结 果
2.1 各组细胞VE-cadherin的表达水平 干扰组细胞VE-cadherin mRNA表达水平(0.299±0.039)明显低于空白对照组(1.137±0.082)和阴性对照组(1.001±0.076)(P<0.05),干扰组较空白对照组和阴性对照组表达水平下调约70%;Western blotting法检测,干扰组细胞的蛋白表达水平(0.297±0.045)明显低于空白对照组(0.833±0.119)和阴性对照组(0.794±0.095)(P<0.05)(图1),而空白对照组和阴性对照组VE-cadherin mRNA和蛋白表达水平差异无统计学意义(P>0.05)。
2.2 VE-cadherin基因沉默后A549细胞增殖情况在0、24、48和72h各组细胞间的A490值比较差异无统计学意义(P>0.05)。见表1。
2.3 VE-cadherin基因沉默后A549细胞周期和细胞凋亡 转染48h后,空白对照组、干扰组和阴性对照组A549细胞在G0/G1、G2/M和S期所占百分比比较差异无统计学意义(P>0.05),见表2。3组 A549细胞凋亡率分别为(11.99±4.77)%、(13.71±4.79)%和(12.02±4.25)%,组间比较差异无统计学意义(P>0.05)。
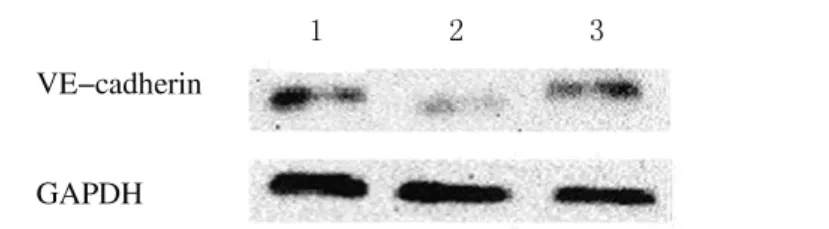
图1 各组A549细胞中VE-cadherin蛋白的表达电泳图Fig.1 Electrophoregram of expressions of VE-cadherin of A549cells in various groups Lane 1:Blank control group;Lane 2:Interference group;Lane 3:Negative control group.
2.4 A549细胞上清液中VE-cadherin的表达水平转染72h后,干扰组上清液中VE-cadherin的表达水平 [(2.89±0.07)μg·L-1]明显低于空白对照组 [(11.43±0.09)μg·L-1]和阴性对照组[(11.15±0.04)μg·L-1](P<0.05),而空白对照组和阴性对照组比较差异无统计学意义(P>0.05)。
2.5 VE-cadherin作用后HUVECs增殖情况 干扰组与阴性对照组上清液比较,A490值在24、48和72h明显降低,差异有统计学意义(P<0.05),见表3。干扰组24、48和72h细胞增殖抑制率分别为29.2%、33.8%和30.1%。
表1 MTS法检测VE-cadherin作用后A549细胞的A490值Tab.1 A490values of A549cells after treated with VE-cadherin detected by MTS method ()
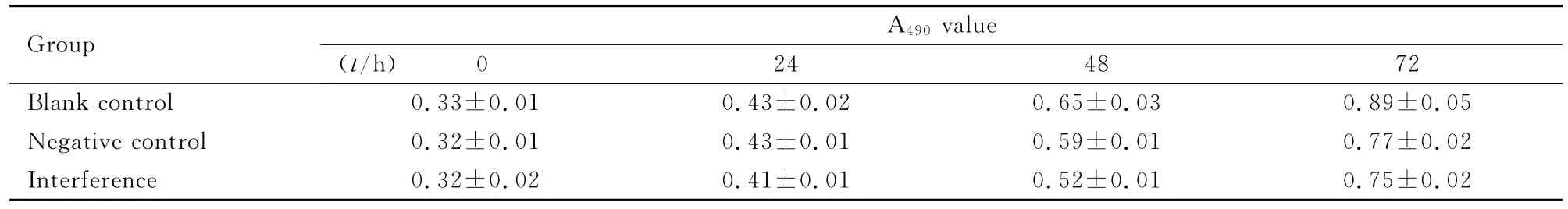
表1 MTS法检测VE-cadherin作用后A549细胞的A490值Tab.1 A490values of A549cells after treated with VE-cadherin detected by MTS method ()
0.89±0.05 Negative control 0.32±0.01 0.43±0.01 0.59±0.01 0.77±0.02 Interference 0.32±0.02 0.41±0.01 0.52±0.010 24 48 72 Blank control 0.33±0.01 0.43±0.02 0.65±0.03 Group A490value(t/h)0.75±0.02
表2 VE-cadherin作用后各组A549细胞周期的变化Tab.2 Changes of cell cycles of A549cells after treated with VE-cadherin in various groups [η/()%]

表2 VE-cadherin作用后各组A549细胞周期的变化Tab.2 Changes of cell cycles of A549cells after treated with VE-cadherin in various groups [η/()%]
09 Negative control 58.33±6.19 9.64±3.52 32.03±3.18 Interference 60.72±4.17 7.89±1.62 31.39±3.M S Blank control 59.30±3.97 8.11±2.11 32.50±2.Group Percentage of A549cells G0/G1 G2/45
表3 VE-cadherin作用后HUVECs增殖的改变Tab.3 Changes of proliferation of HUVECs after treated with VE-cadherin ()

表3 VE-cadherin作用后HUVECs增殖的改变Tab.3 Changes of proliferation of HUVECs after treated with VE-cadherin ()
* P<0.05compared with negative control group.
Group A490 0.91±0.92 Interference 0.27±0.02 0.37±0.04* 0.54±0.04* 0.64±0.030 24 48 72 Negative control 0.26±0.02 0.52±0.02 0.82±0.01 value(t/h)*
2.6 VE-cadherin作用后HUVECs周期和凋亡率 VE-cadherin作用48h后,干扰组与阴性对照组各细胞周期(G0/G1、G2/M 和S)HUVECs所占百分比比较差异无统计学意义(P>0.05)。干扰组和阴性对照组上清液中HUVECs凋亡率分别为(27.12±5.22)%和(13.43±3.50)%,组间比较差异有统计学意义(P<0.05)。见表4和图2。
2.7 VE-cadherin作用后HUVECs小管形成数将干扰组和阴性对照组上清液分别重悬HUVECs,6h后观察小管形成情况,干扰组细胞聚集减少,完整管腔数目明显减少,干扰组和阴性对照组小管形成数量分别为1.53±0.31和14.53±1.51,组间比较差异有统计学意义(P<0.05)。见图3。
表4 VE-cadherin作用后HUVECs周期的变化Tab.4 Changes of cell cycles of HUVECs after treated with VE-cadherin [η/()%]

表4 VE-cadherin作用后HUVECs周期的变化Tab.4 Changes of cell cycles of HUVECs after treated with VE-cadherin [η/()%]
Group Percentag.04 Interference 42.54±2.52 9.49±3.10 47.98±2.M S Negative control 41.94±2.48 11.53±4.66 46.53±4 e of HUVECs G0/G1 G2/42
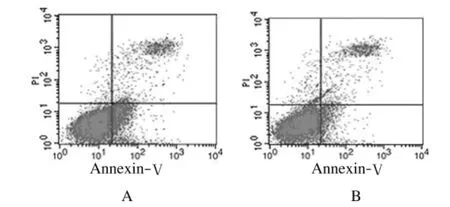
图2 VE-cadherin作用后HUVECs凋亡的改变Fig.2 Changes of the apoptosis of HUVECs after treated with VE-cadherin
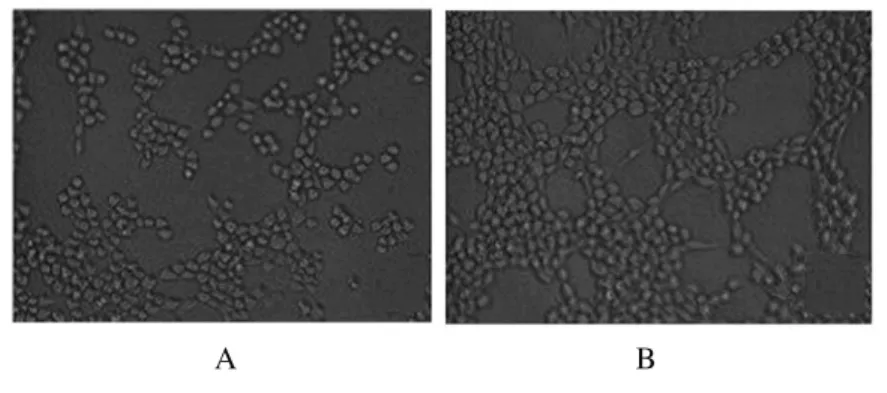
图3 VE-cadherin作用后HUVECs的小管形成Fig.3 Tubule formation of HUVECs after treated with VE-cadherin
3 讨 论
血管生成在肿瘤的生长、侵袭和转移中发挥着至关重要的作用,丰富的血管可以不断地向肿瘤组织提供氧气和营养,并排出肿瘤细胞的代谢产物;同时其远处转移也必须以血管新生为前提和必要条件。目前新的观点认为:血管生成抑制剂还可使肿瘤血管正常化,即使异常的血管在结构和功能上趋于正常。而肿瘤血管和微环境异常是肿瘤化疗和放疗耐受的主要原因,即使用血管生成抑制剂可以提高传统化疗和放疗的疗效。
VE-cadherin是特异性表达于血管内皮细胞的一类细胞黏附分子,定位于内皮细胞与内皮细胞连接处,在维持和控制内皮细胞连接方面起重要作用,且在血管形成过程中必不可少[1]。VE-cadherin氨基端与相邻血管内皮细胞上相同的VE-cadherin同嗜黏附,其羧基末端与2个胞内蛋白β-catenin和γ-catenin相连,2个胞内蛋白属于Armadillo家族,其通过α-catenin将 VE-cadherin与肌动蛋白细胞骨架相连,保证了血管的完整性。新近的研究[4-9]显示:VE-cadherin与肾母细胞瘤、骨肉瘤、恶性黑色素瘤、结肠癌和卡波氏肉瘤等多种恶性肿瘤血管生成关系密切,是乳腺癌转移新的生物标志物。但目前国内外,VE-cadherin与肺癌的关系鲜见报道,仅Dome等[10]研究显示:与健康者比较,NSCLC患者外周血中 VE-cadherin mRNA表达水平并未显著增加。Liao等[11]应用VE-cadherin的抗体阻断VE-cadherin介导的同嗜黏附作用,可以抑制裸鼠皮下种植的Lewis肺癌细胞和人A431上皮癌细胞的生长和转移。然而,肺癌是最常见的恶性肿瘤,全球发病率和死亡率居癌症首位,临床多数肺癌发现时已属中、晚期,其5年生存率不到15%,而其中75%~80%为NSCLC。现有资料未能全面阐述NSCLC血管生成的机制,故有必要寻找新的与NSCLC血管生成相关的基因以进一步完善其发病机制。
本研究前期研究显示:VE-cadherin在NSCLC组织中的表达明显高于正常组织[2],且在癌细胞中大量表达,这促使本文作者猜想VE-cadherin可能参与了NSCLC的发生发展,但是直接对癌细胞起作用还是对血管内皮细胞起作用尚不清楚。本研究利用RNA干扰技术,有效抑制了VE-cadherin在A549细胞中的表达,尚未发现其对A549细胞增殖、周期和凋亡有影响,干扰组细胞上清液中VE-cadherin表达水平明显低于空白对照组和阴性对照组,而空白对照组和阴性对照组比较差异无统计学意义,故后续实验仅研究干扰组和阴性对照组。本研究结果显示:干扰组和阴性对照组上清液对HUVECs增殖、凋亡率和周期变化无影响,表明VE-cadherin对HUVECs有促增殖和抗凋亡作用;本研究结果显示:干扰组HUVECs聚集及形成完整管腔数明显减少,表明VE-cadherin在完整血管管腔的形成过程中起重要作用。以上结果表明:癌细胞为了生长大量合成并分泌VE-cadherin,VE-cadherin促进血管生成,反过来又促进肿瘤生长,这为NSCLC的抗血管生成治疗提供了新的靶点。
VEGF是目前已知的活性最强、特异性最高的内皮细胞促有丝分裂细胞因子,其他的血管生成因子也通过VEGF而起作用。VEGF可促使Rac1依赖性活性氧(ROS)的生成,导致 VE-cadherin Y658和Y731磷酸化,进一步导致细胞间连接的分离和单分子层渗透性增加[12]。Gavard等[13]认为:VEGF通过Src依赖的VaV2磷酸化激活小GTP酶Rac,Rac又促进由PAK介导的VE-cadherin S665磷酸化,引起β-arrestin募集到VE-cadherin上,导致 VE-cadherin内化。血管内皮-蛋白酪氨酸磷酸酶(VE-PTP)加强了VE-cadherin介导的细胞间黏附,VE-cadherin/VE-PTP复合体的分离是白细胞渗出和VEGF诱导的血管渗透性增加所必须的[14]。VEGF诱导VE-cadherin正向连接在鼠肺或睾提肌血管内皮细胞重新排列,这依赖于TSAD,其在内皮细胞连接处与 VE-cadherin、VEGFR2和c-Src形成复合体[15]。总之,VE-cadherin酪氨酸和丝氨酸磷酸化直接影响了VE-cadherin的稳定,从而影响其内皮渗透性,但其具体机制还有待进一步研究。
综上所述,VE-cadherin通过促进血管生成来参与NSCLC的发生发展,有望成为NSCLC抗血管生成治疗的新靶点。因此,本文作者设想通过抑制VE-cadherin的异常表达,进一步抑制NSCLC的生长、浸润和转移,从而改善患者的预后,为NSCLC的靶向治疗提供一个新的靶点。
[1]Vestweber D.VE-cadherin:the major endothelial adhesion molecule controlling cellular junctions and blood vessel formation [J].Arterioscler Thromb Vasc Biol,2008,28(2):223-232.
[2]陶 娟.非小细胞肺癌中VE-cadherin和E-cadherin的表达及意义 [D].重庆:重庆医科大学,2010.
[3]Ha CH,Bennett AM,Jin ZG.A novel role of vascular endothelial cadherin in modulating c-Src activation anddownstream signaling of vascular endothelial growth factor [J].J Biol Chem,2008,283(11):7261-7270.
[4]Kirschner KM,Sciesielski LK,Scholz H.Wilms’tumour protein Wt1stimulates transcription of the gene encoding vascular endothelial cadherin [J].Pflugers Arch,2010,460(6):1051-1061.
[5]Zhang LZ,Mei J,Qian ZK,et al.The role of VE-cadherin in osteosarcoma cells[J].Pathol Oncol Res,2010,16(1):111-117.
[6]Khanna P,Yunkunis T,Muddana HS,et al.p38MAP kinase is necessary for melanoma-mediated regulation of VE-cadherin disassembly [J].Am J Physiol Cell Physiol,2010,298(5):1140-1150.
[7]Li H,Shi X,Liu J,et al.The soluble fragment of VE-cadherin inhibits angiognesis by reducing endothelial cell proliferation and tube capillary formation [J].Cancer Gene Ther,2010,17(10):700-707.
[8]Guilluy C,Zhang Z,Bhende PM,et al.Latent KSHV infection increases the vascular permeability of human endothelial cells [J].Blood,2011,118(19):5344-5354.
[9]Fry SA,Sinclair J,Timms JF,et al. A targeted glycoproteomic approach identifies cadherin-5as a novel biomarker of metastatic breast cancer [J].Cancer Lett,2013,328(2):335-344.
[10]Dome B,Timar J,Dobos J,et al.Identification and clinical significance of circulating endothelial progenitor cells in human non-small cell lung cancer [J]. Cancer Res,2006,66(14):7341-7347.
[11]Liao F,Li Y,O’Connor W,et al.Monoclonal antibody to vascular endothelial cadherin is a potent inhibitor of angiogenesis,tumor growth,and metastasis [J].Cancer Res,2000,60(24):6805-6810.
[12]Monaghan-Benson E,Burridge K.The regulation of vascular endothelial growth factor-induced microvascular permeability requires Rac and reactive oxygen species [J].J Biol Chem,2009,284(38):25602-25611.
[13]Gavard J,Gutkind JS. VEGF controls endothelial-cell permeability by promoting the beta-arrestin-dependent endocytosis of VE-cadherin [J]. Nat Cell Biol,2006,8(11):1223-1234.
[14]Broermann A,Winderlich M,Block H,et al.Dissociation of VE-PTP from VE-cadherin is required for leukocyte extravasation and for VEGF-induced vascular permeability in vivo [J].J Exp Med,2011,208(12):2393-2401.
[15]Sun Z,Li X,Massena S,et al.VEGFR2induces c-Src signaling and vascular permeability in vivo via the adaptor protein TSAd [J].J Exp Med,2012,209(7):1363-1377.
猜你喜欢
中日友好医院学报(2022年4期)2022-10-15
湖南畜牧兽医(2021年6期)2022-01-24
食品安全导刊(2021年21期)2021-08-30
看世界·学术下半月(2020年7期)2020-09-10
中国眼镜科技杂志(2019年9期)2019-11-11
天津医科大学学报(2019年6期)2019-08-13
天然产物研究与开发(2018年11期)2018-11-30
广东农业科学(2017年10期)2018-01-25
中国医学创新(2017年22期)2017-11-15
中国病理生理杂志(2015年8期)2015-12-21
